| CHAPITRE XVII LA PÉRIODE ORDOVICIENNE (SILURIEN INFÉRIEUR) | Página de portada | CHAPITRE XIX LA PÉRIODE DÉVONIENNE |
¶ Formations et histoire physique
Les changements physiques qui ont mis fin à l’Ordovicien ont également marqué le début du Silurien. Ces changements comprenaient (1) des mouvements affectant intensément de petites zones, et (2) des mouvements affectant légèrement de vastes zones. Du point de vue de l’histoire continentale, ces derniers étaient les plus importants. Ces changements se sont sans doute produits lentement et, après leur apparition, la superficie des terres en Amérique du Nord a atteint une superficie jamais atteinte depuis le début du Cambrien. Cette augmentation des terres a entraîné un allongement des cours d’eau et, vraisemblablement, une érosion accrue.
Si la répartition des terres et des eaux au début du Silurien pouvait être définie avec précision, cela permettrait également de définir les zones où la sédimentation marine la plus ancienne de la période a eu lieu et, de manière générale, les zones où la sédimentation a été rapide et celles où elle a été lente, car alors, comme toujours, le taux de sédimentation devait être en relation plus ou moins définie avec les rivages. On peut supposer sans risque qu’au début du Silurien, des couches de sédiments clastiques s’accumulaient aux confins immédiats des terres, et aussi loin que les vagues et les courants pouvaient transporter d’abondants détritus, et qu’ailleurs les sédiments d’origine organique étaient relativement plus importants. Bien que la sédimentation ait été interrompue dans les régions émergées de la mer lors de la transition de l’Ordovicien au Silurien, cette interruption n’était pas universelle, et les strates siluriennes sont localement concordantes avec l’Ordovicien sur le continent, et plus généralement, on peut le supposer, dans les bassins océaniques.
[ p. 537 ]
Comme le système précédent, le Silurien peut être divisé en trois séries principales, comme suit :[1]
Chacune de ces séries est subdivisée en plusieurs formations, mais les subdivisions d’un endroit ne s’accordent pas avec celles d’un autre. Un bref aperçu de la nature et de la répartition de ces principales subdivisions du système offre un aperçu de l’histoire du continent durant cette période.
Silurien de l’Est
La série Oswegan. Les principales subdivisions de cette série à l’est sont l’Oneida et la Medina. Toutes deux apparaissent à la surface au sud du lac Ontario, et peut-être dans la partie occidentale des Appalaches plus au sud.[2] L’Oneida est constituée de conglomérats et de grès, et la Medina de grès et de schiste. Les sédiments de ces formations semblent avoir été déposés dans une mer intérieure peu profonde, comme le montrent leurs fossiles, ainsi que les stratifications entrecroisées, les rides, etc., qui affectent ses couches. La formation de Medina s’étend plus à l’ouest que l’Oneida, atteignant l’est de l’Ohio et de l’Ontario. Sa distribution, comparée à celle de l’Oneida, suggère un affaissement de l’intérieur oriental au début du Silurien. Les deux formations sont probablement continues sous des strates plus jeunes sur des zones considérables au sud du lac Ontario et de la vallée de MohawTk, et à l’ouest des Appalaches.[3] On ne sait rien des sédiments qui s’accumulaient [ p. 538 ] le long de la frontière orientale des Appalaches à cette époque, ni de ceux autour des autres régions terrestres.
La série du Niagara. La formation de Clinton recouvre la Medina en concordance, mais sa distribution est plus large. Vers l’ouest, elle s’étend jusqu’au lac Huron et à l’Indiana, et peut-être jusqu’au Mississippi. Si tel est le cas, elle est représentée par des couches classées avec le calcaire du Niagara. Cette formation est présente dans les Appalaches jusqu’en Alabama et en Géorgie, au sud. Les fossiles de cette formation dans les Appalaches sont si différents de ceux de l’intérieur qu’ils laissent supposer que les sites de sédimentation des deux régions n’étaient pas librement reliés. Des couches d’âge Clinton ont été identifiées en Nouvelle-Écosse et à quelques autres endroits au nord-est des États-Unis. La sédimentation marine y a probablement été continue pendant les périodes ordovicienne et silurienne.
Les variations de caractéristiques des couches Clinton selon les localités sont significatives. Dans les Appalaches, la formation est principalement constituée de grès et de schiste. Dans l’ouest de l’État de New York et plus à l’ouest, elle est en grande partie constituée de calcaire. La présence de calcaire ne signifie pas nécessairement que l’eau était profonde, mais plutôt qu’elle était si peu argileuse que les coquillages, etc., constituaient l’essentiel du dépôt. La vie coquillière peut être tout aussi abondante là où le sable et la boue s’accumulent qu’ailleurs, mais dans ce cas, le produit n’est pas du calcaire, mais du grès, du schiste, etc., contenant des coquillages. Des récifs de bryozoaires, semblables à des récifs coralliens, sont présents dans la formation de l’ouest de l’État de New York.[^4]
L’une des caractéristiques notables de la formation est sa teneur en minerai de fer, généralement sous forme d’hématite (Fe₂₂O₂₃). Ce minerai est souvent constitué de petites concrétions dont la taille et la forme ressemblent tellement à celles des graines de lin qu’elles ont suggéré le nom de minerai de « graine de lin ». Localement, il est également connu sous le nom de minerai « fossile » en raison de l’abondance de fossiles qu’il contient. Le minerai est connu en de nombreux points entre New York et l’Alabama, jusqu’au Wisconsin à l’ouest et jusqu’à la Nouvelle-Écosse au nord-est. Le minerai est interconnecté avec d’autres gisements de la formation et on pense généralement qu’il s’est accumulé par précipitation chimique dans des lagunes ou des vasières marécageuses.
Comme les formations Oneida et Medina qui les ont précédées, les couches Clinton [ p. 539 ] n’ont pas été identifiées dans la moitié occidentale du continent.
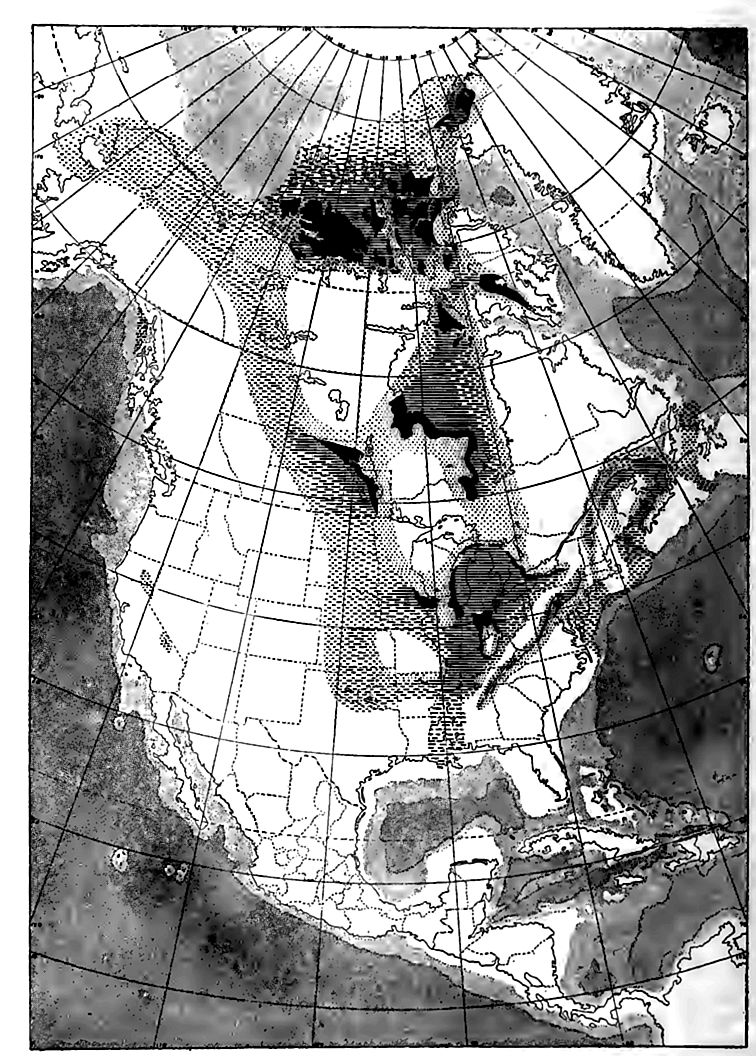
Français La formation de Clinton a été succédée par la formation de Niagara (subdivisée dans l’État de New York en schiste de Rochester, calcaire de Lockport et dolomite de Guelph, p. 537), qui s’étend plus à l’ouest que toutes les formations siluriennes précédentes, montrant que la submersion progressive des époques antérieures se poursuivait encore dans le bassin supérieur du Mississippi. Les chutes de la rivière Niagara se trouvent sur le calcaire de cette série (Fig. 113). Au nord du Missouri, la formation n’est pas connue loin à l’ouest du Mississippi, mais elle s’étend dans le Missouri, l’Arkansas et peut-être même jusqu’aux monts Arbuckle en Oklahoma. On la trouve également dans la région trans-Pecos au Texas. C’est à cette époque que la submersion du bassin supérieur du Mississippi a atteint son maximum, pour l’époque considérée. La limite sud de la mer intérieure est inconnue, mais elle semble avoir été séparée de l’océan dans cette direction par une barrière terrestre quelque part dans la région du Golfe (Fig. 397). Cette barrière se trouvait probablement au sud du Tennessee, car du calcaire du Niagara est présent dans la partie occidentale de cet État.
Une caractéristique importante de la répartition de la formation de Niagara est son fort développement aux hautes latitudes. On la trouve par endroits au Manitoba, à l’ouest de la baie d’Hudson, et en de nombreux points plus au nord, jusqu’au 80e parallèle. Ces endroits semblent être les vestiges d’une formation autrefois continue, et comme les fossiles sont globalement les mêmes et très semblables à ceux de l’Europe du Nord, on en déduit qu’il existait une liaison fluviale entre le bassin du Mississippi et l’Europe du Nord via les îles de l’Arctique, ce qui a permis la migration des espèces marines des eaux peu profondes des deux régions.
À l’est des Appalaches et à l’ouest du Mississippi, la répartition des strates du Niagara est mal connue. On les trouve probablement dans le New Hampshire et le Maine, ainsi que dans les provinces situées entre ces États et le Saint-Laurent. En Nouvelle-Écosse, les strates du Niagara sont représentées par du schiste, ce qui illustre une fois de plus l’accumulation simultanée de différentes sortes de roches dans différentes régions. L’équivalent exact de la formation du Niagara n’a pas été identifié avec certitude dans l’Ouest.
À l’ouest de New York, la formation est principalement calcaire. À l’exception du Trenton, aucune formation calcaire antérieure n’est peut-être aussi répandue. C’est la plus ancienne formation où des récifs coralliens bien développés ont été identifiés, bien que des polypes sécrétant des coraux aient vécu auparavant (p. 532). Ces récifs sont connus dans l’est du Wisconsin, dans l’Indiana et ailleurs.
[ p. 540 ]
[ p. 541 ]
Ces récifs et les dépôts adjacents illustrent, plus clairement que toute autre formation ancienne, l’origine de plusieurs variétés de calcaire. Les récifs eux-mêmes sont composés de vestiges mélangés de la vie qui s’y est développée. D’importantes masses de corail se dressent parfois dans la roche, telles qu’elles ont poussé, ayant échappé à la destruction lors de leur enfouissement dans le récif en croissance. Dans d’autres cas, les masses coralliennes dans le calcaire sont fragmentaires, brisées et usées par les vagues. Aux plus gros morceaux de corail s’ajoutent des détritus grossiers et fins, produits de la fragmentation du corail. Ces éléments se combinent pour former la masse de la roche récifale. Entre et autour des récifs, un mélange de fragments de coquillages et de coraux, ainsi que de sables calcaires, s’est accumulé, devenant plus fin à mesure que l’on s’éloignait du récif parent et que la pente du fond s’adoucissait. Ces matériaux se transformèrent finalement en boue calcaire, largement répandue sur le fond marin autour des récifs. Ce fin sédiment calcaire a finalement donné naissance à un calcaire blanc compact.
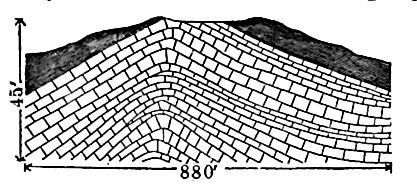
Épaisseur et structure. Contrairement aux formations précédentes du Silurien, le Niagara n’est pas plus épais à l’est qu’ailleurs. À l’est, en effet, où la formation est exposée, son épaisseur n’est que de 30 à 90 mètres, tandis qu’au Wisconsin, elle atteint un maximum de 240 mètres (en incluant peut-être une partie du Clinton), le tout étant constitué de calcaire. Bien que les couches du Niagara à l’intérieur soient en général presque horizontales, elles sont fréquemment bombées, ce qui leur confère un fort angle de pendage (Fig. 398). C’est le cas, par exemple, en divers points de l’extrémité sud du lac Michigan.
[ p. 542 ]
La série Cayugan (Salina). La formation de Salina, qui recouvre la formation du Niagara dans certaines parties de l’État de New York, de la Pennsylvanie, de l’Ohio, du Michigan et de l’Ontario, est beaucoup moins répandue et sa distribution limitée indique l’émergence d’une zone considérable dans le bassin du Mississippi à la fin de l’époque du Niagara.
La série Salina comprend plusieurs variétés de roches, dont des conglomérats, du grès, du calcaire, du schiste et du sel gemme. Ces formations sont associées à du gypse, accompagnement habituel des gisements de sel. Le schiste est le plus abondant et semble avoir pris naissance à la manière des schistes en général, mais la rareté de ses fossiles semble indiquer un dépôt dans des conditions défavorables à la vie.
Le sel est largement répandu. On le trouve en de nombreux points de l’État de New York, sur une superficie de 23 000 à 26 000 kilomètres carrés. Des couches isolées ont localement une épaisseur de 12 à 24 mètres. Plusieurs couches se superposent parfois, interstratifiées avec d’autres types de roches, et leur épaisseur totale atteint parfois 30 mètres. Près de Cleveland, quatre couches de sel, de 15 mètres d’épaisseur ou moins, sont interstratifiées avec 150 mètres de schistes. Par conséquent, à certains endroits, le sel représente une fraction considérable de l’épaisseur totale de la série.
Les couches de sel semblent impliquer l’existence de grandes lagunes ou mers fermées, dans lesquelles la série Salina s’est déposée. Si le climat de cette région avait été aussi humide qu’aujourd’hui, ces lagunes n’auraient pas pu être anormalement salées. Des incursions occasionnelles de la mer, apportant de nouvelles réserves d’eau salée, suivies de périodes où les lagunes étaient coupées de la mer et subissaient une évaporation rapide, semblent réunir les conditions nécessaires à la formation du sel. Il en serait de même pour une légère connexion continue avec la mer, de sorte que l’afflux d’eau de mer dans le bassin ne compense pas l’excès d’évaporation par rapport aux précipitations dans la zone du bassin. Une couche de sel de 12 mètres d’épaisseur implique l’évaporation de trois cents mètres d’eau de mer normale. Une grande partie du sel commercialisé en provenance de New York ne provient pas directement des couches de sel, mais des eaux des puits de sel.
Le calcaire de la Salina est en grande partie contemporain des schistes et des couches de sel. Il est plus épais là où ces couches sont minces, [ p. 543 ] et mince là où elles sont plus épaisses. Il contient peu de fossiles et on pense qu’il pourrait s’agir d’un précipité chimique. Ses relations avec les schistes et les couches de sel sont telles qu’elles indiquent que les zones d’accumulation des différents types de matériaux rocheux ont été déplacées de temps à autre, comme par de légers changements de niveau du sol ou de l’eau.
Au-dessus de la Salina proprement dite de l’État de New York, se trouve une mince série (45 mètres maximum) de calcaires, le Waterlime (Cobleskill, Rondout, Manlius, etc.), généralement considéré comme faisant partie du système silurien. Le nom Waterlime fait référence au fait que le calcaire est la source du ciment hydraulique, bien qu’il ne soit pas partout utile à cet effet, et que de nombreuses autres formations calcaires soient utilisées de la même manière. Le Waterlime est plus répandu que le Salina, s’étendant vers l’ouest à travers l’Ohio jusqu’à l’Indiana et le Wisconsin. Sa répartition et ses caractéristiques montrent que l’intérieur oriental était plus généralement submergé que lors du dépôt de la série salifère qui l’a précédé.
Dans le nord des Appalaches, on trouve une formation conglomérale (Shawangunk, NY, Green Pond Mountain, NJ), autrefois classée comme Oneida, et aujourd’hui considérée comme contemporaine de la série Cayugan. Jusqu’à récemment, on ne connaissait pas de fossiles dans ce conglomérat, mais leur découverte a déplacé la classification de la formation de la série Oswegan ancienne à la série Cayugan plus récente[^5]. Les matériaux de ce conglomérat sont principalement quartzeux et semblent provenir de terres situées à l’est. Ils ont été profondément indurés par la cimentation, ce qui confère à la formation une rigidité extrême. Les bords affleurants de ses lits inclinés constituent la crête de la chaîne de Kittatinny dans le New Jersey, et ses prolongements dans l’État de New York et en Pennsylvanie. Des roches siluriennes ont récemment été identifiées parmi les gneiss du Connecticut.
La formation d’Helderberg, autrefois considérée comme faisant partie du système silurien, est ici classée avec le Dévonien.
Silurien de l’Ouest
À divers endroits à l’ouest, on trouve une série de couches sédimentaires, pauvres en fossiles, entre l’Ordovicien connu en dessous et le Dévonien au-dessus. La nature des fossiles étant indécise, [ p. 544 ] l’âge de ces couches est sujet à caution. Certaines d’entre elles pourraient être siluriennes. Si le Silurien est réellement absent de toutes les régions où sa présence est inconnue aujourd’hui, il semblerait qu’une grande partie de l’ouest de l’Amérique du Nord était terrestre durant la période silurienne. Des couches siluriennes sont cependant connues en Californie du Sud, au Nevada, en Utah et en Alaska, et peut-être dans les Rocheuses canadiennes, et sa répartition pourrait être plus étendue qu’on ne le suppose.
Résumé
Comme dans tous les systèmes antérieurs du Paléozoïque, les plus fortes épaisseurs de strates siluriennes (estimées à environ 1 500 mètres, maximum) se trouvent dans la région des Appalaches. À l’intérieur, le système est relativement mince, mesurant des centaines de mètres plutôt que des milliers. Conformément à ces variations, le système est en grande partie constitué de sédiments clastiques provenant des eaux peu profondes de la ceinture appalachienne, tandis qu’à l’intérieur, il est principalement constitué de calcaire. Le site de sédimentation à l’est était une sorte de fosse (la fosse appalachienne), coupée de toute communication avec la mer intérieure, mais reliée à l’Atlantique, peut-être par l’actuelle région de Chesapeake. La plupart des sédiments de cette fosse ayant été déposés en eaux peu profondes, on pense généralement qu’ils indiquent que la fosse s’enfonçait à une vitesse comparable à celle de leur accumulation. Ce point de vue doit cependant être nuancé, comme suggéré page 461. Dans la mesure où l’affaissement a eu lieu, les épais sédiments pourraient en avoir été la cause, ou l’une des causes. Avec le gauchissement du creux, il pourrait y avoir eu un gauchissement vers le haut de la zone terrestre adjacente qui fournissait les sédiments.
L’histoire du Silurien, telle qu’on la comprend aujourd’hui, comprend : (1) une submersion générale de la partie orientale des États-Unis, à l’ouest des Appalaches, qui a entraîné une extension progressive de la mer jusqu’à la fin de l’époque nigérienne ; (2) un retrait partiel de la mer de cette même zone durant l’époque de Salina ; et (3) une extension de la mer à la fin de cette même époque. Il y a sans doute eu de nombreuses oscillations mineures de niveau qui n’ont pas été déterminées.
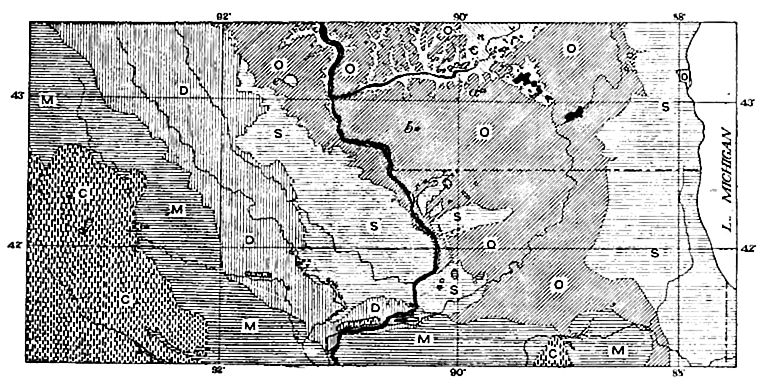
Étendue antérieure et stratigraphie générale. Les marges actuelles des différentes formations siluriennes ne sont pas leurs marges d’origine, [ p. 545 ] car leurs parties exposées ont subi l’érosion, et l’érosion des lits inclinés déplace leurs affleurements. Dans certaines localités, il existe des données permettant d’estimer quelque peu l’extension antérieure des formations. Dans le Wisconsin, par exemple, des vestiges (points aberrants, a, b, c,Fig. 399) du Niagara se trouvent bien au-delà du corps principal de la formation telle qu’elle existe actuellement. Ces points aberrants fixent au moins une limite minimale à l’extension originelle de la formation.
Roches ignées. Des roches ignées d’âge silurien ont été identifiées à quelques endroits en Amérique du Nord. Les formations siluriennes sont parfois affectées par des intrusions ignées, mais leur datation est généralement incertaine. On pense que certaines roches ignées du Nouveau-Brunswick sont d’âge silurien, et peut-être certaines de celles de la Nouvelle-Écosse et du Maine.[4]
Fin de la période. Les changements géographiques à la fin du Silurien étaient moindres que ceux à la fin de l’Ordovicien, et le système silurien est peut-être moins distinctement séparé du Dévonien au-dessus que de l’Ordovicien en dessous.
Climat et durée. Rien n’indique une grande diversité de température au Silurien, mais beaucoup suggère [ p. 546 ] que l’uniformité s’étendait à de grandes étendues de latitude, car les fossiles des régions tempérées chaudes sont en partie les mêmes que ceux des régions arctiques. Certaines régions semblent avoir été temporairement très arides. La période silurienne a peut-être été comparable en durée à l’Ordovicien.
Silurien étranger
En Europe, les strates siluriennes ont une distribution similaire à celle de l’Ordovicien, bien qu’elles soient absentes dans certaines régions où ces dernières sont présentes. Le fait que les strates siluriennes n’apparaissent pas à la surface sur de vastes zones n’indique pas leur absence générale, mais plutôt leur dissimulation généralisée. Dans la majeure partie du nord de l’Europe, hors Grande-Bretagne, le système a été peu déformé. Dans la partie sud du continent, les formations siluriennes n’apparaissent que par petites zones, au milieu de formations plus anciennes. Contrairement aux roches siluriennes de la province nord, celles du sud sont très déformées.
Les formations siluriennes d’Europe, en particulier celles de la province du nord, sont plus largement composées de calcaire que celles de l’Ordovicien, ce qui suggère des mers plus claires.
Des changements géographiques ont eu lieu en Europe à la fin de la période, comme le montre la discordance entre les systèmes silurien et dévonien dans certains endroits (Grande-Bretagne et Irlande), bien que la conformité soit la règle.
L’Ordovicien et le Silurien des autres continents n’ont pas été généralement distingués. Les équivalents des deux systèmes, tels qu’ils sont distingués en Europe et en Amérique du Nord, se retrouvent probablement dans tous les continents moins connus.
¶ La vie silurienne
Le retrait massif de la mer de la surface de l’Amérique du Nord à la fin de l’Ordovicien a réduit la superficie des eaux peu profondes disponibles pour la vie qui y vivait. L’évolution répressive sévère qui a suivi a été la principale caractéristique biologique de la transition de l’Ordovicien au Silurien. Avec la réinvasion de l’intérieur par la mer du Silurien moyen, il s’en est suivi une évolution expansionniste de la faune des eaux peu profondes, qui constitue la principale caractéristique biologique du milieu de la période. Vers la fin de la période, il y a eu une autre restriction de la mer épicontinentale, compliquée par une salinité intense dans la région intérieure orientale, et il s’en est suivi une seconde évolution répressive par laquelle la faune est passée au type dévonien.
Théoriquement, l’histoire de la vie terrestre aurait dû être l’inverse de celle de la mer ; car à mesure que la mer se contractait, la terre s’étendait, et l’évolution expansionniste de la vie terrestre aurait dû aller de pair avec l’évolution restrictive de la vie marine. C’était probablement le cas, mais les archives de la vie terrestre sont trop rares pour le démontrer. Dans la mesure où le climat était aride, il était défavorable à une vie terrestre abondante.
La transition de l’Ordovicien
Il n’existe que peu de données sur la vie en eaux peu profondes du Silurien inférieur. La côte orientale du continent se trouvait alors loin des limites de la plateforme continentale, et les dépôts qui s’y trouvent sont enfouis et inaccessibles. La frontière occidentale a peut-être été submergée, mais la faune y est peu connue.
Outre la réduction de la superficie propice à la vie en eaux peu profondes, les conditions étaient probablement moins favorables, zone par zone, qu’auparavant, car les apports terrestres ont vraisemblablement augmenté. L’augmentation des détritus apportés à la mer a probablement inhibé certaines formes de vie, en a nui à d’autres et n’en a favorisé que quelques-unes. Certains bassins et baies étaient sans doute trop doux, d’autres trop salés, et la salinité de certains d’entre eux a pu varier défavorablement. Ces considérations générales peuvent expliquer la rareté des faunes des premières strates du Silurien. Mais les conditions n’étaient pas défavorables partout. Dans le golfe du Saint-Laurent, les espèces ordoviennes ont survécu plus ou moins longtemps et se sont mêlées aux espèces siluriennes au fur et à mesure de leur développement, marquant ainsi la transition. Ce site semble avoir été le berceau de l’une des phases provinciales de la faune silurienne, mais il est peu probable qu’il ait été le seul. La principale faune silurienne de l’intérieur ne semble pas provenir de celle de la frontière atlantique, mais s’est développée quelque part au nord, ou a migré depuis l’Europe par une route vers le nord.
Le stade expansionniste et la faune du Silurien moyen
À mesure que la mer s’étendait lentement sur le continent vers le milieu de la période, l’espace croissant et les conditions plus favorables à la plupart des formes de vie des eaux peu profondes ont entraîné une évolution expansionniste qui a donné naissance à la faune du Niagara. Les familles et les classes étaient sensiblement les mêmes qu’à l’Ordovicien, mais la plupart des genres étaient nouveaux, ainsi que presque toutes les espèces. En général, il y a eu une avancée biologique, bien que cela ne soit pas vrai pour toutes les classes. Seuls les aspects les plus marquants des changements seront notés.
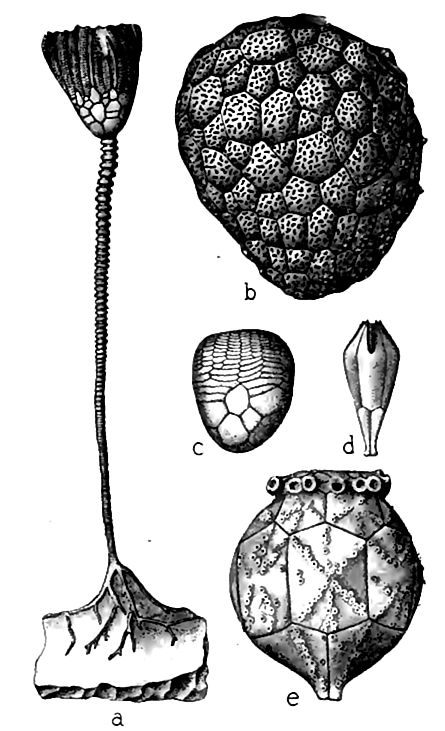
Les échinodermes. Un trait distinctif de la faune silurienne était le développement riche et varié des échinodermes, impliquant à la fois l’essor ou le déclin de formes antérieures et l’introduction de nouvelles. Le grand fait marquant de cette période, en lien avec les échinodermes, fut l’essor des crinoïdes. Ils atteignirent une telle abondance dans certaines localités propices que leurs fragments formaient la majeure partie du calcaire. Ces endroits étaient de véritables « parterres » de « lys de pierre », où des formes belles et variées poussaient en quelque sorte en bosquets. L’assemblage d’espèces à chacune de ces localités avait ses propres particularités, mais les genres étaient identiques ou similaires. Quelques crinoïdes siluriens sont représentés à la Fig. 400, mais un nombre limité de figures ne peut rendre justice à leur variété et à leur beauté. Malgré divers signes de progrès, nombre des caractères les plus primitifs subsistaient, indiquant que la classe n’avait pas encore atteint son apogée.
Les cystoïdes étaient encore abondants, et les véritables blastoïdes apparaissent maintenant pour la première fois. Les étoiles de mer semblent avoir peu progressé et n’avoir occupé qu’une place limitée dans la faune, et les étoiles-serpents et les échinoïdes en occupaient encore moins. Le lent développement de ces types, qui se sont imposés bien plus tard, reflète peut-être un fait général : l’évolution des grandes classes a été lente, aussi soudaine qu’elle puisse paraître. L’ascension des cystoïdes et des crinoïdes vers leur apogée se révélerait peut-être aussi lente que celle de leurs congénères, si l’on pouvait retracer leur histoire depuis ses origines. Une imperfection un peu plus marquée dans les archives fossiles aurait éliminé toute trace des étoiles-serpents et des oursins à ces époques reculées, et leur apparition en abondance à une époque ultérieure aurait semblé soudaine et remarquable.
Les brachiopodes. Les brachiopodes ont résisté aux vicissitudes du passage de l’Ordovicien au Silurien sans perdre leur prestige, [ p. 549 ] bien qu’ils aient subi un changement presque complet d’espèces et un très grand changement de genres. Si l’on considère que les brachiopodes étaient parmi les invertébrés les plus résistants et les plus conservateurs, ce grand changement d’espèces et de genres souligne la tension des conditions qui ont contrôlé la transition et son importance biologique. Les brachiopodes siluriens avaient gagné en différenciation et avaient réalisé des progrès notables en termes de structure. Dans l’ensemble, ils étaient plus robustes et présentaient des signes plus évidents de vitalité abondante qu’auparavant ; mais parallèlement à ces développements progressifs, des modifications rétrogrades ont été observées.
[ p. 550 ]
Les bryozoaires. Les bryozoaires coralliens ont beaucoup moins contribué aux calcaires siluriens qu’à ceux de la période précédente. Cela s’explique en partie par le déclin de cette classe, et en partie par le remplacement des types plus massifs par des formes plus délicates et fragiles.
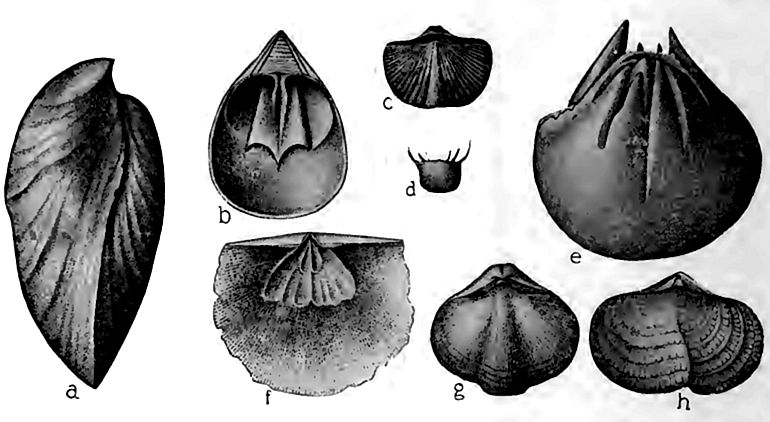
Les mollusques. Les céphalopodes semblent être restés les habitants les plus puissants des mers. Les formes droites étaient encore courantes, mais les formes courbes et enroulées étaient plus nombreuses. Leurs coquilles étaient plus richement ornées qu’auparavant, mais elles restaient simples en comparaison de celles de certaines de leurs successeurs. Les ouvertures des coquilles des espèces ordoviciennes étaient généralement circulaires [ p. 551 ] ou ovales, mais chez les espèces siluriennes, beaucoup d’entre elles étaient curieusement rétrécies (b,Fig. 402), en particulier chez les petites espèces courbes et enroulées. La constriction semble avoir été un dispositif de protection.
Bien que les gastéropodes aient été assez bien représentés au Cambrien et largement à l’Ordovicien, leur nombre n’a pas beaucoup augmenté au Silurien. Ils présentent une progression par la prépondérance des flèches élevées, une plus grande variété de formes et, pour certains, une taille plus importante ; mais les types plus anciens étaient encore nombreux.
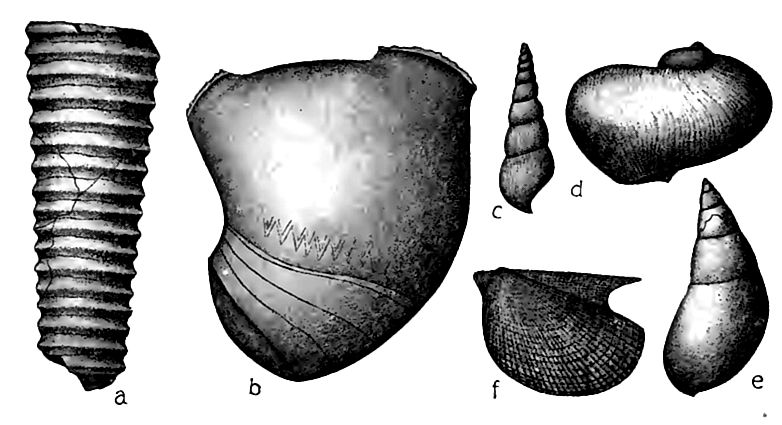
Les pélécypodes (f, Fig. 402) n’étaient pas aussi bien représentés dans les couches du Silurien moyen que dans l’Ordovicien, peut-être parce que les fonds calcaires leur étaient moins favorables.
Coraux. L’importance acquise par les coraux dans des situations appropriées est l’une des caractéristiques notables de la faune silurienne. À l’Ordovicien, les formes simples prédominaient sur les formes composées. Le rapport était maintenant inversé. Parmi les types notables figuraient le corail chaîne unique (Halysites, Fig. 403, c), apparu à l’Ordovicien ; le corail nid d’abeille (Favosites, a) ; [ p. 552 ] le corail tuyau d’orgue (Syringopora, b) ; et le corail coupe (Zaphrentis, e). Un corail très particulier de la classe simple (Goniophyllum, d) était quadrangulaire et son sommet pourvu d’un couvercle (opercule) de quatre plaques triangulaires articulées sur les quatre côtés du bord de la coupe.

Fermées, elles formaient une pyramide au-dessus de la coupe (d, Fig. 403, seules deux des plaques operculaires sont représentées). Il s’agissait d’un dispositif de protection inconnu parmi les coraux modernes.
Avec leur abondance croissante, les coraux ont pris l’habitude de s’associer. Ceci a donné naissance à des récifs. Les récifs connus semblent s’être formés à une certaine distance du rivage et être de type barrière. Cette tendance à la formation de récifs semble avoir été locale plutôt que générale, car sur de vastes étendues, les coraux sont dispersés de manière très dispersée.
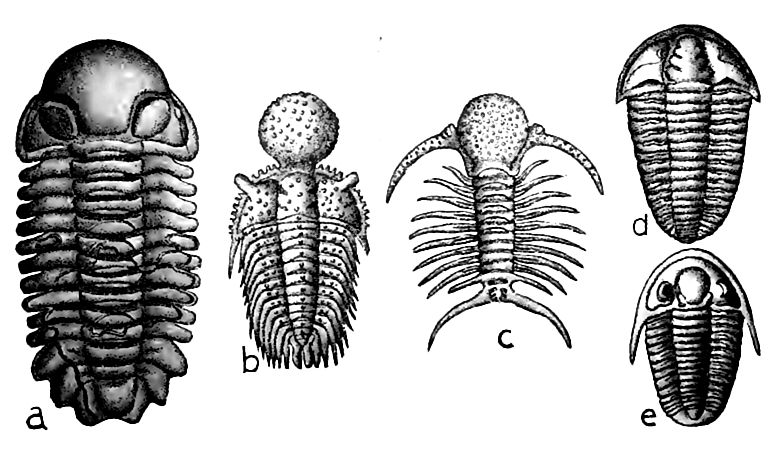
Les trilobites. Aucune nouvelle famille de trilobites n’apparut, bien que quelques nouveaux genres et de nombreuses espèces aient été ajoutés ; cela ne compensa pas la disparition des anciens, et la classe, bien que toujours importante, était déjà entrée en déclin numérique. Les formes les plus élevées étaient cependant structurellement égales, et peut-être supérieures, à toutes celles qui les précédaient.
Autres invertébrés marins. Les éponges prospéraient. Il y avait un champ prolifique d’entre elles dans l’ouest du Tennessee, où les conditions étaient non seulement propices à leur croissance, mais aussi favorables à leur préservation. La famille particulière des Receptaculites (Fig. 405), dont les affinités ont longtemps été mises en doute, était toujours présente, bien que son apogée fût passée. Les graptolites avaient perdu l’importance qu’ils avaient à l’Ordovicien et, à la fin de cette période, étaient proches de l’extinction.
[ p. 554 ]
Les vers marins se reconnaissent à leurs mâchoires, à leurs traces et à leurs terriers, ainsi qu’aux tubes calcaires que certains d’entre eux sécrètent.
Les vertébrés. Dans les dépôts du Silurien ancien et moyen, peu de vestiges de poissons ont été découverts, et ces quelques-uns sont très imparfaits ; mais dans la partie supérieure du système, leurs restes ne sont pas rares.
Plantes marines. La connaissance de la flore marine reste, comme auparavant, insatisfaisante. Bien qu’elle ait dû être abondante en théorie, seules d’obscures traces de type fucoïdal et quelques traces de formes supérieures ont été découvertes, et leur interprétation est plus ou moins douteuse.

Faunes étrangères et migrations
Laissant de côté l’étude de la faune particulière de la fermeture, on peut noter les relations de cet assemblage riche et varié du Silurien moyen. L’évolution générale de la vie sur les autres continents, pour autant que l’on sache, était similaire à celle de l’Amérique, mais peut-être moins prononcée et symétrique. Lorsque les formes sur différents continents étaient simplement similaires, la question de savoir si cette similitude était due à une intermigration ou à une évolution indépendante suivant des lignes similaires reste ouverte ; mais lorsque les formes sur des continents distincts sont identiques, surtout si les espèces sont particulières et aberrantes, on peut supposer qu’elles ont une origine commune et que la migration est indiquée. Un cas frappant de ce genre est celui du corail operculé quadrangulaire déjà mentionné (Fig. 403, d), que l’on trouve avec des idiosyncrasies identiques sur l’île de Gotland, dans la mer Baltique, et dans l’Iowa. Les preuves de migration sont renforcées par la présence à Gotland d’au moins trois genres particuliers de crinoïdes, également présents dans le haut bassin du Mississippi. Outre ces cas particuliers, de nombreuses espèces importantes et familières étaient communes à l’Amérique et à l’Europe, et certaines d’entre elles se trouvent également en Asie, en Australie et en Nouvelle-Zélande. On peut donc supposer l’existence de liens migratoires entre l’Amérique du Nord et ces terres. Étant donné qu’une trentaine d’espèces, voire plus, sont communes à l’Amérique du Nord et à l’Europe (intérieur des terres, Suède et Grande-Bretagne), et que celles-ci englobent un large éventail de genres aux mœurs très différentes, il existe une forte présomption de migration entre l’Amérique du Nord et le nord-ouest de l’Europe.
Il a déjà été dit que la mer intérieure de l’Amérique du Nord ne semble pas avoir eu de communication libre avec la mer à l’est ou au sud, et que son extension loin vers l’ouest est douteuse. Au nord, en revanche, la série de vestiges de formations siluriennes, probablement autrefois reliées, indique une large voie de communication pour la vie en eaux peu profondes entre l’Amérique du Nord et l’Europe, via le Groenland.
Les faunes qui occupaient la fosse des Appalaches, la baie du Saint-Laurent et la côte atlantique semblent n’avoir eu que des liens limités avec la faune de l’intérieur au Silurien moyen. Dans l’état actuel des connaissances, les faunes de la fosse des Appalaches présentent un aspect provincial.
La phase restrictive de clôture
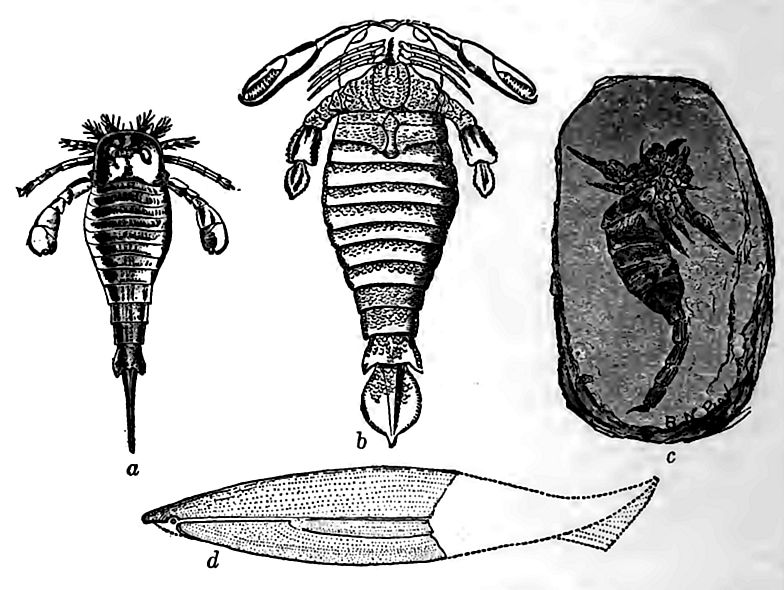
Après la vie luxuriante du Silurien moyen, il y eut, du moins en Amérique du Nord, un déclin notable, dû au retrait des eaux épicontinentales de la plus grande partie de l’intérieur, et à la transformation du reste en une mer excessivement salée, dans les dépôts de laquelle on trouve peu de fossiles. Les couches de Waterlime représentent un retour progressif dans le bassin de Salina à des conditions propices à la vie. La faune de ces couches est limitée, mais radicalement différente de celle de l’époque du Niagara. La plupart des types marins familiers sont absents de la faune plus récente, et sa caractéristique principale était l’abondance de grands crustacés de types à peine représentés auparavant. Les plus caractéristiques d’entre eux étaient le grand Eurypterus (Fig. 406, a) et le Pterygotus encore plus gigantesque (b). Les premiers atteignaient une longueur de 45 cm ou plus, et les seconds plus de 1,80 m à la période suivante. Ces dimensions étaient gigantesques pour des crustacés et n’ont probablement jamais été surpassées. Ces géants de leur espèce semblent clairement avoir été aquatiques, mais il est difficile de savoir s’ils habitaient les eaux salées ou douces. Ils sont totalement éteints, et leur habitat ne peut être déduit que de leurs associations. En Angleterre, en Suède et en Russie, les euryptérides sont associés à des fossiles marins, mais aussi à des graines de plantes terrestres et à des poissons qui, au stade suivant, semblent avoir principalement occupé les eaux terrestres. Au Dévonien et au Carbonifère, les euryptérides sont associés à des plantes terrestres, des scorpions, des insectes, des poissons et des amphibiens d’eau douce, ce qui semble impliquer un habitat d’eau douce. À la lumière de ces faits, l’hypothèse la plus courante est qu’il s’agissait à l’origine de formes marines, adaptées ultérieurement aux conditions d’eau saumâtre et d’eau douce. Une autre conclusion est qu’ils étaient à l’origine des habitants des eaux terrestres, et que leurs restes ont parfois été transportés vers la mer par les cours d’eau, et ainsi fossilisés avec des formes marines. Les mollusques, les crinoïdes, les coraux et autres formes marines similaires sont presque entièrement absents de la faune de Waterlime. Les rares brachiopodes trouvés sont généralement paupéritiques, comme s’ils vivaient dans des conditions défavorables. La présence occasionnelle de quelques formes marines incontestables n’indique pas tant que les eaux étaient habituellement salées, mais qu’elles l’étaient occasionnellement et partiellement.
[ p. 557 ]
C’est également à cette époque que les premiers scorpions connus sont apparus en Amérique et en Europe. Ils étaient apparentés aux euryptérides. Les formes européennes ont été considérées comme des espèces terrestres, bien que cette hypothèse ait été remise en question. Le dard et les glandes à venin ont été identifiés, et le nom significatif de Palceophonus, « ancien meurtrier », leur a été attribué en conséquence (Fig. 406, c). Les espèces américaines ont été considérées comme aquatiques.
La présence de poissons souligne les particularités de cette faune. À l’exception de leur présence en quelques points des Rocheuses à l’Ordovicien, aucun reste de poisson n’a été découvert en Amérique avant cette époque. En Europe, quelques poissons apparaissent un peu plus tôt, mais presque tous les restes de poissons de cette période encore découverts se trouvent dans les horizons les plus élevés du Silurien, ou dans les dépôts qui forment la transition vers le Dévonien, où ils sont associés à des euryptérides et à des plantes terrestres, ainsi qu’à des invertébrés marins. Il semblerait que les poissons de l’époque étaient variés et qu’ils aient été les précurseurs [ p. 558 ] des poissons abondants du vieux grès rouge du Dévonien. Comme le sable de cette formation a probablement été déposé par les eaux continentales, l’association des poissons avec les euryptérides et les plantes terrestres laisse supposer que cette faune particulière de crustacés vivait normalement dans les eaux continentales. Il ne faut cependant pas oublier que, dans les couches de transition, les poissons sont également associés à des fossiles marins, et il ne fait aucun doute qu’avant la fin du Dévonien, certains poissons au moins étaient véritablement marins.
Travail cartographique. Voir note à la fin du chapitre XVII, page 535.
| CHAPITRE XVII LA PÉRIODE ORDOVICIENNE (SILURIEN INFÉRIEUR) | Página de portada | CHAPITRE XIX LA PÉRIODE DÉVONIENNE |
¶ Remarques
[^4] : Sarle, Am. Géol., Vol. XXVIII, p. 282.
[^5] : Hartnagle, Taureau. 107, État de New York Mus.
Dans ce cas, il y a une certaine maladresse dans l’utilisation des termes Silurien inférieur, Silurien moyen et Silurien supérieur, pour les subdivisions du système, puisque le Silurien inférieur a longtemps été utilisé comme synonyme de l’Ordovicien, et le Silurien supérieur du Silurien, tel que ce terme est employé ici. ↩︎
Certaines des formations autrefois classées comme Oneida dans l’État de New York et du New Jersey sont désormais considérées comme datant de Salina (voir p. 543). ↩︎
Peut-être les couches de Richmond, les schistes de Maquoketa, etc. Voir les notes de bas de page, pp. 509 et 510. ↩︎
Williams, HS. Jour, of Geol., Vol. II, pp. 16-18, et folios Penobscot et Rockland, US Geol. Surv. ↩︎