| CHAPITRE XV L'ÈRE PROTÉROZOTIQUE | Página de portada | CHAPITRE XVII LA PÉRIODE ORDOVICIENNE (SILURIEN INFÉRIEUR) |
[ p. 476 ]
¶ Formations et histoire physique
Les grands mouvements de la croûte terrestre qui ont mis fin à l’ère protérozoïque ont transformé en terre une vaste zone située à l’intérieur des limites du continent nord-américain. La répartition géographique des strates basales du Cambrien,[1] le plus ancien système de l’ère paléozoïque, en témoigne. Lorsqu’elle est accessible, la base du système est, dans la plupart des endroits, discordante sur les formations sous-jacentes. La répartition des parties successives du système donne une idée des relations entre la mer et la terre tout au long de cette période, car la plupart des strates sont d’origine marine, comme le montrent leurs fossiles.
Les subdivisions du Cambrien et leur distribution
Le système cambrien a été divisé en trois séries, appelées respectivement Cambrien inférieur, moyen et supérieur. D’autres noms sont parfois attribués à ces divisions. Ainsi, Géorgien (Vermont), Acadien et Potsdam (Saratogan) (New York), noms de localités où les divisions correspondantes du Cambrien ont été différenciées pour la première fois en Amérique du Nord, sont synonymes (en Amérique) respectivement de Cambrien inférieur, moyen et supérieur.
Le Cambrien inférieur. Les formations du Cambrien inférieur ne sont connues en Amérique du Nord qu’à proximité des frontières est et ouest du continent (Fig. 357). À l’est, on les trouve dans la ceinture des Appalaches et à certains endroits plus à l’est ; à l’ouest, on les trouve dans divers États entre les 110e et 120e méridiens. À l’est comme à l’ouest, les strates contiennent des fossiles marins. Les strates de l’est se sont accumulées dans des détroits, des bras de mer, etc., plutôt que sur les rivages de la haute mer.
[ p. 477 ]
[ p. 478 ]
On pense que la vaste étendue comprise entre les Appalaches, d’une part, et l’ouest du Montana et de l’Utah, d’autre part, était une étendue de terre au début de la période. De cette terre, des sédiments étaient probablement charriés vers la mer de part et d’autre. Ces sédiments, ultérieurement cimentés en roches, ont formé une partie des schistes et des grès du Cambrien inférieur.
La discordance entre le Cambrien inférieur et sa base représente ce que l’on appelle parfois un intervalle « perdu ». Cette appellation n’est pas tout à fait appropriée, car la discordance témoigne d’une période d’exposition et d’érosion, suivie d’une submersion et d’un dépôt.
Le Cambrien moyen. Les strates de la série moyenne (acadienne) du système cambrien se rencontrent avec celles du Cambrien inférieur ; on les trouve également au Texas, en Oklahoma, en Arizona, dans certaines parties du Montana et peut-être ailleurs. Comme les couches du Cambrien moyen contiennent des fossiles marins, leur répartition indique que le continent était envahi par la mer venant du sud avant la fin du Cambrien moyen. Comme pour la série précédente, les couches du Cambrien moyen sont absentes d’une grande partie de l’intérieur, si les identifications actuelles sont correctes. Là où le Cambrien moyen repose sur le Cambrien inférieur, les deux sont généralement concordants[2]. Là où le Cambrien moyen chevauche le Cambrien inférieur, il est discordant sur les formations plus anciennes.
Le Cambrien supérieur. Au Cambrien tardif (époque de Potsdam ou Saratogan), la mer recouvrait une plus grande partie du continent, car la série de Potsdam couvre non seulement les frontières est et ouest du continent, mais aussi une grande partie de l’intérieur. Le Cambrien supérieur est concordant avec le Cambrien moyen à l’est et à l’ouest, mais à l’intérieur, il repose en discordance sur des formations précambriennes. La Fig. 358 montre une partie de la distribution du système cambrien dans son ensemble (voir explication sous la figure).
Grande submersion durant le Cambrien. La distribution des différentes séries du système montre que le grand événement physique de [ p. 479 ] la période cambrienne en Amérique du Nord fut la submersion progressive du continent. Théoriquement, cette submersion peut avoir été provoquée par une montée du niveau de la mer ou par un abaissement du niveau des terres, ou par les deux à la fois. L’abaissement du niveau des terres et la montée du niveau de la mer peuvent être dus à la gradation, au diastrophisme, ou aux deux combinés.
La gradation, cause possible de submersion. La gradation est perpétuelle et inévitable là où il y a terre et mer. Les vagues attaquent la terre le long de ses frontières, et les agents de dégradation des terres abaissent sa surface. La première cause directe d’empiètement de la mer sur la terre, et la seconde une cause indirecte, puisque tous les sédiments transportés de la terre vers la mer déplacent un volume d’eau égal et élèvent d’autant la surface de la mer. Aussi faible soit-elle pendant une brève période, elle a pour effet de faire avancer la mer sur la terre ; et l’abaissement simultané des terres par la dégradation augmente la surface de cette avancée. Si ce phénomène se poursuit suffisamment longtemps, le creusement des côtes le long des frontières des terres, s’abaissant sur toute la surface, et l’élévation concomitante du niveau de la mer, entraîneront inévitablement la couverture des continents par l’eau et la propagation de dépôts sur tous les continents, sauf les derniers vestiges, à condition qu’aucune déformation du corps terrestre ne se produise entre-temps.
On a calculé que si la Terre, dans son état actuel, restait sans déformation suffisamment longtemps pour que les continents soient nivelés, le dépôt des sédiments ainsi dérivés de la mer élèverait le niveau de la mer d’environ 208 mètres. Cela submergerait une grande partie des terres nivelées. Les preuves de la gradation au Cambrien sont claires et solides. La plupart des sédiments qui composent le système rocheux cambrien ont été érodés et déposés dans la mer. Cela a abaissé les terres et soulevé la mer. La gradation a donc été un facteur dans la submersion du continent, et il existe des preuves que de grands progrès ont été réalisés vers le nivellement de la base de l’Amérique et d’autres continents, avant la fin du Cambrien. Le nivellement de la base u_, et donc en soi, plaide en faveur de l’idée qu’aucune déformation importante n’a affecté le continent pendant son déroulement. Conformément à cette hypothèse, il n’existe aucune preuve directe de déformation profonde au cours de la période cambrienne. Il existe donc une présomption d’absence de diastrophisme important au cours de cette période.
Si la gradation était le seul facteur impliqué dans la submersion des terres, l’avancée de la mer aurait dû être régulière, bien que pas nécessairement constante à tout moment et en tout lieu. Sans entrer dans les détails, il semble certain que des changements dans les zones de dépôt, autres que ceux attribuables à la gradation, ont eu lieu, mais aucun de ces changements n’implique de déformations prononcées comme celles observées dans les roches des époques protérozoïque et archéozoïque.
La déformation comme cause de submersion. Les déformations qui provoquent la submersion des terres (et l’émergence des fonds marins) peuvent être soit superficielles, impliquant les roches jusqu’à quelques kilomètres de profondeur au maximum, soit profondes, impliquant les roches à des profondeurs beaucoup plus importantes. On peut citer une ou deux phases de déformation.
1. Extension latérale ou fluage des continents. Les continents se situent à environ 4 500 mètres au-dessus du fond océanique. Leur poids exerce une pression de 6 800 à 9 000 kg par pouce carré sur leur base. Cette pression tend à provoquer l’expansion des continents par fluage dans les bassins océaniques, selon le même principe que l’expansion d’une grande masse de glace, telle qu’une calotte glaciaire. Cet élargissement est contré par la pression hydrostatique des océans sur les parois des plateformes continentales. Cette pression est d’environ 2 260 kg par pouce carré au fond, de sorte qu’il subsiste une pression déséquilibrée de 4 500 à 6 800 kg par pouce carré, tendant à provoquer le fluage. Est-ce suffisant pour vaincre la résistance de la roche, qui s’oppose au fluage ? Même la plus faible de ces valeurs équivaut à la résistance à l’écrasement de certaines roches plus fragiles, et représente un pourcentage notable de la résistance à l’écrasement même des plus résistantes. Sous une pression inférieure, on observe souvent un fluage de la roche dans les mines. Il n’est donc pas improbable qu’une telle pression, exercée de façon constante pendant une période prolongée, puisse provoquer un certain élargissement des grandes plates-formes continentales, et donc (1) un abaissement de leur surface, (2) une submersion de leurs bords, et (3) simultanément une élévation du niveau de la mer. De nombreux phénomènes, qui ne peuvent être cités ici, semblent étayer cette hypothèse du fluage latéral[^3], mais son efficacité n’est pas déterminée.
[ p. 481 ]
2. Ajustements entre les segments continentaux et océaniques, une cause possible de submersion. Il a été démontré [3] récemment qu’il existe un plan à environ 160 kilomètres sous la surface de la Terre où la pression vers le bas, sous les zones continentales sus-jacentes, est égale à la pression vers le bas sous les zones océaniques sus-jacentes, malgré le fait que les surfaces des continents se trouvent, en moyenne, à 5 kilomètres au-dessus du fond océanique. La raison en est que la roche sous les océans est plus dense que celle sous les continents. À tous les horizons au-dessus de ce plan isostatique, à une profondeur d’environ 160 kilomètres, une colonne de roches continentales moyennes mesurée vers le bas à partir de la surface terrestre, pèse plus qu’une colonne similaire de roches sous-océaniques. Cela tend à favoriser le mouvement d’expansion mentionné ci-dessus, et donc à provoquer la submersion des terres. Un état de choses similaire prévalait probablement à l’époque cambrienne. Il apparaît cependant que des régions particulièrement élevées, comme la Cordillère, se situent plus haut qu’elles ne le seraient si elles étaient en équilibre isostatique.[4] Ce déséquilibre peut donner lieu à des mouvements particuliers visant à assurer un équilibre plus parfait, et donc à des déformations régionales ou locales. Au cours de toute grande période de déformation, comme celle de la fin du Protérozoïque, il est probable que certaines portions de la croûte ont été poussées au-dessus d’une position d’équilibre, puis ont eu tendance à se tasser. D’autres portions ont pu être abaissées en dessous de la position d’équilibre, puis ont eu tendance à s’élever. De tels mouvements seraient distribués de manière asymétrique et pourraient entraîner un gauchissement lent et silencieux, mais inégal.
3. Autres ajustements, comme causes possibles de submersion. Divers autres changements et ajustements de la surface des différentes parties de la lithosphère, tels que le déchargement des continents par l’érosion et le chargement des bassins océaniques par le dépôt, les déversements de lave et les ajouts ou pertes inégaux de chaleur dans différents endroits, peuvent également avoir contribué à provoquer la submersion ou l’émergence des terres.
Base de la subdivision du Cambrien
Nous devons maintenant rechercher les moyens par lesquels les strates du Cambrien [ p. 482 ] peuvent être reconnues, et en outre les moyens par lesquels le Cambrien inférieur peut être distingué du Cambrien moyen, et le Cambrien moyen du Cambrien supérieur.
Superposition. Lorsqu’une formation est concordante avec une autre d’âge connu, comme le Cambrien moyen avec la formation inférieure, la présomption est forte que la formation supérieure s’est formée après l’inférieure, sans interruption. Dans ce cas, l’âge approximatif de la formation supérieure est connu. En revanche, lorsqu’une formation est discordante avec une autre d’âge connu, les relations stratigraphiques entre elles ne déterminent pas l’âge de la formation supérieure, si ce n’est qu’elle est généralement plus récente que l’inférieure.
Fossiles. Le Cambrien est le plus ancien système rocheux connu pour contenir d’abondants fossiles. La plupart d’entre eux représentent des coquilles ou d’autres parties dures d’animaux marins enfouis dans les sables et les boues au moment de leur dépôt. Les fossiles des strates de chaque division du Cambrien constituent la faune connue de cet étage, bien qu’il ne faille pas supposer que des fossiles de toutes les espèces ayant vécu aient été préservés.
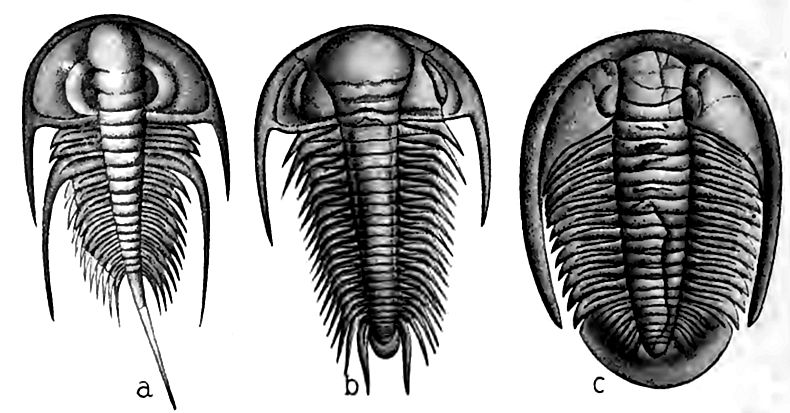
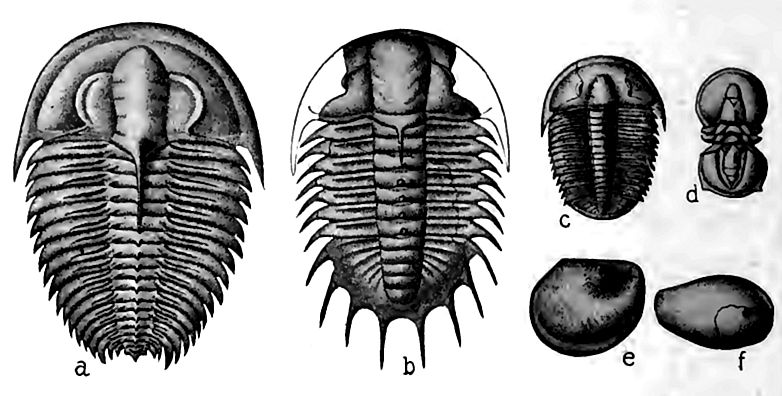
La série du Cambrien inférieur contient certains fossiles caractéristiques. Parmi eux se trouvent des espèces d’un genre de trilobites connu sous le nom d’Olenellus (Fig. 359, a). Outre les représentants de ce genre, on trouve de nombreuses autres espèces de divers types de vie. Dans l’ensemble, le nom de faune d’Olenellus a été donné, et Olenellus Cambrian est synonyme de Cambrien inférieur et de Géorgien. Il ne faut pas en conclure que des représentants du genre Olenellus se trouvent partout au Cambrien inférieur[5], ni que d’autres genres de trilobites y sont absents.
[ p. 483 ]
Là où des formations représentant l’ensemble de la période sont présentes, les fossiles des couches supérieures ne sont pas identiques à ceux des couches inférieures. En règle générale, aucun changement d’espèce n’est notable sur aucun plan particulier ; au contraire, dans les couches successivement plus hautes, certaines espèces trouvées en dessous disparaissent, et de nouvelles espèces apparaissent, comme pour les remplacer. Ces variations montrent que les habitants de la mer ont évolué au fil du temps. À peu près au même stade du système cambrien où le genre Olenellus disparaît, le genre Paradoxides (Fig. 359, b) apparaît des deux côtés du bassin nord-atlantique. Les autres espèces associées aux Paradoxides sont quelque peu différentes de celles du genre Olenellus. Les Paradoxides et leurs associés constituent la faune des Paradoxides, une faune qui comprend également de nombreuses espèces d’autres genres de trilobites, ainsi que de nombreuses espèces d’autres classes d’animaux non apparentés aux trilobites. De l’avis général, le Cambrien moyen, des deux côtés de l’Atlantique Nord, est défini par la faune à Paradoxides, de sorte que Paradoxides Cambrian est synonyme de Cambrien moyen et d’Acadien (p. 476). Dans la partie occidentale de l’Amérique du Nord, ainsi que de l’autre côté du Pacifique Nord, le Cambrien moyen ne contient pas de Paradoxides, mais des Olenoides, et sa faune est connue sous le nom de faune à Olenoides, mais elle n’en est pas moins distincte de la faune du Cambrien inférieur.
De même, aux faunes de Paradoxides et d’Olenoides succède une autre, connue sous le nom de faune de Dikellocephalus (359, c), qui caractérise les strates cambriennes situées au-dessus du Cambrien moyen. Les géologues s’accordent à définir le Cambrien supérieur comme la série de strates contenant la faune de Dikellocephalus.
Il ne faut pas comprendre que chaque espèce de la faune de Paradoxides diffère de chaque espèce de la faune d’Olenellus ci-dessous et de la faune de Dikellocephalus ci-dessus. Ce n’est pas le cas ; mais tant d’espèces des trois faunes sont différentes, qu’avec un nombre considérable d’espèces à partir desquelles juger, leur distinction est possible pour ceux qui connaissent les fossiles cambriens.
Pour distinguer ces faunes, une analogie avec les animaux vivants est suggérée. Les faunes actuelles d’Amérique du Nord et d’Amérique du Sud sont raisonnablement distinctes ; mais il ne s’ensuit pas qu’elles n’aient pas d’espèces communes. Étant donné un seul animal, même un expert pourrait ne pas être en mesure d’affirmer qu’il provient d’un continent ou de l’autre, bien que cela soit possible pour certaines espèces ; mais si l’on dispose d’un grand nombre d’animaux de l’un ou l’autre continent, il est possible de déterminer à quel continent, c’est-à-dire à quelle faune géographique, ils appartenaient. Il en va de même pour les différentes faunes cambriennes. Elles ont des espèces en commun, et ces espèces ne distinguent pas les groupes de strates qui les contiennent. Cependant, certaines espèces ne se trouvent que dans la partie inférieure, d’autres seulement dans la partie moyenne, et d’autres encore seulement dans la partie supérieure du système, et ces espèces servent à distinguer les principales divisions.
Séquence des faunes basée sur la stratigraphie. Il ne faut pas comprendre que les roches qui contiennent de telles faunes soient classées ensemble simplement parce qu’elles contiennent certains fossiles. Ce n’est pas la raison, ou du moins pas la principale, de les regrouper. L’ordre de séquence des faunes est d’abord déterminé par la superposition des strates. La faune du Cambrien inférieur ne pourrait être considérée comme plus ancienne que celle du Cambrien moyen si les couches contenant la première ne se trouvaient pas sous-jacentes à celles contenant la seconde. En d’autres termes, la base principale de la corrélation au moyen de fossiles est la stratigraphie.
Sédimentation au Cambrien
Français La sédimentation au Cambrien semble avoir suivi les lois générales qui régissent le dépôt dans les périodes d’absence relative de grands mouvements de déformation, et donc de nivellement progressif de la base. La plupart des sédiments cambriens connus se sont déposés dans la mer, et leur superficie peut être considérée comme une mesure approximative de la superficie de la mer à cette époque. La sédimentation était probablement plus rapide au début de la période, lorsque la superficie terrestre [ p. 485 ] était la plus grande et la plus élevée, et plus lente aux stades ultérieurs, après que la surface terrestre eut été usée par l’érosion et rétrécie par l’empiètement de la mer. La sédimentation était probablement plus importante près des terres, et moins loin des côtes en eau profonde.
Sources et types de sédiments. Comme lors des autres périodes géologiques, les sédiments d’origine terrestre provenaient de toutes les formations alors exposées à l’érosion. Les sédiments situés le long des limites immédiates du territoire étaient sans doute différents de ceux déposés plus loin. Même le long du littoral, on observait des variations considérables, à la fois en raison des variations d’origine des sédiments et des différences d’action des vagues, des rivières et des courants.
Les formations cambriennes comprennent toutes les phases courantes des roches sédimentaires. On y trouve des conglomérats, vraisemblablement accumulés près des côtes à cette époque ; des grès, dont le sable s’est déposé en eaux peu profondes, où les vagues étaient suffisamment vigoureuses pour empêcher la sédimentation ; des schistes argileux représentant des dépôts effectués en eaux calmes ou plus profondes ; et des couches de calcaire représentant, pour la plupart, des accumulations de coquillages, etc., là où les sédiments terrigènes n’étaient pas charriés en quantité.
Variations géographiques des sédiments. La répartition de ces diverses sortes de roches sédimentaires montre que divers types de couches détritiques s’accumulaient en différents endroits au même moment, et au même endroit à des moments différents. De plus, leur accumulation s’est faite à des rythmes très différents. Ainsi, la totalité du Cambrien moyen (tout ce qui s’est déposé pendant toute la période du Cambrien moyen) semble être présente en de nombreux points, mais l’épaisseur des strates du Cambrien moyen est loin d’être uniforme. Des épaisseurs de roche égales ne s’accumulent pas nécessairement sur des périodes de temps égales.
Le fait que, dans l’intérieur nord des États-Unis, la formation du Cambrien supérieur soit généralement constituée de grès, et que ce grès soit largement répandu, indique que l’eau était si peu profonde lors de son dépôt que les vagues pouvaient transporter le sable sur de longues distances. De plus, la structure des strates, avec leur stratification entrecroisée (Fig. 266), leurs rides, etc., montre que l’ensemble de la série épaisse, du bas vers le haut, s’est déposé dans des eaux peu profondes [ p. 486 ], et donc sur une surface qui s’est progressivement abaissée par rapport au niveau de la mer, à mesure que les sédiments s’accumulaient. La plus grande proportion de calcaire (principalement de la dolomie) dans le Cambrien supérieur de l’intérieur sud et sud-est suggère des mers plus claires, mais peut-être pas profondes. Les terres adjacentes étaient peut-être trop basses pour fournir des sédiments abondants. Le calcaire constitue également une part importante du Cambrien supérieur des Rocheuses[6], bien que les roches clastiques prédominent plus à l’ouest. Là où le Cambrien supérieur est calcaire, il n’est généralement pas nettement différencié de l’Ordovicien sus-jacent.
Répartition et affleurements du système cambrien
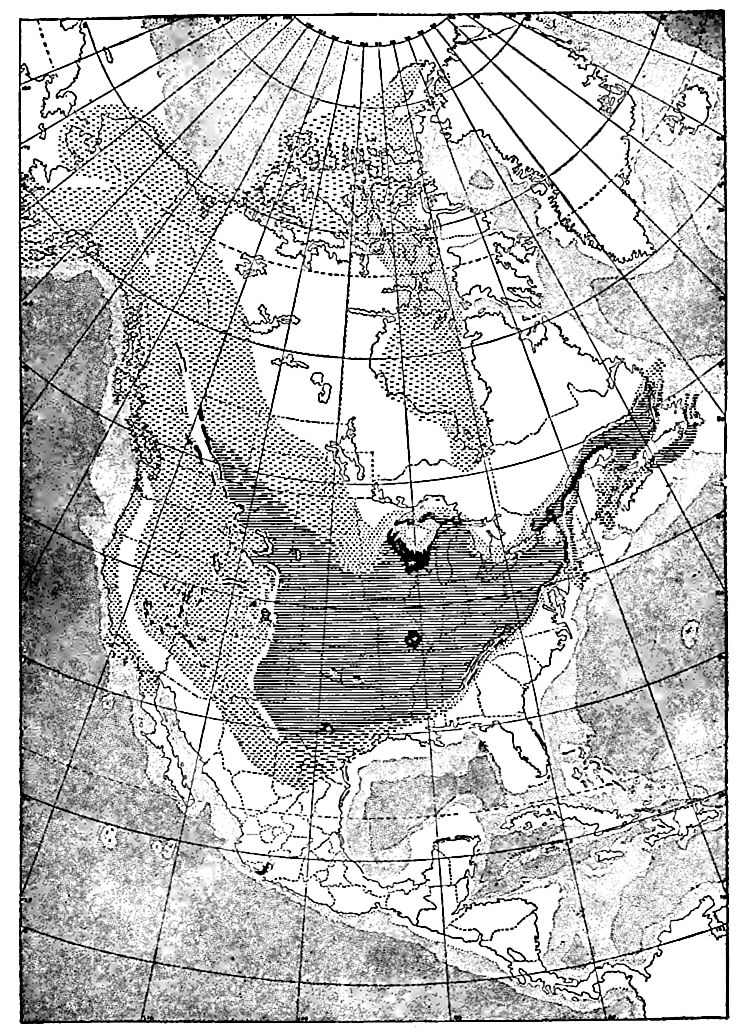
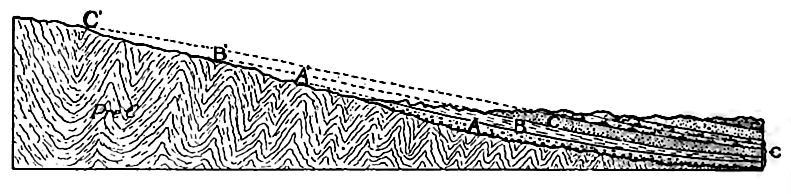
Les formations cambriennes étaient autrefois aussi étendues que les mers cambriennes elles-mêmes, mais elles ne sont plus présentes aujourd’hui sur toute la zone qu’elles couvraient autrefois. Les bords exposés des strates ont subi l’érosion, de sorte que la limite du système telle qu’elle apparaît aujourd’hui autour des zones de roches précambriennes n’est plus sa limite originelle et ne représente pas le littoral de la mer cambrienne lorsque ses eaux étaient les plus étendues. Fig. 360 illustre les conditions qui prévalent souvent. Chacune des formations cambriennes, représentées par A, B et C, s’étendait autrefois plus à gauche.
[ p. 487 ]
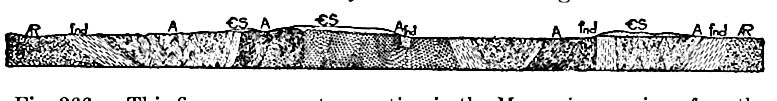
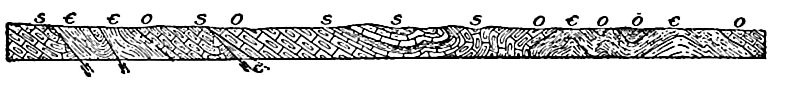
Les zones où les formations cambriennes sont exposées ne doivent pas être confondues avec les zones où elles existent réellement. Les formations cambriennes sont exposées, par exemple, dans le Wisconsin, le Missouri et le Texas ; mais les strates du Texas sont sans doute continues, sous des formations plus récentes, avec celles exposées dans le Missouri, et celles du Missouri avec celles du Wisconsin, et celles-ci avec celles des Black Hills à l’ouest, et avec celles de l’État de New York à l’est (Fig. 361).
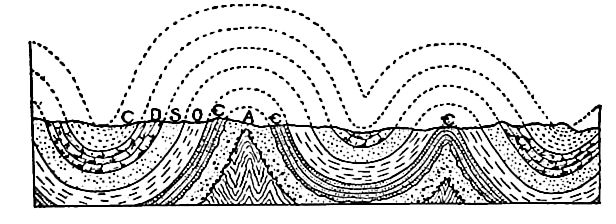
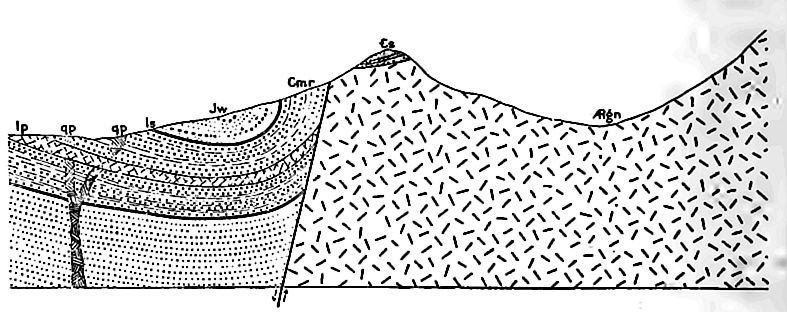
Position des affleurements. La carte (Fig. 358) montrant les zones où le système cambrien est maintenant exposé, révèle plusieurs points importants. (1) . De nombreux affleurements sont associés à des affleurements des systèmes archéen et protérozoïque (Fig. 341). À certains endroits, le Cambrien exposé se trouve le long de la frontière des parties exposées de ces systèmes plus anciens, d’un seul côté, tandis qu’à d’autres, il les entoure complètement. Cette distribution n’est pas particulière au Cambrien, mais est caractéristique de la plupart des formations par rapport à celles plus anciennes. (2) Les zones exposées du Cambrien dans les Appalaches [7] se présentent en ceintures parallèles ou subparallèles (Fig. 358). Ceci est le résultat (a) du plissement auquel les strates cambriennes et ultérieures de cette région ont été soumises, et (b) de l’érosion que les plis ont subie. Fig. 362 aidera à expliquer la répétition des affleurements. Dans ce diagramme, A représente les strates précambriennes, Ç représente le Cambrien, et O, S, D et C les systèmes ordovicien, silurien, dévonien et carbonifère, respectivement. Après le plissement des strates, l’érosion a coupé les plis. Un pli qui implique des couches cambriennes, s’il est tronqué sous le niveau du fond de ces couches à leur point le plus élevé, expose deux ceintures de strates cambriennes, une de chaque côté d’un axe précambrien, comme représenté dans la partie gauche de la figure. Si la troncature se situe à un niveau inférieur au sommet et supérieur au bas du Cambrien (côté droit de la Fig. 362), les strates de ce système sont exposées en une seule ceinture le long de la crête du pli. (3) . Par endroits, les affleurements cambriens sont entourés de formations plus anciennes. Dans ce cas, les affleurements cambriens représentent vraisemblablement des vestiges ayant échappé à l’érosion. Ils pourraient occuper des dépressions à la surface de formations précambriennes, ou constituer des collines (Fig. 363).

[ p. 488 ]
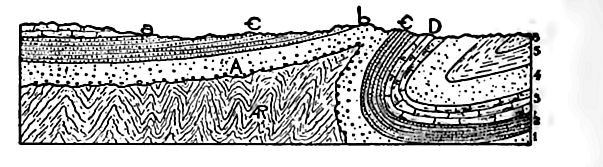
Largeur des affleurements. Les affleurements continus les plus étendus du Cambrien (Fig. 358) se trouvent dans le Wisconsin ; pourtant, seul le Cambrien supérieur y est présent, avec une épaisseur de moins de 300 mètres, tandis que dans les Appalaches, où le système a une épaisseur totale de plusieurs milliers de pieds, il apparaît à la surface en bandes étroites. Autrement dit, les affleurements sont étroits à l’est, où le système est épais, et larges à l’intérieur, où il est mince. L’explication de cette apparente anomalie réside principalement dans l’attitude des strates. Dans le Wisconsin, elles sont presque horizontales, tandis que dans les régions montagneuses, à l’est comme à l’ouest, elles sont inclinées, souvent à des angles prononcés. Lorsque les strates sont verticales, la largeur de leur affleurement sur une surface horizontale est à peu près égale à l’épaisseur des lits (Fig. 364) ; là où elles sont presque horizontales, comme dans la partie gauche de la Fig. 364, la largeur de l’affleurement sur une surface horizontale est beaucoup plus grande.
[ p. 489 ]
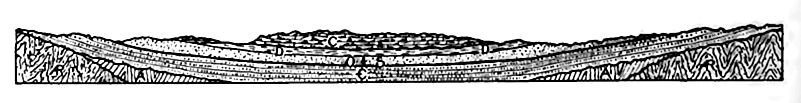
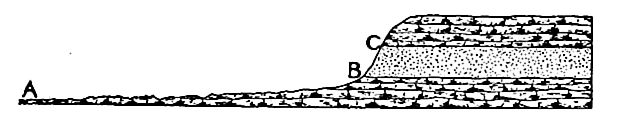
Il ne faut cependant pas en déduire que les strates horizontales présentent nécessairement un large affleurement. La largeur de l’affleurement est également influencée par la topographie, comme le montre la Fig. 365. Ici, la strate horizontale entre B et C a à peu près la même épaisseur que •€ de la Fig. 364, mais son affleurement est étroit. En général, la largeur de l’affleurement, dans la mesure où elle est déterminée par la topographie, dépend de l’angle entre les plans de stratification et la surface où la formation affleure. La largeur de l’affleurement diminue à mesure que cet angle augmente.
Évolution des sédiments cambriens depuis leur dépôt. Les sédiments du système cambrien ont subi plus ou moins de modifications depuis leur dépôt. Dans la plupart des régions, les graviers, les sables et les boues ont été compactés et cimentés en conglomérats, grès et schistes respectivement. À certains endroits, la cimentation du grès a même conduit à sa transformation en quartzite.
[ p. 490 ]
Sur de vastes zones de l’intérieur (Missouri, Wisconsin, Texas, etc.), les strates restent horizontales ou presque, tandis que dans d’autres régions, elles ont été inclinées, plissées et faillées. Là où des plis serrés se sont produits, les roches ont été plus ou moins métamorphisées. Dans les cas extrêmes, les grès ont été transformés en schistes quartzeux, les schistes en ardoises et schistes, et les calcaires en marbre. La Fig. 361 montre la position générale des strates cambriennes (Ç) sur une grande partie de l’intérieur, et les Fig. 367 et 368 illustrent leur position et leurs relations là où elles ont été plissées et faillées.
Fin du Cambrien. Aucun changement physique de grande importance ne semble avoir marqué la fin du Cambrien en Amérique. Nulle part sur notre continent, à notre connaissance, de montagnes ne se sont formées à cette époque, et nulle part de vastes étendues de fonds marins ne se sont transformées en terres, bien que des discordances locales[8] entre ce système et le suivant témoignent de changements locaux dans les sites de dépôt.
Dans d’autres continents
Europe.[9] En Europe, comme en Amérique du Nord, une déformation généralisée avant le début du Cambrien a transformé de vastes zones [ p. 491 ] du continent actuel en terres, et il existe des preuves que ces terres, comme celles de l’Amérique, ont été soumises à une érosion prolongée avant le dépôt du système cambrien, car il est généralement discordant sur les strates plus anciennes.
Les formations cambriennes d’Europe, comme celles d’Amérique, sont principalement clastiques. Une part considérable du matériel qu’elles contiennent est grossière, et les strates sont souvent ondulées, affectées par des stratifications entrecroisées et des fissures solaires, caractéristiques qui montrent qu’une grande partie des sédiments cambriens se sont déposés en eaux peu profondes, et certains d’entre eux là où ils n’étaient pas continuellement recouverts d’eau.
Au Pays de Galles (Cambria), pays d’où le système tire son nom, l’épaisseur du système atteint 3 600 mètres ou plus. Cette épaisseur considérable est égalée, voire dépassée, en Bretagne. En Scandinavie, en revanche, où les Cambriens inférieur, moyen et supérieur sont tous présents, l’épaisseur globale ne dépasse parfois pas 120 mètres. En Russie occidentale également, elle est mince. Ces différences signifient probablement que les sédiments se déposaient à certains endroits beaucoup plus rapidement qu’à d’autres. Elles signifient probablement aussi que les sédiments étaient parfois répartis sur des plateaux sous des eaux peu profondes, et parfois sur des fonds en pente où les couches étaient inclinées. Le Cambrien moyen est beaucoup plus étendu que le Cambrien inférieur ou supérieur, ce qui montre que des changements dans les relations entre la mer et la terre étaient en cours pendant la période cambrienne, déplaçant les zones d’érosion et de sédimentation.
Les strates cambriennes d’Europe occidentale ont été fortement plissées depuis leur dépôt. En Europe centrale et orientale, en revanche, elles sont essentiellement horizontales. On y trouve des couches d’argile encore plastiques et des couches de sable encore non cimentées.
Aucun changement géographique de grande importance ne semble avoir marqué la fin du Cambrien. À cet égard, comme à d’autres, les histoires cambriennes de l’Europe et de l’Amérique du Nord concordent.
Autres pays. Les roches cambriennes se trouvent dans diverses régions de Sibérie, de Chine, d’Inde, d’Australie et de Tasmanie, ainsi que dans la partie nord-ouest de l’Argentine, mais leur répartition en dehors de l’Amérique du Nord et de l’Europe est mal connue.
[ p. 492 ]
Formations glaciaires. (1) Dans les environs du fjord de Varanger, dans le nord de la Norvège, à 70° 8’ N. de latitude, se trouve une couche de roches à boules (les couches de Gaisa)[10] reposant sur un pavage lisse et strié de type glaciaire distinctif. Les couches de Gaisa reposent sur la surface érodée d’un terrane cristallin et ont été considérées comme appartenant à la partie la plus ancienne du système cambrien, ou comme antérieures à celui-ci. (2) Des explorations récentes en Chine[^12] ont montré l’existence, sur le fleuve Yangtsé, à 30° de latitude, d’une épaisse formation (170 pieds) de roches à boules d’origine glaciaire, contenant de nombreuses boules striées de diverses sortes de roches (Fig. 369). La formation glaciaire se situe ici à la base du Paléozoïque et sous la série qui porte les trilobites cambriens.

Français Des formations glaciaires d’à peu près le même âge ont été trouvées en Australie, et peut-être en Afrique du Sud.[11] L’interprétation la plus probable, avec les connaissances actuelles, est que ces formations à cuvettes de Norvège, de Chine et d’Australie (Fig. 370) appartiennent soit à la période de transition qui a accompagné et suivi [ p. 493 ] la déformation qui a fermé le Protérozoïque, soit aux premiers stades du Paléozoïque, antérieurs au Cambrien démontré. La profonde signification climatique de ces formations glaciaires est évidente. Le témoignage des fossiles cambriens, d’autre part, implique des conditions climatiques presque uniformes dans toutes les régions où des fossiles ont été trouvés, et la large extension de la mer pendant la dernière partie de la période semblerait indiquer des climats océaniques plutôt que continentaux à cette époque.


Durée de la période cambrienne
Il n’existe aucun moyen d’estimer avec certitude la durée du Cambrien. La destruction et l’évacuation vers la mer de volumes de roche aussi importants que ceux représentés par les sédiments du Cambrien ont nécessité une très longue période ; mais comme il n’existe pas de rythme standard d’accumulation de sédiments, quel qu’il soit, cette longue période ne peut être réduite à des années. On estime que le calcaire se forme parfois à un rythme de 30 cm par siècle. Dans certaines régions de l’Ouest, on trouve 1 800 mètres de calcaire, en plus d’épais massifs de roches fragmentaires. Au rythme d’accumulation indiqué ci-dessus, ces 1 800 mètres de calcaire nécessiteraient une période de 600 000 ans, et si l’on prenait en compte le temps des autres formations de la même région, la période serait considérablement allongée. Il convient toutefois de rappeler que si un pied par siècle peut être le rythme d’accumulation du calcaire, il ne s’ensuit pas que ce soit le rythme de formation des calcaires cambriens. Les données sur lesquelles repose cette estimation sont considérées comme trop élevées, et une accumulation moins rapide impliquerait une période de temps proportionnellement plus longue.
De nombreuses estimations du temps géologique, fondées sur diverses données, ont été tentées[12]. Appliquées au Cambrien, ces estimations attribuent généralement à cette période une durée de 1 à 3 millions d’années. Il convient toutefois de garder à l’esprit que l’intérêt principal de ces chiffres est de souligner la longue durée de cette période.
¶ La vie du Cambrien
Aucun événement de l’histoire de la Terre ne présente peut-être un intérêt plus grand que la première apparition de la vie ; mais la date de son apparition est inconnue. Il existe de bonnes preuves que la vie existait avant la fin de l’ère archéozoïque et, selon l’hypothèse de l’accrétion, il n’est pas improbable que son apparition ait précédé de loin les plus anciennes formations archéennes accessibles. Si tel est le cas, il est tout à fait impossible que les premières formes de vie soient jamais connues à partir de leurs vestiges. Les traces de plantes et d’animaux à l’ère protérozoïque sont si indirectes, obscures ou rares qu’elles ne donnent qu’une conception très incomplète de la vie avant le Cambrien. Les informations fournies par les fossiles imparfaits indiquent qu’ils ne racontent même pas l’essentiel de l’histoire de la vie protérozoïque, car la vie représentée par les fossiles n’aurait pu exister sans d’autres formes de vie non représentées.
Le Cambrien est le système le plus ancien pour lequel on dispose de traces raisonnables de vie, et même ici, ces traces sont loin d’être complètes. Le règne animal est assez bien représenté parmi les fossiles, mais les restes végétaux sont difficilement identifiables.
La rareté des fossiles végétaux. L’existence des plantes au Cambrien serait peut-être mise en doute si l’on ne savait pas que tous les animaux en dépendent, directement ou indirectement, pour leur alimentation. Il faut donc supposer que les plantes abondaient. L’inadaptabilité des plantes inférieures à la fossilisation est sans doute la principale explication de leur faible représentation parmi les fossiles cambriens. Des raisons d’ordre physique ont déjà été avancées pour penser que la surface terrestre était recouverte d’une certaine forme de végétation. Pourtant, il n’existe aucune trace identifiable de plantes terrestres, et seulement d’obscures traces de plantes marines. La leçon à tirer est l’extrême imperfection des archives fossiles.
Les fossiles d’animaux
Chaque grande division du règne animal, à l’exception des vertébrés, avait ses représentants au Cambrien. Les Arthropodes (p. 945) étaient représentés par des crustacés ; les Mollusques par des gastéropodes, des ptéropodes et des pélécypodes ; les Molluscoïdes par des brachiopodes ; les Vermes par des annélides ; les Échinodermes par des cystoïdes ; les Cœlentérés par des graptolites, des méduses et des coraux ; les Porifères par des éponges ; et les Protozoaires par des rhizopodes. Tous les représentants de ces groupes parmi les fossiles cambriens semblent être marins. Il n’existe aucune trace d’animaux terrestres, mais cela ne prouve pas qu’ils n’aient pas existé. Bien qu’aucun reste de vertébré n’ait encore été découvert, il serait téméraire de supposer qu’aucun d’entre eux n’a existé.
Arthropodes. Parmi les Arthropodes, seuls les crustacés (représentés aujourd’hui par les écrevisses, les crabes, etc.) ont été trouvés dans les strates cambriennes. Leurs représentants étaient les trilobites et les entomostracés. Les trilobites étaient de loin les formes de vie les plus distinguées dans les mers cambriennes. Ils étaient non seulement les plus organisés, mais aussi les plus caractéristiques de la période. Leurs genres successifs distinguent le mieux ses stades successifs, et leur répartition est un moyen essentiel de corréler les formations de différents continents et de différentes provinces d’un même continent, comme indiqué précédemment (p. 481). Ils sont éteints depuis longtemps. Les figures 359 et 372 montrent leurs trois lobes longitudinaux (d’où leur nom) et leurs trois divisions transversales : tête, thorax et bouclier caudal. Les trilobites étaient très avancés en termes de développement, possédant presque tous les systèmes anatomiques et fonctions physiologiques des crustacés modernes. Leurs yeux composés, formés de nombreux œillets, constituent peut-être le meilleur indicateur de leur développement. À cette époque et aux suivantes, le nombre d’œillets dans les yeux des trilobites variait d’une vingtaine à plusieurs milliers. Certains trilobites du Cambrien, cependant, n’avaient pas d’yeux, tandis que d’autres possédaient des rudiments avortés, ce qui suggère que leurs ancêtres en avaient possédé. L’acquisition et l’avortement d’un organe aussi important semblent indiquer une variation des conditions de vie. Cela pourrait se résumer à une migration vers des eaux profondes et sombres, ou à l’habitude de s’enfouir dans la vase, où les yeux sont devenus inutiles. Les yeux étaient souvent légèrement surélevés sur des lobes en croissant, la face convexe tournée vers l’extérieur. Plus tard, ces croissants sont devenus de plus en plus courbés, élargissant le champ de vision d’avant en arrière, à l’avantage évident du trilobite.
[ p. 497 ]
La surface supérieure du corps était ornée de diverses granulations, d’épines et d’autres marques, dont la signification est mal comprise. Ces ornementations variaient au fil du temps, augmentant généralement jusqu’à ce que l’apogée des trilobites soit passée. Les trilobites possédaient une rangée de membres fins et articulés de chaque côté, ainsi que de délicats filaments servant d’organes respiratoires. La nature de leurs membres indique que les trilobites marchaient et nageaient. Ils possédaient des antennes qui servaient sans doute d’organes tactiles, et ils muaient leur coquille à différents stades de croissance, comme les crabes modernes. Sans plus de détails, il convient de noter qu’à cette époque reculée, une organisation très complexe et bien différenciée avait été acquise, possédant presque tous les organes et fonctions des arthropodes actuels. Les Molluscoidea. Cet embranchement était bien représenté par les brachiopodes (lampes-coquilles, Fig. 373). En termes d’importance géologique, les brachiopodes de cette période étaient surpassés par les trilobites ; mais contrairement à ces derniers, les brachiopodes sont encore vivants et sont des représentants remarquables de la stabilité et de la persistance. Bien que les espèces et la plupart des genres aient changé, la classe dans son ensemble n’a subi que peu de modifications depuis le Cambrien. À l’époque, comme aujourd’hui, les valves d’une division du groupe étaient articulées, tandis que celles d’une autre ne l’étaient pas. Une division formait des coquilles de phosphate de calcium, et une autre de carbonate de calcium.

[ p. 498 ]
Mollusques. Les céphalopodes (coquilles logées), la classe la plus élevée de mollusques, se trouvent dans les couches supérieures du Cambrien. Comme ils étaient déjà très développés à cette époque, il ne fait guère de doute que cette classe ait connu une longue histoire avant la fin de la période.[13] Les pélécypodes (bivalves, 6,Fig. 374) ont vécu dès le début de la période, bien que leurs restes soient peu abondants. Les gastéropodes (univalves, c, d, e,Fig. 374) étaient plutôt abondants tout au long de la période. Les formes primitives sont principalement de type conique bas, tandis que les formes plus amplement enroulées et spiralées sont devenues courantes plus tard. La ressemblance étroite de certains d’entre eux avec les gastéropodes modernes mérite d’être soulignée.

[ p. 499 ]
Vermes et échinodermes. Les vers marins ont laissé des traces de leur abondance par des forages, des traces, etc. (Fig. 375). Quelques cystoïdes, précurseurs des magnifiques crinoïdes (lys de pierre), étaient présents, mais aucun crinoïde fossile n’a été découvert.
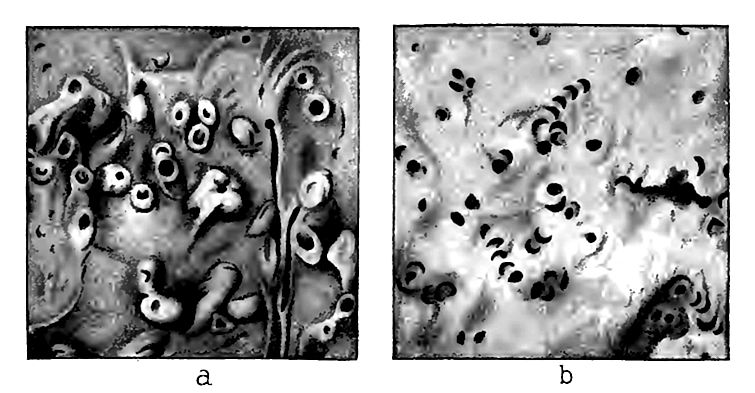
Cœlentérés. Les cœlentérés étaient représentés par des hydrozoaires (graptolites et méduses) et des anthozoaires. Nulle part ailleurs les bizarreries de la fossilisation ne sont mieux illustrées qu’ici. Des vestiges de graptolites, parmi les formes animales les plus délicates, et de méduses, parmi les plus tendres, ont été préservés, tandis que certaines formes, beaucoup moins facilement détruites, ont laissé peu de traces. Les graptolites, aujourd’hui éteints, étaient des organismes élancés, en forme de panache, constitués d’une série de* cellules indurées, dans lesquelles vivaient les zooïdes individuels, attachés à un axe commun élancé qui unissait la colonie. La colonie entière semble avoir flotté librement dans la mer (Fig. 376, e). Le secret de leur préservation réside probablement dans le fait que, étant des formes flottantes, elles se sont souvent installées dans des eaux calmes et assez profondes au large, où de fins limons s’accumulaient et où les conditions étaient favorables à un enfouissement sans destruction. Le cas de fossilisation le plus singulier est la préservation de traces de méduses, ou du moins de celles qui sont ainsi identifiées. Celles-ci sont illustrées dans la Fig. 376, c et d. Leurs empreintes ont été trouvées au Cambrien inférieur. On trouve également des fossiles obscurs de coraux (Fig. 376, a et b), dont les formes ressemblent tellement à des éponges qu’elles ont longtemps été considérées comme telles. Les coraux semblent avoir été plus abondants dans d’autres régions du monde qu’en Amérique du Nord.[14]

Types inférieurs. Les éponges étaient présentes en abondance tout au long de cette période. Il est probable que de nombreuses formes simples et inférieures, classées comme protozoaires, aient existé, mais seules quelques formes identifiables ont été découvertes.
Vie implicite. L’existence d’une vie animale aussi abondante implique une vie végétale abondante pour fournir la nourriture nécessaire, comme indiqué précédemment. De plus, diverses caractéristiques des fossiles suggèrent la présence d’animaux inconnus des fossiles. Un pourcentage important des animaux cambriens connus étaient dotés de coquilles, de tests, de plaques ou d’autres formes de revêtements durs. Il s’agit généralement de dispositifs de protection, impliquant des ennemis ou des rivaux contre lesquels une protection était nécessaire. La caractéristique la plus significative de ces dispositifs de protection réside peut-être dans le fait qu’ils sont généralement du même type que ceux des animaux similaires des époques ultérieures. Les coquilles des gastéropodes, des pélécypodes et des brachiopodes ne diffèrent de celles d’aujourd’hui que par des caractéristiques mineures. Les revêtements des trilobites ressemblaient beaucoup à ceux de leurs parents actuels, et on peut en dire autant de presque toutes les formes. Si le caractère de leurs ennemis ou rivaux avait radicalement changé, on pourrait s’attendre à une évolution notable des dispositifs défensifs. Il est donc naturel d’en déduire que les conflits qui régnaient dans les mers cambriennes étaient similaires à ceux d’aujourd’hui. On peut pousser l’hypothèse plus loin et en déduire que les conflits qui ont conduit à l’évolution des dispositifs défensifs étaient très semblables à ceux qui ont marqué la période de leur maintien.
Stade d’évolution représenté. Quel stade d’avancement de la vie avait déjà été atteint au début du Cambrien ? Les fossiles du système indiquent-ils que la vie de cette période était primitive, ou impliquent-ils qu’elle avait largement dépassé les formes primitives ? La réponse à ces questions se trouve (1) dans l’estimation du degré de développement des diverses structures et fonctions organiques, et (2) dans l’ampleur de la divergence des types animaux.
(1) À titre de comparaison, on peut supposer que les formes de vie primitives étaient aussi simples que les formes existantes les plus simples. Il existe une multitude de plantes et d’animaux constitués d’une seule cellule, et si l’on considère qu’elles représentent la forme la plus proche des formes primitives existantes, dans quelle mesure la vie cambrienne les avait-elle dépassées ?
Nous ne sommes pas entièrement réduits à présumer que les premières formes de vie animale étaient beaucoup plus simples que celles du Cambrien, car les stades de développement des jeunes de certains animaux cambriens révèlent quelque chose de leur histoire ancestrale. C’est une loi bien établie de l’embryologie que les animaux, aux premiers stades de leur développement, passent par une succession de changements dans lesquels leur structure ressemble à celle de leurs ancêtres à maturité ; en d’autres termes, l’histoire individuelle de tout animal est un résumé de l’histoire de ses ancêtres. Or, les trilobites du Cambrien sont connus pour avoir subi une série de changements remarquables après que les individus se soient suffisamment développés pour être fossilisés, et on en déduit qu’ils ont traversé d’autres stades auparavant. Il existe donc des raisons spécifiques de croire qu’ils avaient une longue lignée d’ancêtres.
Sur le plan anatomique et physiologique, il est clair que la quasi-totalité des organes fondamentaux étaient développés. On y trouvait des systèmes squelettiques de diverses formes, des systèmes musculaires, des systèmes nerveux très développés, comme l’indiquent les yeux et autres organes sensoriels, des dispositifs de capture et d’ingestion de nourriture, des organes de digestion, de sécrétion, d’excrétion et de respiration ; en bref, pratiquement tous les grands systèmes anatomiques et physiologiques que possèdent aujourd’hui les animaux. Les animaux du Cambrien avaient acquis les diverses habitudes de vie des animaux actuels de leur espèce, ainsi que les divers modes de survie.
(2) Les études des dernières décennies ont convaincu les chercheurs que les formes de vie ultérieures ont dérivé des formes antérieures par des processus d’évolution dont la méthode exacte n’est pas entièrement comprise ; mais le fait de cette dérivation n’est pas aujourd’hui considéré comme une question ouverte. À mesure que les diverses formes se sont développées et ont divergé d’une souche ancestrale commune, de nombreuses formes intermédiaires ont disparu, et les formes divergentes se sont ainsi largement séparées. Par divergence continue, avec la perte des types intermédiaires, une série de formes de plus en plus discontinue s’est développée, et les branches qui ont survécu sont devenues de plus en plus distinctes. Ce processus n’est pas sans rappeler l’évolution de la cime d’un arbre, où la disparition de la plupart des branches intérieures laisse quelques grandes branches portant les branches les plus nombreuses et les plus récentes, tandis que celles-ci portent à leur tour les rameaux les plus hauts et les plus externes représentant la phase vivante. On pense ainsi que la divergence actuelle des organismes en règnes, branches, classes, ordres, familles, genres, espèces et variétés s’est établie.
Si l’on suppose que l’ensemble du système des êtres vivants dérive d’une forme primitive commune ou de quelques formes primitives, une comparaison de l’état primitif avec le degré de divergence à l’époque cambrienne donnera une idée.
[ p. 503 ]
une évaluation de l’ampleur de l’évolution déjà accomplie. Si l’on y ajoute une comparaison entre la vie cambrienne et celle d’aujourd’hui, on peut estimer l’ampleur relative de l’évolution avant et depuis la période cambrienne.
Français Il est à noter que non seulement tous les sous-règnes animaux, sauf peut-être les vertébrés, étaient présents, mais que, dans beaucoup d’entre eux, les formes avaient fini par avoir si près de l’aspect des formes vivantes que les classes et certains ordres sont facilement reconnaissables. L’initiation et la divergence des structures et des types qui ont précédé l’étape cambrienne signifient beaucoup plus en termes d’évolution que toute l’évolution des temps ultérieurs. Formulée en termes numériques, nous pouvons peut-être dire que 60 à 90 pour cent de l’évolution représentée par la vie d’aujourd’hui a été accomplie à l’époque précambrienne.
Développement mental. Les guerres des animaux cambriens, illustrées par leurs armes d’attaque et de défense, ne peuvent guère être exemptes d’un développement mental notable, quelle que soit la nature de ce développement. Que les trilobites cherchaient leur nourriture ou poursuivaient leurs proies par la vue et se guidaient par le toucher, est suggéré par leurs yeux et leurs antennes, et il est difficile de concevoir des sens fonctionnels sans les processus mentaux qui accompagnent généralement la poursuite et la capture.
Adaptations écologiques. La répartition des fossiles cambriens indique qu’à cette époque, comme depuis, la vie s’est adaptée à son environnement physique. Il semble y avoir eu des zones de vie côtière, de vie en mer et de vie en eaux profondes, bien que les preuves de cette dernière soient rares. Ces variations doivent être prises en compte dans la comparaison et la corrélation des faunes, car des différences géographiques considérables peuvent exister entre des faunes strictement contemporaines.
Provinces zoologiques. Les assemblages de la vie de cette période semblent avoir varié de manière plus large, donnant naissance aux provinces zoologiques. Il est probable que les principaux facteurs de développement de ces provinces furent des barrières qui isolaient, ou en partie, certaines parties de la mer des autres. Cette séparation a dû atteindre un degré tel que la vie de chaque région s’est développée selon ses propres lignes, plus ou moins indépendamment de l’évolution des autres régions.
[ p. 504 ]
Les faunes primitives du Cambrien étaient plutôt provinciales, mais vers la fin de cette période, à mesure que les mers s’étendaient sur les continents, une tendance marquée au cosmopolitisme se manifesta. Il ne faut pas croire que la vie était partout la même à un stade donné. Il y avait probablement moins de variations géographiques, durant la plupart des périodes géologiques, qu’aujourd’hui, bien qu’il ne soit pas certain que cela ait été le cas à toutes les époques passées. Il est improbable qu’il y ait jamais eu uniformité sur l’ensemble du globe.
La Succession des Faunes
Selon la doctrine de l’évolution, on présume que la vie de chaque stade passé est issue de celle qui l’a immédiatement précédée et qu’elle a fusionné avec celle qui l’a immédiatement suivie. On suppose généralement qu’en l’absence d’influences exceptionnelles, le processus s’est accompagné d’une série continue de changements lents, sans lignes de démarcation nettes. Si cette conception se réalisait, il serait moins approprié de parler d’une succession de faunes que d’une faune continue en constante évolution. Il n’est cependant pas encore démontré que l’évolution se soit faite uniquement par de très légers changements survenant de génération en génération. Elle a pu se dérouler par des changements distincts et abrupts[15]. Cette doctrine, telle qu’elle est actuellement admise, ne soutient pas qu’une faune entière soit susceptible de se transformer brutalement en une autre, mais simplement que de nouvelles espèces pourraient y apparaître brutalement. Indépendamment de cette hypothèse ou de toute autre hypothèse spécifique, il convient de noter que les archives géologiques, telles que connues actuellement, ne montrent pas de gradations complètes d’une espèce à une autre. Dans certains cas, il existe une approximation proche d’une série graduée d’une espèce à une autre, mais les étapes de la gradation ne sont pas suffisamment proches et définies pour décider entre l’évolution par un nombre infini de petits changements et un nombre plus restreint de grands changements.
Si l’on passe des espèces aux faunes, il est évident qu’il faut adopter un point de vue plus général. L’observation montre que, dans certains cas, une faune évolue progressivement vers la suivante, tandis que dans d’autres cas, le changement semble brutal. Si l’on pouvait étudier la progression de la vie à travers le monde comme une unité, il apparaîtrait probablement qu’il y a eu une gradation presque parfaite de la vie d’un stade à celle du suivant. Cette gradation a probablement eu lieu plus rapidement à certaines époques qu’à d’autres, et il est tout à fait certain que certaines formes ont changé beaucoup plus vite que d’autres. Mais si l’on limite notre étude à la succession des faunes sur un continent ou une province, il est évident que la progression de l’évolution dans la région étudiée a été interrompue par des changements physiques qui ont affecté la profondeur, la température ou la clarté de l’eau, ainsi que la nature du fond, et que ces changements ont entraîné des variations dans le caractère et la répartition de la vie. De ces influences locales superposées au progrès général de la vie sont nées des périodes bien définies de changements notables, entre lesquelles les faunes ont conservé un caractère relativement constant, bien que subissant toujours des modifications. Lorsque le changement faunique au sein d’une série cohérente est brutal et qu’il n’existe aucune trace de hiatus dans les archives, l’explication est généralement à rechercher dans la migration, par exemple lorsqu’une nouvelle faune arrive d’une autre région, submergeant l’ancienne faune. L’ensemble du processus est étroitement analogue à la succession bien connue des races humaines provoquée par les migrations humaines.
Dans l’étude du progrès de la faune, il y a donc lieu de reconnaître (1) des changements plutôt brusques provoqués par des invasions écrasantes ; (2) des changements moins brusques provoqués par l’entrée plus progressive d’espèces extérieures et le mélange progressif des immigrants avec les espèces résidentes ; (3) des changements très progressifs dus à l’évolution lente des espèces résidentes lorsqu’elles ne sont pas beaucoup affectées par l’immigration ou par des changements physiques ; et (4) une évolution plus rapide due à des changements profonds dans les conditions physiques ou à d’autres agents moins bien compris.
L’apparition brutale de la faune cambrienne. L’explication de l’apparition soudaine et apparente de la faune cambrienne est l’une des questions ouvertes de la géologie. D’une manière générale, on peut dire que les formations plus anciennes ont été soumises au métamorphisme, ce qui a eu pour effet de détruire leurs fossiles ; mais cette suggestion n’est pas tout à fait adéquate, car certaines formations plus anciennes ne sont pas profondément modifiées et semblent tout à fait adaptées à la préservation [ p. 506 ] des fossiles. De plus, des fossiles sont parfois conservés dans des formations plus récentes, fortement perturbées et altérées. Il est également vrai que certaines formations plus récentes, qui semblent bien adaptées à recevoir et à conserver des empreintes organiques, en sont dépourvues. Les géologues ont tendance à attribuer la rareté des fossiles précambriens, et donc l’apparition brutale et apparente de la faune cambrienne, à des conditions défavorables à la fossilisation à l’époque précambrienne, combinées à des modifications ultérieures de la roche. Cela fait de cette brusquerie une question de dossier plutôt qu’un fait avéré.
Français : Travail cartographique._ Les folios suivants de l’US Geological Survey fournissent de bonnes cartes _pour l’étude de la stratigraphie et des relations stratigraphiques du système cambrien dans différentes parties des États-Unis. Les textes des folios donnent un compte rendu de l’histoire physique des différentes régions : Alabama, Gadsden ; Arizona, Clifton ; Colorado, Anthracite-Crested Butte ; Géorgie, Rome ; _Maine, Penobscot Bay ; Massachusetts, Holyoke ; Michigan, Menominee ; Montana, Little Belt, Fort Benton ; New Jersey, Franklin Furnace ; Caroline du Nord, Mount Mitchell, Nantahala et Pisgah ; Oklahoma, Tishomingo ; Tennessee, Maynardville et Morristown ; Utah, Tintic ; Virginie-Occidentale, Bristol, Harper’s Ferry, Tazewell ; Wyoming, Absaroka (Crandall Sheet), Bald Mountain-Dayton, Cloud Peak-Fort McKinney et Sundance.
| CHAPITRE XV L'ÈRE PROTÉROZOTIQUE | Página de portada | CHAPITRE XVII LA PÉRIODE ORDOVICIENNE (SILURIEN INFÉRIEUR) |
¶ Remarques
[^3] : Chamberlin et Salisbury, Histoire de la Terre, Vol. II.
[^12] : Willis, Recherches en Chine, vol. II.
Un résumé de la littérature sur le Cambrien nord-américain avant 1892 est donné par Walcott dans Bull. 81, US Geol. Surv. ↩︎
Ulrich et Schuchert pensent que le synclinorium des Appalaches du Cambrien précoce a été largement drainé à la fin de cette époque. Bull. 52 (Paléontologie 6) NY Mus. Rept. du paléontologue d’État, 1901, p. 636. ↩︎
Hayford, Washington Acad, of Sci., Vol. VIII, 1906, pp. 25-40. ↩︎
Putnam et Gilbert, Bull. Phil. Soc. Washington, Vol. XII, pp. 31-75, et Gilbert, Jour. Geol., Vol. Ill, p. 351. ↩︎
Walcott, lour, de Geol., Vol. XVII, 1909. ↩︎
Dawson, Bull. Geol. Soc. Am., Vol. XII, pp. 64-68. ↩︎
Voir les folios de l’US Geol. Surv. ↩︎
Cushing. Bull. Geol. Soc. Am., Vol. XIX, p. 155. ↩︎
Le meilleur résumé, en anglais, du Cambrien de l’Europe, se trouve dans le Geikie’s Text-book of Geology, 4e éd., Vol. II. ↩︎
Reusch, Geological Survey of Norway : La géologie de la Norvège du Nord, 1891. Également Strahan, Quar. Journal. Géol. Soc., Vol. LIII, 1897, pp. 137-146. ↩︎
David, Rapport du Congrès géologique international de Mexico, 1907; et Howchin Quar. Jour. Geol. Soc, Vol. LXIV, p. 234, 1908. ↩︎
Pour une discussion générale de cette question, voir Williams’ Geological Biology, Chap. II. ↩︎
Ulrich rapporterait les couches contenant les Céphalopodes à l’Ozarkien, un système qu’il ferait inclure la partie supérieure du Cambrien et la partie inférieure de l’Ordovicien, comme on le classe habituellement. ↩︎
Howchin, Quar. Jour. Geol. Soc. Vol. LXIV, p. 237. ↩︎
DeVries Die Mutationstheorie, 1903. Voir aussi Material! for the Study of Variation de Bateson, 1894 ; et W.B. Scott, On Variations and Mutations, Am. Jour. Sci., 1894, p. 355. ↩︎