| XXVII. El Período Pensilvánico, la época de mayor producción de carbón | Página de título | XXIX. El carbón y su presencia en la naturaleza |
[ p. 373 ]
Las plantas de algún tipo se encuentran en todas las tierras y en todos los climas. No se limitan a los suelos húmedos de los valles, sino que también viven en las montañas más altas e incluso con poca frecuencia en los desiertos más áridos. En las aguas dulces poco profundas se encuentran numerosos tipos de plantas, y en los fondos oceánicos, hasta una profundidad de unos 180 metros, crecen las algas marinas adheridas. Fijados a rocas, árboles y otros objetos se encuentran los líquenes marrones y grises, una fina película de materia vegetal que se extiende. En los bosques húmedos y en las praderas crecen los fimgi, incluyendo los hongos venenosos comunes, plantas carentes de clorofila que se alimentan del material húmico (moho vegetal) de la vegetación muerta y moribunda. Una gran variedad de plantas microscópicas, las bacterias, viven en todas partes: en el suelo, en casi todas las aguas, en los océanos, sobre y dentro de otras plantas y animales. Estas y los hongos son los principales agentes de la naturaleza para disolver organismos muertos, consumir oxígeno y liberar dióxido de carbono. Las bacterias de la levadura convierten el azúcar en alcohol mediante la fermentación y se utilizan en la elaboración de licores alcohólicos; otras bacterias son útiles en la elaboración de queso; otras causan enfermedades y se encuentran incluso en todos los organismos sanos.
Floras Paleozoicas. — Del registro geológico aprendemos que la primera flora conocida surgió en la segunda mitad del Devónico, aunque se conocen algunas plantas terrestres del Silúrico. Sin embargo, probablemente tengamos razón al considerar que las tierras bajas estaban cubiertas de un verdor al menos desde el comienzo del Champlainiano, pero que las plantas primitivas aún no habían adquirido suficiente tejido leñoso en su organización como para preservarse en los sedimentos, por lo que carecen de importancia en la geología histórica. Desde el Devónico Medio, las plantas terrestres con tejido leñoso se han vuelto cada vez más prolíficas y han alcanzado mayores dimensiones y estatura, lo que significa que también han alcanzado gradualmente un crecimiento individual más largo.
Vida pantanosa del Pensilvánico. — Las floras carboníferas del Paleozoico tardío son bien conocidas, y se estima que se han descrito más de tres mil especies (véanse las figs., págs. 360 y 374). Destacan por su distribución casi mundial [ p. 374 ] [ p. 375 ] y su exuberancia y abundancia, junto con un sotobosque denso y variado. Sus representantes más llamativos en número y tamaño son los árboles de escama, una especie de árbol perenne con hojas aciculares comparativamente pequeñas; algunos de estos árboles alcanzaban más de 30 metros de altura y un diámetro de hasta 1,8 metros (véase la lám., pág. 377). En ellos no hay anillos de crecimiento, lo que indica también la ausencia de estaciones hasta casi el final del Pensilvánico. Otro grupo notable, los gigantescos calamites o juncos (lám., pág. 381), alcanzaban al menos 18 metros de largo y 38 centímetros de grosor. Se asemejaban a los cañaverales y matorrales de bambú. Esta flora también incluía muchas formas parecidas a helechos, delicadas y resistentes, algunas trepadoras, mientras que otras se convertían en árboles majestuosos (Fig., pág. 374); la mayoría producía semillas, pero algunas producían esporas y, por lo tanto, eran helechos verdaderos.

En general, estos bosques debieron alcanzar copas superiores a los 12 metros. Crecieron rápidamente y sus maderas eran blandas e incluso esponjosas, como se aprecia en la corteza lisa, dura y persistente, en las células grandes y de paredes delgadas, con una cantidad comparativamente grande de tejido medular y cortical y amplios espacios intercelulares, y en los numerosos poros de agua en las hojas.
Los tonos verdes eran el color dominante, y la monotonía del verdor no se veía realzada por flores brillantes. Sin embargo, había flores, pero de escaso tamaño, insignificantes y sin duda poco atractivas. Probablemente más de la mitad de la flora era heterosporosa (esporogénica) y podemos considerar con seguridad que la mayoría de las plantas más comunes de las Medidas del Carbón no tenían semillas. La fertilización aún no se lograba mediante insectos melíferos y poleníferos, como es tan común entre las plantas con flores actuales, sino que en la mayoría de las floras del carbón se producía por las lluvias y los vientos. En el momento en que los árboles y helechos productores de esporas las liberaban, todo el bosque estaba cubierto de un polvo amarillo verdoso o marrón, y algunos carbones están compuestos principalmente por estas esporas (White y Jeffrey).
Las esporas (lám., pág. 377, figs. 4-6) se diferencian de las semillas en que estas últimas dan lugar directamente a plantas sexuadas, mientras que las primeras también se desarrollan en plantas sexuadas, pero estas tienen una vida corta y, a su vez, dan lugar a las plantas asexuadas y longevas que contienen las esporas. En otras palabras, las plantas con esporas tienen una alternancia de generaciones, mientras que las plantas con semillas tienen un desarrollo directo. Este tema se abordará más adelante en este capítulo.
El aire no estaba perfumado con olores dulces, pues no había miel, pero es probable que hubiera olores resinosos como los que impregnan los bosques de coníferas vivos.

Fig. 1, Alethopteris grandifolia, 2. Neuropteris fasciculata; 3, N. capitata; 4, Pseudopecopteris mazonana, with spore cases; 5, probably a seed-bearing fern-like tree, restored by Kidston; 6, Sphenopteris mixta; 7, Archaeopteris stricta; 8, Aneimites x 2; 9, same with a young seed. x 4; 10, same, one of the seeds x 3. Fig. 1—4, from the Illinois and Ohio State Surveys; Figs. 8—10 after David mite.
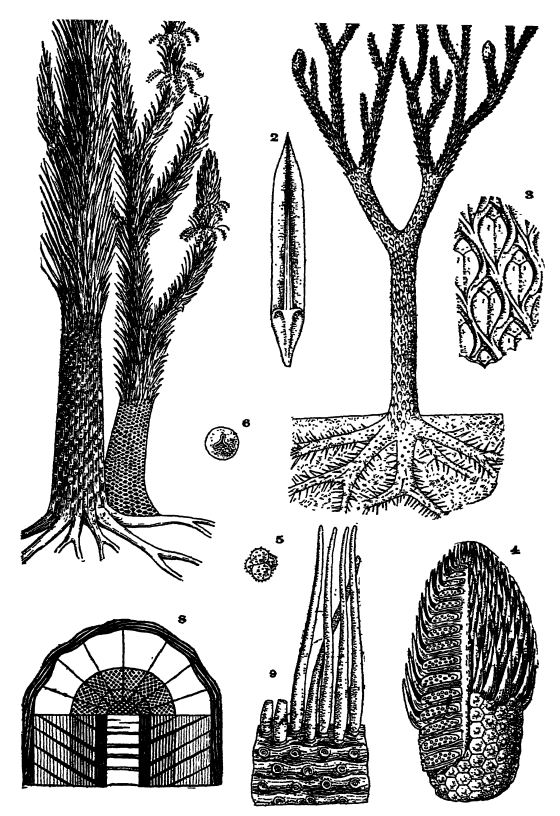
Fig. 1, restoration of the tree Lepidodendron, about x 1/100; 2, leaf of the sporebearing cone, about x ⅓, those of the branches being very similar but longer; 3, outer surface of tree, showing nature of leaf bases, about x ½ (L. cbovatum ) ; 4, sporebearing cone (Lepidostrobus) , with and without leaves, sectioned to show spores and spore-developing cases (sporangia) ; 5, bunch of four microspores; 6, macrospore; 7, two species of the tree Sigtllaria, restored, about x A; 8, section through trunk, showing relative thicknesses of outer leaf-bearing layer, two zones of cork-like bark, comparatively thin internal cylinder of wood, and medial pith; 9, roots of these trees (Stigmaria ficoides ) , with rootlets and scars of their attachment, figs. 1-6 after Steinmann; Figs. 7 and S after Zittel. (677)
[ p. 378 ]
En estos bosques de la época de Pensilvania se podían ver volando los insectos más grandes que jamás hayan existido: grandes libélulas, que alcanzaban una envergadura de 73 centímetros. Las cucarachas enormes abundaban por doquier en gran variedad, y no eran raras las de 10 centímetros (véase lámina, pág. 363, fig. 4). Por lo general, estos insectos eran carnívoros y no transferían el polen de una flor a otra, con la consiguiente fecundación cruzada, como ocurre tan comúnmente entre los insectos vivos en las plantas actuales. Los insectos más pequeños eran depredados por escorpiones y arañas; estas últimas no tejían telarañas, sino que vivían en el suelo o en troncos podridos, junto con numerosos miriápodos o arañas de mil patas. Ningún insecto de esta época, que se sepa, producía chirridos ni otros sonidos, y el susurro del viento entre los árboles solo se veía interrumpido por el croar de los anfibios en las marismas. Los anfibios eran comunes en los pantanos, y es probable que muchos reptiles pequeños corrieran por el suelo y entre los árboles. No se observaron animales terrestres de mayor tamaño, como los que conocemos, ni aves.
Clasificación. — Para nuestros propósitos, las plantas se dividen más convenientemente en (1) aquellas que no tienen flores pero se reproducen por esporas y pasan por una alternancia de generaciones (Criptógamas), como se explicó anteriormente; y (2) aquellas que tienen flores y se reproducen directamente a partir de semillas (Fanérogamas).
En este capítulo estudiaremos las plantas terrestres más antiguas y, como éstas pueden conservarse como fósiles solo si tienen material fibroso o leñoso, el paleontólogo no se ocupa directamente de las criptógamas inferiores.
Todas las plantas superiores, superiores a los musgos en cuanto a organización, se caracterizan por tener raíces y por la presencia en sus tejidos blandos de lo que se conoce como haces vasculares. Estos haces están compuestos de fibras resistentes, huecas y entrelazadas, cuya función principal es servir como canales conductores (de ahí el nombre vascular, de vascidum, pequeño vaso) a través de los cuales pasan los diversos nutrientes, el agua y las sales ascienden y el azúcar y las proteínas descienden. En segundo lugar, son importantes para formar los esqueletos de soporte de las plantas. Todo el material leñoso está compuesto por estas hebras, y es esta sustancia resistente la que permite que las hojas y otras partes de las plantas impriman en los sedimentos las huellas de su existencia anterior.
La clasificación adicional puede expresarse de la siguiente manera:
Criptógamas inferiores, plantas portadoras de esporas
- Talofitas, bacterias, hongos, la mayoría de plantas marinas (algas)
- Briófitas, musgos
Criptógamas superiores, también portadoras de esporas
- Pteridofitas, helechos (Lám., pág. 376, Figs. 1-5)
- Artrófitos, juncos vivos y calamitas paleozoicas (Lám., pág. 381, Figs. 1, 2)
- Lepidofitas, árboles de escamas o licopodos (Lám., pág. 377)
[ p. 379 ]
Fanerógamas, plantas que producen semillas
- Gymnospenns, plantas con flores imperfectas y poco visibles.
- Pteridospermofitas (o Pteridospermas, plantas parecidas a los helechos (Lám., pág. 376, Figs. 6—10)
- Cicadofitas, cícadas (Figs., págs. 27 y 3S6)
- Coniferofitas
- Cordaites, antiguos árboles perennes (Lám., p. 381, Figs. 3-6)
- Coníferas, árboles perennes modernos
- Gingkos, árboles de culantrillo (Lám., pág. 381, Fig. 7)
- Angiospermas, verdaderas plantas con flores
¶ Plantas sin flores que producen esporas
¶ Helechos o Pteridofitas
Casi todos conocemos el aspecto de un helecho vivo, pero al aplicar este conocimiento a las plantas paleozoicas de apariencia similar, ni siquiera los mejores botánicos pueden estar seguros en la actualidad, en muchos casos, de que la forma estudiada sea un helecho verdadero. En el siglo pasado, aún se creía que la mitad de las plantas carboníferas eran helechos verdaderos y que estas hermosas formas dominaban la flora de aquella época. Sin embargo, gracias a la acumulación de conocimiento botánico, y más concretamente al descubrimiento de especímenes con órganos reproductivos, ahora se sabe que muchos de los llamados helechos paleozoicos tenían semillas y, por lo tanto, pertenecían a una organización superior, por lo que deben clasificarse de forma totalmente independiente de los helechos verdaderos. Estos eran los helechos con semillas, en muchos sentidos una transición entre los helechos con esporas y las plantas con semillas (véase lámina, pág. 376, figs. 6-10). Probablemente más de la mitad de las plantas paleozoicas que antiguamente se consideraban helechos (unas mil especies) acabarán demostrando ser productoras de semillas.
Helechos Vivos. — Hoy en día existen más de seis mil tipos diferentes de helechos, desde formas diminutas y delicadas hasta los hermosos helechos arborescentes de las tierras tropicales, que alcanzan una altura de 15 metros. La mayor variedad se encuentra en los trópicos, pero los helechos herbáceos se encuentran en todos los climas, incluso en la gélida Groenlandia. Sin embargo, por regla general, prefieren las regiones boscosas y húmedas. El helecho es la generación asexuada. Sus esporas suelen desarrollarse en el envés de las hojas, en cápsulas, algunas de las cuales siempre están agrupadas. En cada cápsula hay muchas esporas microscópicas, todas iguales, y estas a menudo se desarrollan en cantidades incontables (véase la Lám., pág. 376, Fig. 4).
Algunas de estas esporas pueden germinar en el suelo, dando lugar a una planta sexuada, el protalo, que suele ser un brote verde, primitivo y acorazonado que yace postrado en el suelo y a menudo mide [ p. 380 ] menos de 1,27 cm de largo. En su envés se desarrollan los órganos masculinos que dan lugar a células masculinas o polen, y los órganos femeninos que, al polinizarse, desarrollan ciertas células en plantas asexuadas. Tenemos, por lo tanto, una alternancia de generaciones: plantas asexuadas dan lugar a plantas sexuadas, y estas, a su vez, producen los helechos asexuados.
En los helechos herbáceos más pequeños, existe un tallo corto, robusto y sotobosque que da origen a numerosos tallos foliares, cada uno de los cuales se desarrolla enrollado en espiral y permanece así durante un año o más. Posteriormente, los tallos se desenrollan y crecen rápidamente hasta convertirse en hojas completamente desarrolladas, generalmente llamadas frondas. Estas pueden alcanzar una gran longitud, como en los helechos arborescentes, y se llaman frondas porque las hojas están compuestas de numerosos folíolos.
En los helechos arborescentes existe un solo tallo sin ramificar, similar al tronco de una palmera, y las frondas, compuestas por una o más hojas con sus numerosos folíolos, se desarrollan en la parte superior como una corona, desenrollándose cada fronda al alcanzar su máximo crecimiento. El tronco presenta las cicatrices de las frondas que se han desprendido, y bajo esta capa más externa suele haber una zona gruesa, la corteza, que rodea el cilindro leñoso central, más o menos grande (Lám., p. 376, Fig. 5).
Helechos fósiles. — Los helechos ancestrales (Marattiales, o helechos que se desarrollan a partir de una sola célula) eran raros en el Devónico y no son comunes como fósiles hasta el Pensilvánico, donde muchas especies de las clases herbáceas más pequeñas aparecen asociadas con los helechos arborescentes (véase Lám., pág. 376, Figs. 1, 4). Estos últimos tipos se hicieron visibles por primera vez en el Pensilvánico y fueron abundantes en la segunda mitad de este período y a principios del Pérmico. En algunos casos, el tallo alcanzó una altura de 15 metros, y en ciertas formas las frondas se desarrollaron en dos o cuatro columnas paralelas, pero en la mayoría de los casos se dispusieron en espiral alrededor del tronco (véase Lám., pág. 376, Fig. 5).
¶ Arbustos o Artrófitos
Entre las plantas vivas existe un pequeño grupo de formas con una distribución muy amplia, conocidas popularmente como juncos (similar a la Lám., pág. 381, Figs. 1, 2). Por lo general, son pequeñas, de menos de 45 cm de altura, y aparecen en lugares húmedos a principios de año; sin embargo, en América Central, del Sur y en Cuba, se encuentra una forma gigante en arboledas, que alcanza una altura de 12 metros, pero con un tallo que no supera los 2,5 cm de diámetro. Esta y todas las demás formas vivas, unas veinticinco, pertenecen al género Equisetum (véase también la Lám., pág. 468).
Este género actual de juncos posee en el suelo un peciolo horizontal, perenne, sin sexado, rastrero y ramificado que, a intervalos de [ p. 382 ], da lugar a tallos aéreos erguidos y raíces descendentes. Los tallos constan de una delgada zona leñosa, externamente estriada, con una gran médula central. A intervalos regulares, el tallo se divide en internodios mediante tabiques transversales llamados nudos, y de estos surgen los verticilos de hojas pequeñas o reducidas, con una o más nervaduras longitudinales, y también las ramas, cuando las hay. Las ramas repiten, a menor escala, las características generales de los tallos. En la parte superior del tallo o sobre las ramas se encuentran los conos portadores de esporas.

Fig. I, árboles de calamita restaurados, el de la izquierda con una caja de esporas, aproximadamente x 1/100; 2, extremo basal de una calamita; 3, árbol de cordaita restaurado (Dorvcordaita), aproximadamente x 1/100, también mostrando la naturaleza de las flores similares a amento; 4, diferentes tipos de cordaitas aproximadamente x 1/40; 5, Trigonocarpon ornatum, un fruto o semilla de una cordaita; 6. Rhabdocarpus apiculatus, semilla de una oordaita; 7, ramita de flores masculinas de árbol de culantrillo (Ginaba biloba) vivo, y a la derecha la semilla madura.
Calamitas fósiles. — En el Paleozoico, desde el Devónico hasta el Pérmico, pero más especialmente en el Pensilvánico, existió una gran variedad de juncos antiguos, los más grandes de los cuales se conocen como calamitas (Lám., pág. 381, Figs. 1, 2). Eran plantas prolíficas y White afirma que en América alcanzaron un diámetro de hasta 30 cm y una longitud de más de 9 metros. En ellas, el cilindro de madera era mucho más grueso que en los juncos actuales, y se conocen ejemplares con 5 cm de corteza por fuera del mismo grosor de madera. Las capas de madera secundaria se añadían externamente bajo la corteza, un tipo de crecimiento presente en los árboles modernos. Los nudos no eran equidistantes en toda su longitud como en las formas actuales, sino que en los primeros crecimientos estaban progresivamente más separados y los tallos se engrosaban rápidamente. Las calamitas también presentaban una ramificación mucho más abundante que en los juncos actuales, y sus hojas eran proporcionalmente más grandes. La parte superior del tronco consistía en un penacho denso de círculos de hojas muy juntos, y los conos con esporas reproductivas se encontraban con mayor o menor abundancia en las ramitas frondosas más pequeñas o en la parte superior del tallo principal. Por regla general, las esporas eran todas iguales en los artrófitos del Paleozoico, pero en algunos casos se diferenciaban en tipos pequeños y grandes (heterosporas, véase lámina, pág. 377, figs. 5 y 6).
¶ Árboles de escala o lepidofitas
Licópodos Vivos. — Licópodos es el nombre de un grupo de plantas perennes herbáceas, primitivas y ampliamente distribuidas, conocidas como pinos terrestres, licopodios y pinos corredores. Estos últimos se utilizan ampliamente en decoraciones florales. Existen alrededor de cien especies vivas del género Lycopodium; el nombre, que proviene del griego «pata de lobo», se debe a la apariencia de sus raíces. En la flora actual, los licopodios nunca crecen en masas densas que excluyan a otras plantas, por lo que no son visibles. Su apariencia general sugiere la de los musgos más grandes o las ramas más pequeñas de los pinos, ya que las hojas son siempre notablemente pequeñas.
Lepidofitas fósiles. — En el Paleozoico, desde el Devónico hasta el Pérmico Medio, pero especialmente durante el Pensilvánico, [ p. 383 ] las lepidofitas fueron las plantas dominantes y alcanzaron proporciones gigantescas en comparación con los licopodios actuales, considerablemente reducidos, pues las primeras se encontraban entonces en la cúspide de su evolución (Figs., págs. 360 y 374). En el Pensilvánico, estos árboles liberaron sus gérmenes reproductivos heterosporosos en cantidades tan enormes que constituyeron un factor importante en la producción de carbón (véase Lám., pág. 377, Figs. 4-6).
A finales del Paleozoico existían dos tipos principales de lepidofitas: lepidodendron o árboles de escamas y sigillaria o árboles de focas, términos que hacen referencia a la apariencia escamosa de las bases de las hojas en los troncos y ramas de estos árboles. Se han descrito más de cien especies de lepidodendron, que abarcan desde el Devónico hasta el Pérmico (Lám., pág. 377, figs. 1-3). El tronco se estrechaba lentamente y, en algunas formas, alcanzaba una altura de más de 30 metros y un diámetro de 0,9 metros. Estos árboles, en su aspecto general, se diferenciaban de los modernos en que el tronco y muchas de sus ramas se bifurcaban con bastante regularidad. Las ramas más delgadas terminaban, por lo general, en conos lineales u ovalados con esporas de entre 2,5 y 30 cm de largo (Lám., pág. 377, fig. 4).
Las hojas del Lepidodendron eran aciculares, siempre comparativamente pequeñas, pero con un ancho en la base de 1,27 cm y una longitud de 15 a 18 cm (Lám., pág. 377, Figs. 1, 2). En sus caras inferiores había dos surcos laterales, y en estos huecos se situaban los diminutos pero numerosos órganos respiratorios para la extracción del dióxido de carbono de la atmósfera. Las hojas caían pronto y generalmente se encuentran adheridas solo a las ramitas jóvenes en crecimiento. Las bases de las hojas tenían forma de diamante y estaban dispuestas en espiral sobre las ramas y troncos, formando así las marcas características de los árboles. La hoja estaba adherida en la parte superior del rombo (Lám., pág. 377, Fig. 3).
De Sigillaria (Lám., pág. 377, Fig. 7), se han descrito alrededor de cien especies, que se diferencian del Lepidodendron en que rara vez ramifican. Se han encontrado troncos con un diámetro, justo por encima de las raíces, de 1,8 metros, y uno alcanzó una altura de casi 30 metros sin ramificar. En la punta, a lo largo de unos 3 metros, estos árboles estaban cubiertos de hojas erectas, rígidas y herbáceas, que en la mayoría de los casos eran similares a las del Lepidodendron, pero a veces eran mucho más grandes y anchas.
Las cicatrices foliares de Sigillaria se disponían en filas verticales, alternando las cicatrices de series adyacentes. En muchas formas, la superficie de los troncos presentaba nervaduras longitudinales, cada una de las cuales presentaba una sola fila de cicatrices foliares (véase la lámina p. 377, fig. 7).
[ p. 384 ]
Las Sigillaria son desconocidas en el Devónico y se extinguieron antes del cierre del Pensilvánico.
La estructura del tronco y las ramas en las lepidofitas fósiles era peculiar en el sentido de que el mayor espesor debajo de la capa exterior con las cicatrices de las hojas estaba compuesto por dos tipos de corteza parecida al corcho que rodeaba un cilindro leñoso interno comparativamente delgado que puede haber estado completamente lleno de madera que crecía hacia adentro —el tipo de crecimiento endógeno— o con más o menos médula (Lám., p. 377, Fig. 8). Se agregó madera secundaria de hasta 2 pulgadas de espesor al corcho exterior debajo de la capa de hojas, y como este corcho también era resistente, los árboles tenían un poder de resistencia considerable contra los vientos y las tormentas de esa época, pero eran menos rígidos que los árboles forestales modernos.
Básicamente, tanto Lepidodendron como Sigillaria terminaban en entre cuatro y siete raíces que se cree que se extendían casi horizontalmente en el suelo, y no existía una raíz vertical o pivotante que continuara el tronco como en los árboles modernos. Estas raíces se dividían una o dos veces y se estrechaban hasta una punta, excavando en todas direcciones a través de la masa de vegetación en descomposición del pantano. En su superficie había cicatrices circulares dispuestas en grupos de cinco, a las que se adherían apéndices redondos y tapered que irradiaban en todas direcciones; estos últimos alcanzaban una longitud de 38 cm y servían para extraer alimento y agua del suelo (Lám., p. 377, Fig. 9). Estas raíces se conocen como Stigmaria (del latín stigma, marca) y se ven con mucha frecuencia en las arcillas subyacentes a los yacimientos de carbón.
Los conos de las lepidófitas, si bien se parecían mucho a los de las plantas perennes o coníferas modernas, no tenían semillas, sino que estaban llenos de esporas (Lám., pág. 377, Figs. 1, 4-6). Estas últimas crecían en grandes cápsulas, que contenían esporas diminutas, las microesporas o partes masculinas fecundantes (Lám., pág. 377, Fig. 5), o grandes, las macroesporas femeninas (Lám., pág. 377, Fig. 6), a menudo cuarenta veces más grandes que las microesporas. Estos dos tipos de esporas se desarrollaban en conos diferentes o en partes distintas del mismo cono, y había muchas más esporas diminutas que de grandes.
¶ Plantas con flores que producen semillas o fanerógamas
Las plantas con semillas, sumamente variadas y numerosas, se dividen en dos grandes divisiones. El grupo, geológicamente más antiguo y primitivo, se conoce como gimnospermas (o plantas con semillas desnudas), en el que las partes masculinas y femeninas pueden estar en flores separadas o combinadas en una sola, pero siempre son discretas y a menudo carecen [ p. 385 ] de color brillante. Las semillas se desarrollan en ovarios abiertos, que se dice que están desnudos porque el polen cae directamente sobre los óvulos, mientras que en las plantas superiores o con verdaderas flores (angiospermas) el ovario está cubierto por un estigma que recibe el polen.
¶ Helechos con semillas o pteridospermas
Al hablar de los helechos en una página anterior, se afirmó que en el Paleozoico existían muchas plantas con la apariencia de verdaderos helechos, pero que se sabía que producían semillas en lugar de esporas. En otras palabras, estas plantas no presentaban alternancia de generaciones para completar su ciclo de desarrollo, sino que eran plantas sexuadas, es decir, crecían directamente de la semilla, dando lugar a plantas masculinas que desarrollaban polen o a plantas femeninas que desarrollaban semillas que fructificaban y se desarrollaban en un embrión mientras estaban adheridas a la planta. Tenían la apariencia general de los helechos y sus semillas eran muy similares a las de las cícadas. El descubrimiento de estas pteridospermofitas, o, en el lenguaje común, pteridospermas, es uno de los logros más destacados de los paleobotánicos de principios de este siglo. En el origen de las semillas, las pteridospermas dieron un gran paso adelante, y se convirtieron en la característica dominante de las floras posteriores (véase lámina, pág. 376, figs. 6-10).
Las semillas y el polen se originaban en plantas sexuadas independientes. Las semillas podían ser pequeñas y encontrarse en las puntas de las hojas, o bien colgaban de la cara inferior en los extremos exteriores de las hojas lobuladas, o, si eran grandes, estaban adheridas a la gruesa nervadura central de la fronda; nunca se presentaban en conos. El tamaño de las semillas variaba desde 0,63 cm hasta al menos 5 cm de longitud, a menudo alargadas en el extremo libre, donde se encontraba la entrada a la cámara polínica. El embrión o nuez estaba rodeado por una cáscara que, en algunas formas, tenía un exterior grueso y carnoso y una capa interna dura y pétrea. Semillas similares también se encontraron en las cordaítas, en los gingkos (Lám., pág. 381, Figs. 5-7) y en las cícadas.
Algunas de las helechos con semillas tenían tallos largos y delgados, y trepaban entre otras plantas; muchas eran herbáceas, y otras (Psaronius) eran tan altas y robustas como los helechos arborescentes. Se originaron en helechos primitivos anteriores al Devónico Medio, y su desarrollo culminaba en el Pensilvánico y el Pérmico temprano, pero aún no se conoce ninguna más allá del Paleozoico.
¶ Cícadas o cicadofitas
En la flora moderna de climas cálidos, las cícadas actuales, de unas 110 especies, son los restos de un grupo una vez más diversificado, nunca muy común en la flora paleozoica, pero dominante en la de la primera mitad del Mesozoico. Un ejemplo vivo sorprendente [ p. 386 ] es la «palma de sagú» de Ceilán, la fuente de nuestro sjigo comestible. La importancia de estas antiguas plantas con semillas en relación con las demás plantas con flores la hemos aprendido en gran medida de los estudios de G. R. Wieland; parecen haber surgido de las pteridospermas, posiblemente ya en el Devónico. Las cicadas aparecieron con el Aliddle Pensilvánico, pero no fueron comunes hasta la época de las Triásicas.
La característica más destacada de las cícadas es que sus tallos leñosos, cortos y columnares, con centros medulares muy grandes, están recubiertos por una gruesa armadura de bases foliares persistentes, con una masa intermedia similar al fieltro. Los troncos varían en tamaño, desde muy pequeños hasta los de 18 metros de longitud en las cícadas actuales. Las cícadas crecen lentamente y no maduran hasta muchos años después, y un tronco de 1,8 metros de largo puede tener 1000 años. A veces se conservan de forma excepcional en los estratos mesozoicos. Las hojas son de diversos tipos, pero comúnmente son del tipo de palmera pinnada, y se utilizan mucho en la decoración floral (véanse las figuras, pág. 27 y superiores).

¶ Coníferas o Coniferofitas
Cordaitas, o Coníferas de Hoja Grande. — En el Paleozoico, comenzando al menos con el Devónico Superior, se encuentran troncos cuya estructura es [ p. 387 ] similar a la de los pinos o coníferas modernos. En el Pensilvánico, se observan a menudo moldes de estos troncos en las areniscas, y en las pizarras de los tejados de las carboneras abundan las hojas largas y estriadas de estos árboles; de hecho, a menudo constituyen una buena parte de las carboneras. Estas plantas, conocidas como cordaitas (en honor al paleobotánico Corda), incluían diversas formas y fueron las gitanospermas dominantes del Paleozoico. Eran árboles perennes de madera blanda, altos y esbeltos, a veces de 36 metros de altura y 0,9 metros de grosor. En estos árboles altos, dos tercios del tronco carecían de ramas, y el tercio o cuarto superior del tallo presentaba una copa densa y abundante en hojas simples de gran tamaño (véase Lám., pág. 381, Figs. 3-6).
Las hojas de las cordaítas siempre fueron grandes, y se conocen algunas de 1.8 metros de largo y 15 centímetros de ancho. Algunas tenían forma de correa, con los extremos libres puntiagudos o redondeados; otras eran parecidas a la hierba, de 50 centímetros de largo y 1.25 centímetros de ancho. La sustancia foliar era gruesa y presentaba nervaduras paralelas, como en la yuca actual (véase lámina, pág. 381, figs. 3 y 4).
Si bien el tronco leñoso de las cordaítas era muy similar al de los pinos modernos, se distingue de estos últimos por tener siempre una médula central que puede tener un diámetro de hasta 12,7 cm. La madera nueva se formaba bajo la corteza gruesa; es decir, se añadía externamente a las capas anteriores (crecimiento exógeno).
Las cordaítas estaban emparentadas con las coníferas, pero, como se desprende de la descripción anterior, ambos grupos presentaban una composición muy diferente. Las primeras presentaban una gran ramificación solo en la parte superior, carecían de conos con semillas, sino que las desarrollaban en amentos, las hojas no eran aciculares, sino estriadas, y, por último, los troncos presentaban un centro medular. Las cordaítas estaban estrechamente emparentadas con los gingkos.
Los gingkos se diferencian de las cordaitas en que tienen hojas mucho más pequeñas y lobuladas, mientras que los troncos están ramificados en casi toda su longitud (véase lámina, pág. 381, fig. 7).
Coníferas o Árboles Perennifolios. — Los árboles perennifolios, o coníferas verdaderas de flora actual, cuyas semillas se desarrollan en conos femeninos prominentes y el polen en conos masculinos, no se conoce que sean de origen más antiguo que el Pérmico, pero ahora son las gimnospermas más comunes. Este orden abarca las araucarias, las sequoias o «árboles grandes» de California (árboles estrechamente relacionados, pero de menor tamaño, que se remontan al Pérmico), el pino, la pícea, el abeto, el enebro, el alerce, el ciprés y el tejo.
[ p. 388 ]
¶ Angiospermas o plantas con flores verdaderas
Como las verdaderas plantas con flores no aparecen en la Geología Histórica hasta el Cretácico, su descripción se pospone al capítulo dedicado a esa época.
¶ Lectura colateral
E. W. Berry, Paleobotánica: Un bosquejo del origen y la evolución de las floras. Informe anual del Instituto Smithsoniano, 1918, 1920, págs. 289-407.
C. J. Chamberlaix, Las cícadas vivientes. Chicago (University of Chicago Press), 1919.
J. M. Coulter y C. J. Chamberlain, Morfología de las gjunnospermas. Chicago (University of Chicago Press), 1910.
D. H. Scott, Introducción a la botánica estructural, Parte II, Plantas sin flores. Londres (Black), 1904.
A. C. Seward, Vínculos con el pasado en el mundo vegetal. Cambridge (University Press), 1911.
D. White, Flora fósil de las zonas carboníferas inferiores de Misuri. Servicio Geológico de Estados Unidos, Monografía 37, 1899.
D. White, La sucesión estratigráfica de las floras fósiles de la Formación Pottsville en el yacimiento de carbón antracita del sur, Pensilvania. Servicio Geológico de Estados Unidos, Vigésimo Informe Anual, Parte II, 1900, págs. 749-918.
G. R. Wieland, Araucariales, Cycadales y Cordaitales. Enciclopedia Americana, 1918, págs. 135-138, 351-360, 683-686.
| XXVII. El Período Pensilvánico, la época de mayor producción de carbón | Página de título | XXIX. El carbón y su presencia en la naturaleza |