Public domain
[p. 772]
¶ Formations and Physical History
The remaining periods of geological history constitute tl Cenozoic era, or the era of modern life. The era is variously sul divided, as shown below:
The threefold subdivision of the Tertiary (Eocene, Miocem Pliocene) is the one which seems to fit the phenomena of our continent best, but there is a growing tendency toward the recognition of the Oligocene. This means that beds are found in our continent with fossils similar to those of the Oligocene of Europe, rather than that the Oligocene of this continent constitutes a natural and major subdivision of the Tertiary.
Eocene formations are found in widely separated parts of North America (Fig. 524), though they do not appear at the surface over large areas. They include beds laid down in the seas and bays, and on the land. The former include- formations of marine ami brackish-water origin, and the latter those of lacustrine and su1>aerial (fluvial, pluvial, eolian) origin. The subaerial formations are probably more important than has commonly been recognizees The marine and brackish-water beds are confined to the borders of the continent, while the terrestrial deposits are found in many places in the Great Plains and farther west. Many of the formation! are not indurated, but locally they are even metamorphosed.
[p. 773]
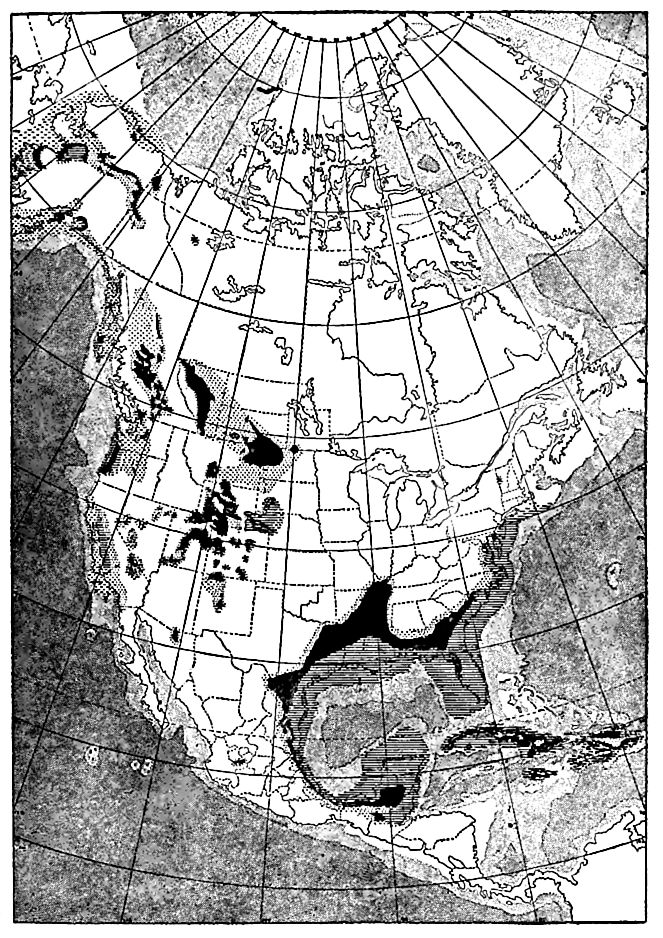
[p. 774]
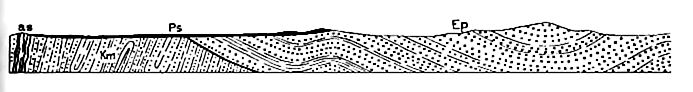
The eastern coast. Eocene formations appear at the surface in an interrupted belt from New Jersey to Texas. Their structure is similar (Fig. 499) to that of the Cretaceous, from which they are separated by an unconformity. The materials of the Eocene were derived largely from the Cretaceous, and subordinately from older formations farther inland. Clays, sands, and greensand marls are the most common materials of the system, and the conditions of sedimentation were much as in the preceding period.
The system is more fully represented in the Gulf region than en the Atlantic coast, and is thicker (1,700 feet maximum). It contains much lignitic matter in places, showing that marine conditions were not uninterrupted. In Texas,[1] gypsiferous and saliferous sediments recur at various horizons, though most of the beds are of marine origin, and there are numerous local unconformities in the system, suggesting recurrent changes in the conditions and areas of sedimentation. The principal subdivisions[2] are (1) Midway an; (2) Chickasawan; (3) Claibornian, and (4) Jacksonian.
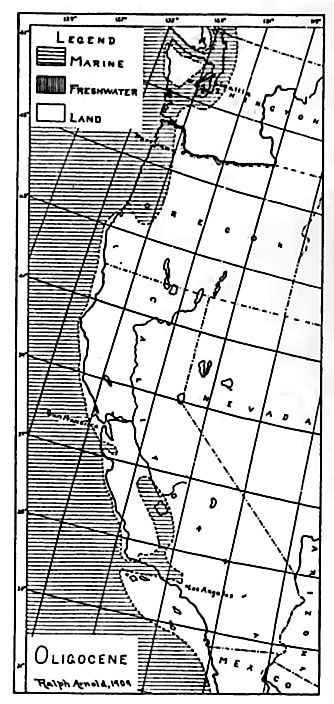
The Pacific coast. Marine and brackish-water beds.[3] Marine; Eocene formations are wide-spread west of the Sierra and Cascade ranges of California, Oregon, and Washington (Fig. 525), and have considerable development in Alaska. Throughout Washington and Oregon and in some parts of California, the Eocene is unconformal »le on the Cretaceous (in most places on the Lower Cretaceous = Comanchean or Shastan), while in much of California it is conformable on the Chico, the division plane between the two being indicated by a paleontological hiatus. These relations sugn that just before the Eocene, all of Washington, most of Oregon, and parts of the coastal region of California were land, over which the sea advanced later. The Coastal lands of the time were mostly low, but in southern Oregon and northern California there seem to have been mountainous areas, as also at some points farther south.
[p. 775]
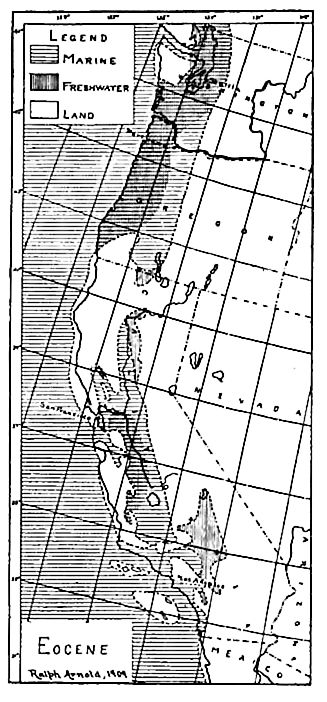
The rocks of the Eocene system are mostly clastic, sandstone and shale predominating, but there are conglomerates, tuffs, and liatomaceous shales, the last thought to be a source of oil. In not a few places, marine beds are succeeded by brackish-water deposits.
By the beginning of the Eocene, the Puget Sound depression, perhaps to be correlated with the great valley of California id the Gulf of California, had igun to show itself.[4] The Hympic and Cascade mountain regions on either side were high lands, but not mountainous; and the region of the sound was a reat estuary, in and about which leposition was in progress. Some of the sediments accumulated in brackish water and on land, and suited in the thick coal-bearing Puget series of Washington, the upper part of which is Oligocene or even Miocene.[5] The series is said to contain 125 beds of coal thick enough to attract prospectors. Most of the workable coal is in the lower part of the series. The area of deposition extended south into Oregon, and east well toward the Blue Mountains of that state.
British Columbia appears to have been land during the period,[6] but Eocene beds, much disturbed, [p. 776] have been recognized in Alaska (Kenai scries) whore they arc coalbearing in places.
The Eocene system has an estimated thickness of 10,000 to 12,000 feet in southern Oregon, and but little less in southern California.
After the Eocene, there was a time of temporary elevation, erosion, and volcanic activity along the Pacific coast, with considerable basaltic flows in Washington and Oregon.
The western interior. The warpings, faultings, and the intrusions and extrusions of lava which marked the close of the Mesozoic era in the west appear to have developed lands which were relatively high, adjacent to tracts which were relatively low. The steep slopes of the mountain folds, fault scarps, and volcanic piles seem to have afforded the conditions for rapid erosion, while the adjacent lowlands furnished places of lodgment for much of the sediment. Some of it took the form of fans and alluvial plains, and some of it probably lodged in lake basins formed by warping, or by the obstruction of valleys by lava flows. The wind also made its contribution to the deposits of the time. The result was an inextricable combination of fluvial, pluvial, eolian, and lacustrine deposits. Deposition on land was therefore a feature of the period. as of all subsequent time, and among the accessible formations of this and later periods, those of terrestrial origin are more widespread than those of marine origin.
The sites of principal sedimentation shifted somewhat from time to time, and among the widely distributed deposits referred to this period, there are great differences of age. Several more or less distinct stages of deposition have been made out, the distinctions being based partly on the superposition of the 1km Is, and partly on their fossils.[7] These several stages are not readily correlated with those of the coasts.
1. Reference has already been made to certain formations which have been classed commonly as Cretaceous, which should probably be regarded as early Eocene (p. 752).
TABLE OF TENTATIVE CORRELATIONS OF THE TERTIARY FORMATIONS OF CALIFORNIA, OREGON AND WASHINGTON. RALPH ARNOLD, January, 1909
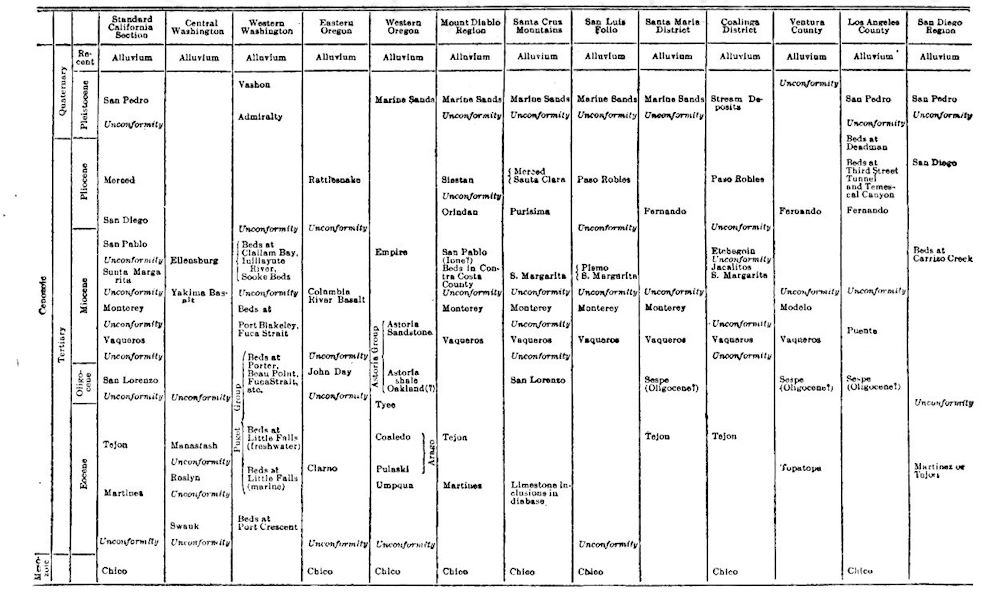
[p. 777]
Some of these beds are inseparable from the Ft. Union formation (or series), heretofore regarded as the oldest division of the Tertiary in the western interior. During the Ft. Union stage, there was an extensive area of aggradation in parts of North Dakota[8] and Montana, and a still larger area in Canada. The Fort Union beds, composed of sand, clay, etc., are said to be locally 2,000 feet or more thick, and have usually been described as lacustrine. The presence of freshwater shells (unios, etc.), is consistent with this conclusion for some parts of the formation; but the abundance of the leaves at many places is indicative of subaerial aggradation for other parts.[9] Eocene formations of similar age are found in Colorado (Telluride [10] and Poison Canyon[11] formations) , New Mexico (part of the Puerco beds) , and elsewhere.
The sites of early Eocene deposition were finally shifted. In so far as the sedimentation was in lakes, the basins may have been filled or warped out of existence, and in so far as it was subaerial, the deformative movements of the time, or the progress of the gradational work of the streams, or both, may have been respon Isible for the shifting.
2. During the next or Wasatch stage of the period, sediment was being deposited over parts of Utah, western Colorado, and Wyoming, and in some other places. The beds of this stage have a maximum thickness of several thousand feet, and are now 6,000 to 7,000 feet above the sea. About 77% of the fossils of this stage are of terrestrial life.[12]
3. The third recognized stage of the Eocene of the west is the Bridger,[13] during which sedimentation was in progress in various places. One area was in the Wind River basin, north of the mountains [p. 778] of that name, and another, a little later, in the basin of the Green River, both in Wyoming, and in Utah south of the Uinta Mountains. It may have been during this stage, too, that the volcanic tuff (San Juan formation, 2,000 feet and less thick) of the Telluride region was made.[14] This last formation is of interest as an index to the vigor of volcanic action in this region.
4. The Uinta stage[15] followed the Bridger. Deposition was then in progress in southeastern Utah and southwestern Colorado. The deposits occupied a part of the area covered by the Wasatch and Bridger formations, so that these formations are found in superposition, in some places. The Uinta beds now have an altitude of as much as 10,000 feet, though they were probably deposited at a much lower level.[16]
The northwest. In the northwest there are Eocene formations not definitely correlated with the preceding stages. Thus in northern Oregon, there are late Eocene beds of terrestrial origin (Clarno formation, largely volcanic tuff[17]) in the John Day basin, which was the site of aggradation during a large part of the Tertiary. In Washington two thick sedimentary formations (the Swank, early Eocene, 3,500-5,000 feet, below, and the Roslyn, 3,500 feet) of Eocene age and non-marine origin, are separated by 300-4,000 feet of basalt. The Payette formation of Idaho, said to have been accumulated in a lake formed by the damming of the upper basin of the Snake River by the early lava-flows of the Columbia River region,[18] is now referred to the Eocene.[19] Eocene beds of terrestrial or volcanic origin are imperfectly known in many other places wesl of the Rocky Mountains. The erosion of the Eocene has given rise locally to the topography characteristic of “Bad Lands.”
General considerations. It has been customary to regard the Eocene and later periods as much shorter than those of the Paleozoic [p. 779] and Mesozoic; but this conclusion may be questioned. On the basis of thickness, the showing of the system is great, as the formations of Puget Sound, Coos Bay, Ore., and southern California show. In the western interior, too, the thickness of the system is great, if the thicknesses of the beds deposited in the several successive areas of deposition, are added. Furthermore, any just estimate of the duration of the period must take account of the great erosion after the post-Laramie deformation before the deposition of the recognized Eocene began. On the physical side, therefore, there is no warrant for the conclusion that the period was short. The faunal developments of the period, too, were such as to make great demands upon time. On the whole, it does not seem improbable that the period was as long as the average of those since the beginning of the Paleozoic.
The conditions requisite for so great thicknesses of terrestrial sediment as occur in the Eocene of western North America are not easily conceived, if the thicknesses are really as great as they have been thought to be. If the areas concerned were in process of more or less continuous warping, the depressions going down as surrounding lands went up, or if troughs or basins of deposition were produced by faulting, the bottoms sinking while their surroundings rose, the conditions for thick sediments would perhaps be met. It has sometimes been urged that these and similar formations are [p. 780] too thick to be subaerial; but it is not apparent that it is more difficult to acount for thick subaerial sediments, under the conditions indicated, than to account for thick lacustrine or marine formations.
The relations of the Eocene beds of the western interior an such as to indicate that both the attitude and altitude of the surfaces in that part of the continent were very different from those which now exist. That region must have been much lower than now, and, locally and temporarily at least, without well-established drainage. The present mountains were certainly not so high as now, though considerable elevations and great relief doubtless existed.
Close of the Eocene in North America. The closing stages of the Eocene were marked by crustal movements in the west, resulting in considerable changes in geography. Such movements had been in progress throughout the period, as has been indicated; but the changes at the close were on a larger scale. The deformative movements seem to have included both faulting and folding. The result was the retreat of the sea along the Pacific coast, the development of new areas of high and low lands, and therefore a shifting of the sites of rapid degradation and aggradation. Outside the Cordilleran region the changes were less.
Along the Atlantic and Gulf coasts the Miocene is in many places unconformable on the Eocene, and it was at the close of the Eocene (or perhaps during the Oligocene) that an island, now included in the peninsula of Florida, was formed. In the Carolinas, and in the western Gulf region, the conformity between the Eocene formations and those classed as Oligocene seems to preclude notable changes of geography along the coast in the southeastern part of the United States at the close of the period.
Foreign
Europe. Considerable lakes, estuaries, and perhaps other areas of deposition remained over western Europe within the ai from which the sea withdrew, at the close of the Mesozoic i Later, but still early in the Eocene, submergence of the land set in, allowing the sea to again overspread considerable areas from which it had been temporarily excluded. In western and central Europe the maximum submergence of the Eocene seems to have been accomplished by the middle of the period. Toward its close, the epicontinental waters of the northwestern part of the continent were again restricted. These changes are inferred from the considerable development of fresh- and brackish-water beds in the lower part of the system, of marine beds above them, and of non-marine formations again at the top. Most of the system in central and western Europe is clastic, and the beds are still unindurated, for the most part, and not very thick.
[p. 781]
TABLE OF TENTATIVE CORRELATIONS OF THE TERTIARY FORMATIONS OF CALIFORNIA, OREGON AND WASHINGTON. RALPH ARNOLD, January, 1909

[p. 782]

In the south, the Eocene sea spread much beyond the borders of the present Mediterranean, covering much of southern Europe, northern Africa, and part of southeastern Asia. The eastward extension of this sea joined the Indian Ocean, cutting off the southern peninsulas of Asia from the Mainland to the north. A narrow sound east of the Urals probably connected the Arctic Ocean with the expanded Eocene Mediterranean. Out of this extended sea rose many islands, some of which corresponded in position to the Alps, Carpathians, Apennines, and Pyrenees.
On the bottom of this great body of water, which should perhaps be thought of as a part of the i rather than as a mediterranean sea, limestone was deposited on an oxtensive scale. Much of it is made up almost wholly of the shells of nummulites (foraminifera, Fig. 528) and is found from one side of the Old World to the other. Since it is often thick (locally several thousand feet), as as wide-spread, the sea must have swarmed with foraminifera, and the period must have been long. Hardly anywhere else in the rocks of the whole earth are there indications of such great numbers of organisms of one kind. The Fusulina limestone of the Carboniferous is perhaps most nearly comparable. Some idea of the deformative movements since the Eocene may be gained from [p. 783] the fact that the nummulitic limestone occurs at elevations of more than 10,000 feet in the Alps, up to 16,000 feet in the Himalayas, and 20,000 feet in Tibet. The principal elevation of the Pyrenees, Carpathian, Caucasus, Thian Shan, and other high mountains of Eurasia, is post-Eocene. In the Old World, therefore, as well as in the New, the greater relief features of the present were undeveloped in the Eocene period.
Other continents. In Africa, marine Eocene is known along the northern and western coasts, and in the Soudan. It is known also in South Australia, New Zealand, and Tasmania, though not generally sharply differentiated from the later Tertiary. At the head of the great Australian Bight, there is a thick bed of Eocene chalk. Eocene beds are also known in various islands of the Pacific.
The Tertiary formations of South America have not been closely correlated with those of other regions. There is marine Eocene along some parts of the western coast, in Patagonia[20] (Magellanian series) , where the beds are usually unconformable on the Cretaceous, probably in Argentina, and along at least a part of the coast of Brazil.[21] Eocene beds of non-marine origin also occur in Patagonia.[22]
Eocene beds, not always distinctly separable from the Oligocene, are extensively developed in the West Indies, where limestone is the dominant type of rock. In the Caribbean region they occur up to elevations of 5,000 feet on the mainland, and 10,500 feet in Hayti.[23] It was formerly thought that the Atlantic and Pacific oceans connected freely across Panama during the early Tertiary, but the work of Hill renders it doubtful whether there were more than shallow and restricted connections in the Eocene, and whether there was connection of any sort later.
General geography of the Eocene. Eocene geography was very different from that of the present time, and the differences were [p. 784] perhaps even greater than has been indicated. It has been conjectured that North America was connected with Asia on the west . via Alaska, and with Europe on the east, via Greenland and Iceland. Land seems to have failed of making a circuit in the high latitudes of the north only by the strait or sound east of the Urals. In the southern hemisphere, it has been surmised that Antarctica was greatly extended, connecting with South America, Australia, and possibly with Africa, and that Africa and South America were connected across the Pacific[24] from some earlier time until after the beginning of the Eocene. The basis for these conjectures Is found in the distribution of life at that time, as shown by fossils.
If these conjectured extensions of land were real, it will be seen that the division of land and water in the northern and southern hemispheres was far less unequal than now, and that the land was massed in high latitudes to a greater extent than at present, while tropical seas were more extensive. If extensive polar lands were the cause of glacial periods, as has been suggested, the geographic conditions of the Eocene were favorable in the extreme, if the relations sketched above were the real ones. In spite of this, the climate of the period seems to have been genial, and less markedly zonal than now.
Close of the Eocene. During the later part of this period, and at its close, there were some notable deformations in Southern Europe. The initiation of the Pyrenees, and of some of the mountains farther east, is assigned to this time, and the distribution of the later formations, when compared with the distribution of the Eocene, shows that changes of a less pronounced type were in progress. The greater deformations which express themselves in the mountains of Southern Europe are post-Eocene, and most of them considerably later than the close of the Eocene.
¶ The Eocene Life
The Transition from the Mesozoic
Four salient features marked the transition of life from the Mesozoic to the Cenozoic: (1) Among marine animals, nearly all [p. 785] Cretaceous species were replaced by new ones; (2) among land plants so many species lived over as to make the plane of division between the Mesozoic and Cenozoic very difficult of location; (3) the great saurians almost disappeared, and most other reptiles showed profound changes; and (4) placental mammals appeared in force, and promptly took a leading place. The combination is unique, in that, while half the land life joined with the sea life in undergoing a profound transformation, the other half did not participate notably in the revolution. In explanation of profound transformations of epicontinental marine life, appeal has been made repeatedly to the withdrawal of the sea, to the extension of the land, and to climatic changes incident to deformative movements; and this appeal may be renewed so far as the change in the sea life is concerned. The withdrawal of the sea after the Cretaceous period seems peculiarly well fitted to explain the great change in epicontinental sea life, because of the great restriction of the area of shallow water. The increase of the land and the establishment of new land connections at the same time might well have caused the existing vegetation to spread and nourish, if the climate remained congenial; but why did the land faunas not respond in like manner? Closely connected with this problem is the sudden rise of the mammals.
It is an open question whether the placental mammals of North America and Eurasia arose from non-placental mammals which lived in these continents earlier, or whether they were immigrants. No satisfactory evidence of a transition from non-placental mammals has been produced, and the strength and suddenness of the placental development suggest invasion from some quarter where their evolution had reached an advanced stage. The deformative movements which closed the Cretaceous period and inaugurated the Eocene quite certainly made many new land connections, and furnished the conditions for an invasion, if mammals in sufficient variety and numbers were awaiting the opportunity.
The rise of placentals may have caused the downfall of the reptiles, though this cannot be affirmed. The habit of bringing forth relatively mature offspring, and of nourishing and protecting them, gave the mammals an immense advantage. To this advantage were added superior agility and higher brain power. It [p. 786] is not surprising, therefore, that the rise of the mammals drove the clumsy, affectionless, small-brained reptiles either into extinction, or to the assumption of new and smaller forms.
The angiosperms may have been a factor in the placental dispersion, through the fact that they are a staple source of food of the mammals. It may have been the spread of this flora from its originating tract, until it came into contact with the primitive placentals in their originating tract, that caused the rapid spread and evolution of the latter.
The Vegetation
In plant history, the Eocene was not the dawn of the recent, for the great change from medieval to modern plants took place in the Comanchean period. The Eocene was not even the period of any radical innovation. There was, however, much progress toward living species, and toward present adaptations of plants to climate, soil, and topography, and to each other.
Among the plants of the earliest known Tertiary flora of Europe were oaks like those of the present elevated districts of warm temperate zones. With them were willows, chestnuts, laurels, ivies, aralias, etc., which have been likened to the flora of southern Japan.
The flora of the Denver beds (p. 752), contains figs, poplars, laurels, magnolias, and many ferns. The early Eocene flora of southern Canada[25] included similar forms, together with oaks, beeches, etc., a flora indicating a temperate climate.
The Middle Eocene flora of England records a flora “the most tropical in general aspect which has yet been studied in the northern hemisphere”[26] while a later flora “suggests a comparison of its climate and forests with those of the Malay Archipelago and t ropical America.” The Mid-Eocene series of America in temperate latitudes contains palms and bananas, mingled with many other trees of similar climatic significance.
[p. 787]
The Land Animals[27]
The undifferentiated nature of the early Eocene mammals. It is difficult to carry our conceptions of mammals back to the Eocene prototypes, without carrying with them distinctions which did not then exist. The earliest Eocene mammals were much more primitive than those of the Middle Eocene, and their rapid convergence backward seems to point to some set of conditions which caused a rapid advance of the class at this time whatever their previous history. The coming into a new domain of rich and varied conditions whether by immigration or indigenous development, may be safely included among these conditions.
The mammals of the earliest Eocene, as now understood, included several vaguely differentiated groups, in which existing orders were foreshadowed rather than represented. The herbivores were foreshadowed by the Condylarthra, and the carnivores by the Creodonta; but the two were not sharply differentiated. Both were five-toed plantigrades, whose phalanges had horny coverings that were neither hoofs nor claws. Ancestral edentates, insectivores, rodents, and lemuroids seem to have been represented in an obscure fashion.
The evolution of the mammals was so rapid that before the close of the Eocene, the Herbivora (Ungulata), Carnivora, Edentata, Insectivora, Rodentia, Quadrumana, Cetacea, and Sirenia, and probably the Cheiroptera were distinctly defined (see p. 945). None of the present genera, however, are known as early as the Eocene. When it is recalled that the name Eocene was founded on the presence of some few species of living invertebrates, the great difference between the stage of their evolution and that of the mammals may be realized. In general the mammalian faunas of the early Eocene were closely similar to those of Western Europe, while in the middle and late Eocene there seems to have been faunal separation from Europe.[28]
The main herbivore line. While the condylarths and creodonts were- near each other at the beginning of the period, the hoofed [p. 788] herbivores and the clawed carnivores soon became distinct. The condylarths (Fig. 529) were small generalized forms with five toes and forty-four teeth, not yet developed into the true herbivorous type. They have lived on till now without radical change, but one branch, adapted to forests and marshes, seems to have diverged early, and to have given rise to the ungulates. In the course of the period many of them became fitted for life on grassy plains.
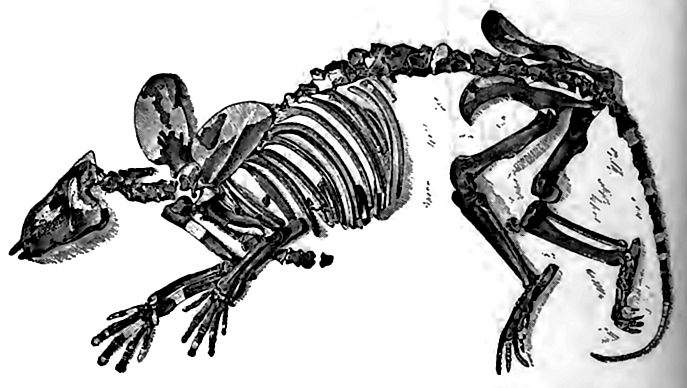
To this end, there was a progressive abandonment of the flat, heavy, palmate form of foot, and the acquisition of the light, springy, digitate habit, adapted to a quick start and swift flight. At the same time hard hoofs, and powerful grinding teeth were develops 1. The evolution of hoofs and grinding teeth has been thought to be connected with the prevalence of grassy plains, the firm turf of which is in contrast with the soft soil of forest and marsh. Gras also are much associated with dry and even semi-arid grounds, and dessication intensifies the firmness of the bottom, and gives additional occasion for the hoof. The forests perhaps helped to preserve a section of the evolving order in its more primitive form.
Back of these influences lay the physical conditions that promoted them. In western America, where the evolution is best [p. 789] known, the lakes and rivers were undergoing changes. If these followed the methods of to-day, they left behind them, as they shrank or shifted, borders of grassy or sedgy ground which, on fuller drainage, often became prairie. Such changes were suited to the evolution of herbivorous prairie life, and this in turn must have invited its appropriate contingent of predaceous animals. If these considerations are valid, the prime factors in the evolution of the ungulates were (1) an undifferentiated plastic animal group susceptible of modification; (2) a plant group (grasses and fodderfurnishing angiosperms) affording appropriate food for the new type; and (3) the shrinkage and shifting of lakes, marshes, and lodgment plains, and the drying up of the plains of the continent, resulting in prairies whose hard turf favored the development of foot and limb modification in the interest of speed.
The era of bulk and heavy armor, such as had been possessed by the reptiles, had passed, and an era of agility and dexterity had begun. No small factor in this progress was the increase in intelligence indicated by the larger brains. The lighter and more agile frame was accompanied by the development of smaller, simpler, but more effective weapons of attack and defense. Nevertheless size continued to be important, and some species in almost every sub-order reached and passed the limit of bulk-advantage, and then declined.
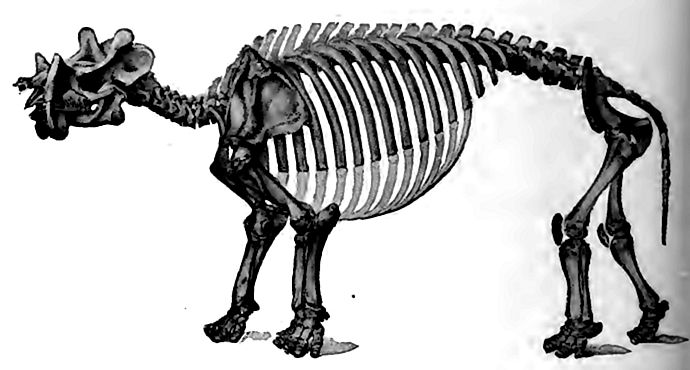
In the course of the early evolution strange forms appeared, and soon became extinct. Among them were the Dinocerata (Fig. 530) , grotesque monsters whose skulls were armed with three pairs of protuberances, perhaps horn cores, and a pair of enormous canine teeth or tusks projecting below (at least in the male), and an extravagant attempt at armature on both upper and nether sides. Their brains were smooth and singularly small for such ponderous bodies. In them, brute mass and low brain-power seem to have reached their climax among mammals.
The divergence of ungulates into odd- and even-toed. Early in the Eocene, the hoofed animals began to diverge into oddtoed (perissodactyls) and even-toed (artiodactyls) types. In the former, the main line of support is in the axis of the middle toe; in the latter, between the third and fourth toes. In the course of time [p. 790] the lateral toes fell out of use and were atrophied. The first class! reached its extreme type at length in the horse, and the second in’ our cloven-hoofed cattle; but these perfected types were not attained in the Eocene, for the evolution of the perissodactyls did not pass beyond three-toed forms in the Eocene.
The horse has become a classic example of evolution. The earliest recognized form was the Hyracotherium (Fig. 531), whose equine characters were obscure. Pachynolophus represented a slight advance, and the Orohippus (Epihippus) a greater one. The latter was four-toed (three functional) in front and three-toed behind, and the limbs and teeth were slightly modified in the direction of the horse. These forms were about the size of a small dog, and as much canine as equine in appearance. The evolution continued through the remaining periods of the Tertiary, true horses appearing in the Pliocene. The primitive Eocene forms lived both in Europe and America, and the evolution followed similar lines in the two continents.
The rhinoceros family appears in the record in the later part of this period, but had its development chiefly in the next.
Artiodactyls emerged from their generalized beginnings more slowly. Suina (pigs, peccaries, hippopotamuses) were represented [p. 791] in the Bridger epoch by a primitive small hog with strong canine teeth of somewhat carnivorous aspect. Strangely enough, Tylopoda (camels, llamas) seem to have their beginnings on the American continent in the middle and later Eocene, and to have flourished here until the Pliocene. Then, having previously sent a branch to South America to evolve into llamas and vicunas, and another into the Old World to become the present camels, the tribe died out in its primitive home.

The carnivore line. It has been thought by some paleontologists that the creodonts were more primitive than the condylarths, and that the latter diverged from the former, as also the edentates and the rodents. If this is so, it gives the creodonts the central position among the primitive mammals. The creodonts ranged throughout the whole period and passed into the next, gradually giving way meanwhile to their own more progressive descendants. They were common in America and in Europe, and they lived in South America. Modern types began to emerge definitely from the ancestral forms toward the end of the period. Patriofelis (“the father of cats,” a name not to be taken too literally) of the Bridger epoch, presented a suggestive combination of characters, some features resembling [p. 792] those of cats, and others those of seals. Some species seem to baa been aquatic. Primitive representatives of the dog family appeal! in Europe late in the period.
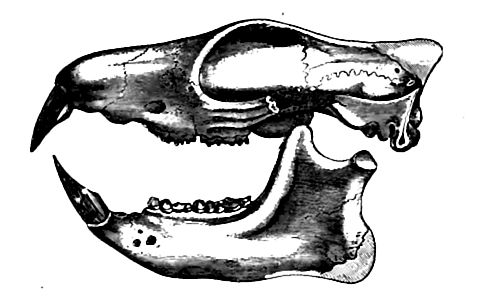
Edentates, rodents, and insectivores. The similarity of the ai cestral edentates to the condylarths and creodonts of the earlie Eocene seems to imply that the three orders had but recently diverged from common ancestors. The primitive rodents of the early Eocene had incisors which had just begun to assume their specific gnawing functions. By the middle of the period, rodents became a notable element in the fauna, the Tillotherium of the hri< Information having finely specialized incisors (Fig. 533). The primitive squirrel appeared in Europe in the latter part of the period. Even to-day, the rodents retain many primitive characters, and since the Miocene they have undergone few radical changes. derivation is not yet determined.

Most living families of insectivores can be traced back to the Eocene. They still retain many primitive characters, and arc the least altered of the great mammalian branches.
[p. 793]
The non-placental mammals. During the Eocene, early forms of the opossum appeared in both the Old and New World. They retained this wide distribution until the Miocene, when they disappeared in Europe, but they have persisted in North and South America to the present time. It is a singular fact that the monotremes, the lowest of the mammals, are not known to have appeared until after the Tertiary.
The primates (Quadrumana). No traces of apes have been found in the Eocene, but representatives of the lower primates, the lemuroids, appeared in the Wasatch epoch in America, and in a similar horizon in Europe. This is the more notable, as the lemurs are now confined to Madagascar, Africa, and southern Asia. The lemurs show many affinities with the insectivores, and were possibly derived from them. The apes are probably descended from the early lemuroids.
The mammals go down to sea. Just as the land reptiles of Mesozoic times took to the sea by choice or necessity, so did the mammals in their day. Thus arose cetaceans (whales, dolphins, porpoises), sirenians (manatees, dugongs), and pinnipeds (seals, sealions) . In parts of Alabama, vertebra? of primitive whales (Zeuglodons) were originally so abundant as to attract popular attention and call forth legends of divers catastrophes.
[p. 794]
Birds. Fossils of ancestral gulls, herons, flamingoes, albatn buzzards, falcons, eagles, owls, woodcock, quails, ploverostrich-like, flightless birds of great size, with not a few forms I doubtful interpretation, have been found, showing great deployment of this class.
Reptiles and amphibians. One of the greatest contrasts in geological history is found in comparing the size, power, and multin tude of the Cretaceous land reptiles with those of the Eocene. Oi the great saurians, only a few lived on into the Eocene and they did not live long. Land reptiles seem to have become ran in the period. There were turtles on both land and sea, and souk of them attained large size. There were crocodiles which belong^ about equally to land and water; also snakes, some of them Large. Amphibians were present, but apparently not abundant.
The insect life. There has been little important change in the insect world since the beginning of the Cenozoic era. Few new families have appeared, though the genera and species have changed.[29]
Marine Life
The name Eocene, founded upon the presence of a small percentage (less than 5%) of living species among the marine invertebrates, implies their stage of advancement. Not only were bite existing orders, families, and genera established, with some e tions, but even the present species had begun to appear. The changes that follow from this time on are valuable as criteria of correlation, climate, migration, and other elements of the later history, but they do not record profound biological transformations They stand in striking contrast with the radical and rapid evolution of the mammals.
Geologically, the most striking feature of the marine E< life was the extraordinary abundance and size of the foraminifers (Fig. 534). Reference has already been made to the prodigious abundance of the nummulites. Gastropods and pelecypods of modern types were very numerous, but cephalopods were much less important than in the Cretaceous. Sea-urchins continued to be [p. 795] abundant, the corals had taken on modern forms, and decapods were rising in importance.
The American Eocene faunas[30] were rather pronouncedly provincial, though some species have a rather wide range. So pronounced is the provincial character of the faunas, that much difficulty is experienced in making correlations between formations along different parts of the Atlantic and Gulf coasts, and greater difficulties arise in regions more widely separated. The variations are, however, variations of detail, not of broad features.

[p. 796]

[p. 797]
Climate. The marine fauna of the Pacific coast,[31] and the flora[32] as far north as Puget Sound, point to a subtropical climate.
¶ The Oligocene Formations
In North America. As already stated, formations correspond| ing to the Oligocene of Europe have not usually been differentiated, in North America. The differentiation may be justified on paleontological grounds, if it is desirable to make the classification for this country conform closely to that of Europe.[33] If the Oligocene is to be recognized, certain formations along the Atlantic and Gulf coasts, formerly regarded as late Eocene or early Miocene, should be classed as Oligocene. In the Gulf region the Vicksburg (below) and Grand Gulf formations[34] of Alabama, Mississippi, and Louisiana, and the Fayette[35] formation of Texas, belong to this category.
The early Oligocene is represented generously about the Caribbean sea, where its association with the Eocene is generally close,[36] and its separation from the Miocene distinct. This is in keeping with the phenomena of the Gulf States. Limestone is the dominant type of rock in the Antillean region.
The Oligocene stage is also recognized among the terrestrial deposits of the western part of the continent. The White River formation, now classed as Oligocene, occupies an extensive area in northeastern Colorado, southwestern Wyoming, western Nebraska (Brule and Chadron formations[37]) , and South Dakota, and perhaps [p. 798] in Kansas.[38] In the light of present knowledge, it seems probable that all phases of land aggradation, lucustrine, fluvial, and eoliajj are represented in this formation.[39] Even thin beds and len limestone and volcanic ash enter into it. The formation is said to have originally covered most of the Black Hills region, and possibly all of it.[40] Remnants are now found up to elevations of more than 6,000 feet, and the highest points of the hills are but little higheft The Florissant beds in South Park, Colo., consisting largely of volcanic ash, and famous for their extraordinary number of insects, are classed as Oligocene. So also are some of the beds of the John Day Basin of Oregon,[41] unconformable above the Eocene. Marine Oligocene beds are found in California, Oregon, Washington, British Columbia, and Alaska, at least,[42] but at many points on the Pacific coast, the record of the period is found chiefly in Che uncomformity between the Eocene and the Miocene. The lands which contributed the Oligocene sediments were mostly low. Shale is the dominant rock of the series, which is much thinner than the Eocene.

[p. 799]
Considerable geographic changes occurred during the Oligocene, or at its close, especially in the Gulf and Caribbean regions,[43] where the Oligocene (early Oligocene) is commonly conformable on the Eocene, and unconformable beneath the Miocene.

Foreign
Europe. The Oligocene is more distinctly differentiated from the Eocene in Europe than in most parts of America. Toward the close of the Eocene, the epicontinental sea of northern Europe was greatly restricted, but considerable areas stood so near sea-level that slight changes served to greatly diminish or extend the epicontinental waters.
[p. 800]
The oldest Oligocene deposits of central and western Europe are largely of terrestrial, fresh- and brackish-water origin. Local deposits of salt, gypsum, and coal are suggestive of the physical
conditions at various times and places. The Oligocene of southern Europe is chiefly marine, hut in the upper part of the series, lake and marsh deposits are not rare. In Italy it has been estimated to have the extraordinary thickness of nearly 12,000 feet.
In Europe, as in North America; there were considerable igneous eruption, during the Tertiary, and especially during the Oligocene. The results are to be seen in Bohemia, where there is much igneous rock; in northern Ireland and western Scotland, where outpourings of lava probably made great plateaus, of which some of the adjacent islands are remnants; in Iceland, and in the Vienna basin. Between eruptions, vegetation grew in the marshes and shallowlakes and over the surface of the lava. The substance of this v< tation is locally (Faroes, and land) preserved in the form of coal between the lava beds. Someoi lakes of France seem to have been obliterated by volcanic action.
Amber. One of the peculiar accessories in the Lower Oligocene is the amber of northern Germany, principally in the vicinity of Konigsberg. While amber in small quantities is found in Sicily and a few other places, that of the Baltic region is more abundant than that of any other part of the earth, so far as now known.
[p. 801]
Amber is fossilized resin, apparently from certain varieties of coniferous trees. Its original position in the Baltic region appears to be in certain glauconitic beds of a clayey nature, but parts of this formation have been worn by the waves, and the amber distributed. Some of that which finds its way into commerce is picked up on the Baltic shore, while some is taken from the beds in which it was originally entombed. One of the interesting features of the amber is the fact that it frequently contains insects. The insects seem to have alighted upon the resin while it was soft, and to have become completely immersed in it, and perfectly preserved. About 2,000 species have been found thus embedded.
Considerable deformative movements made themselves felt in Southern Europe at or about the close of the Oligocene, as in the Balkan and Carpathian Mountains.[44]
Other continents. In other continents, the Oligocene has not been generally differentiated, but it is known in northern Africa and in Patagonia,[45] where it is partly marine and partly non-marine.
¶ The Life of the Oligocene
The vegetation. The mixed evergreen and deciduous forests of the Oligocene were similar to those of the Eocene, especially in Europe, where palms continued to be abundant and varied, growing even in north Germany. They seem to have become rare, however, in the United States, for in the Florissant sediments, which are rich in plant fossils as well as insects, palms are barely represented. The Florissant fossils show a variety of angiosperms, widely distributed through the several orders that are now found in the latitude of the middle and southern states.
The land animals. All the species of insects of the Florissant beds[46] (over 700) are extinct. This seems to indicate that although the types had all become modern, the species continued to change with relative rapidity. Fish fossils are abundant in the same beds.
[p. 802]
Mammals continued their rapid evolution without interruption! and perhaps even with some acceleration. The Carnivora came into clear definition, and were represented in the White River beds by ancestral dogs, cats, coons, and weasels, while some creodonts remained. Rodents were represented by squirrels, beavers, pocket-gophers, rabbits, and mice. Among perissodactyls, the rapidly developing horse family was represented by Mesohippus and Anchippus. The rhinoceros tribe had deployed into three branches, one a lowland form, ancestral to the existing family, another aquatic, and a third an upland, horse-like, running form. The tribe had a cosmopolitan range.
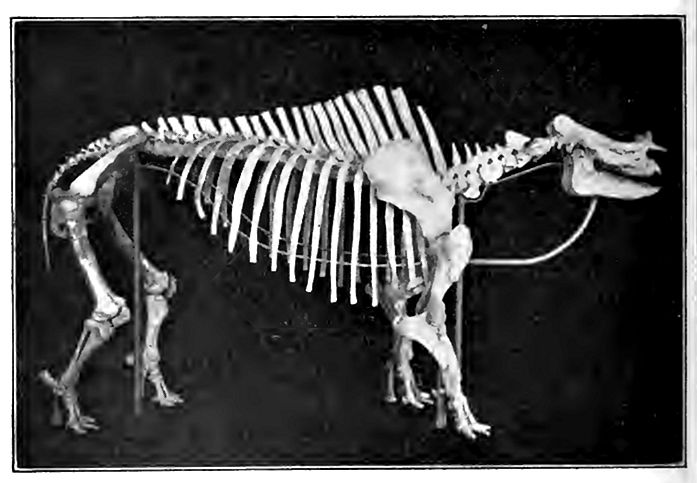
The titanotheres, an erratic branch of the odd-toed ungulates which arose late in the Eocene, reached their climax in the Oligcj cene (White River), and then disappeared. They were intermediate in proportions between the rhinoceros and the elephant, and were distinguished by a long, depressed skull armed with a pair of [p. 803] horns near the extremity of the nose, as were their kin the rhinoceroses, but placed transversely, as in the ox (Fig. 539). They reached some fourteen feet in length and ten in height. They were American arid apparently rather local. Another odd type were the elotheres, which appeared in North America in the White River stage, and continued into the Miocene. An interpretation of their general appearance is shown in Fig. 540.

Artiodactyls were prominent, represented by various extinct forms, and by ancestral peccaries, camels, ruminants, and the pingularly specialized horned and tusked Protoceras. Protoperas was remotely related to the deer family, and was profusely ind strangely horned, as though in diminutive mimicry of the [p. 804] Dinocerata. The animal was about the size of a sheep, and had but a short career.
The ruminants seem to have been introduced or foreshadowed by the Tragulidae (chevratains) , now represented in Farther India by a slender little ruminant, isolated and scarcely known, the Tragulus, “the scarcely altered survivor of a great tribe which flourished abundantly in Europe, and less so in North America, before the typical and fully differentiated ruminants had made their appearance”[47] Many of the foregoing types were present also in Europe, where there were types not now known in America.
The marine life. If the Vicksburg formation is regarded as Oligocene, the general aspect of the Eocene sea life remained unchanged. In the later stages of the Oligocene, provincialism became pronounced and the correlation of beds, even in the same province, has been the subject of much difference of opinion.[48] this time, the foraminifers had declined greatly, and the fauna was overwhelmingly molluscan.
On the Pacific coast, the Oligocene fauna shows closer relation to the Miocene fauna than to that of the Eocene, and suggests a climate transitional between that of the Eocene and that of the Miocene.[49]
¶ Notes
Dumble, Jour. Geol., Vol. II; also Reports of the Texas Geol<> Survey, and the Austin and Uvalde folios, U. b. Geol. Surv. ↩︎
Dall, 18th Ann. Rept., U. S. Geol. Surv., Pt. II. This classification places the Vicksburg in the Oligocene, instead of associating it with the Jacksonian. There appears to be no physical reason for this separation. ↩︎
Arnold, Jour. Geol., Vol. XVII. ↩︎
Willis, Tacoma folio, U. S. Geol. Surv. ↩︎
Willis, Bull. Geol. Soc. Am., Vol. IX, 1897-8. See also 18th Ann. Rept., U. S. Geol. Surv., and Landes, Washington Geol. Surv., Vol. II, p. 170. ↩︎
Dawson, Science, Vol. XIII, 1901, p. 401. Also Spencer, A. C., Bull. Geol. Soc. Am., Vol. XIV, p. 131. ↩︎
For an account of the deposits near the loth parallel, see King’s report, Vol. I, already cited. For an attempt at correlating the several formations, see Dull, 18th Ann. Rept., U. S. Geol. Surv., Pt. II; also J. H. Smith, Jour. Geol, Vol. VIII, pp. 444-471. ↩︎
Wilder has recently called into question the separability of the Fort Union and Laramie, in western North Dakota. Jour. Geol., Vol. XII, p. 290. ↩︎
For criteria for distinguishing lacustrine and subaerial formations, see Davis, Science, N. S., Vol. VI, p. 619, 1897, and Proc. Am. Acad. Arts and Sci., Vol. XXXV, p. 345, 1900. ↩︎
Purington, Telluride, Colo. Folio, U. S. Geol. Surv. ↩︎
Hills, Science, Spanish Peaks and Walsenburg, Colo. Folios, U. S. Geol. Surv. ↩︎
Loomis, Am. Jour. Sci., Vol. XXIII, 1907. ↩︎
The Green River group of Hayden and Powell, the Wind River group of Hayden, and the Dinoceras beds of Marsh, are included here. ↩︎
Purington, Telluride, Colo., folio, U. S. Geol. Surv. ↩︎
Here belong the Diplacodon beds of Marsh and the Browns Park group of Powell; Geol. of the Uinta Mountains, pp. 63, 168, 208. ↩︎
It is possible that some of these beds should be referred to the Oligocene. ↩︎
Merriam, Jour. Geol., Vol. IX, p. 71, and Bull. Univ. of Cal., Vol. II. p. 285, and Knowlton, Bull. 204, U. S. Geol. Surv. ↩︎
Lindgren and Drake, Nampa and Silver City, Idaho, folios, U. S. GeoL Surv. ↩︎
Knowlton, op. cit., p. 110. ↩︎
Hatcher, Am. Jour. Sci., Vol. IX, 1900, pp. 97-99; also Vol. IV, 1897, pp. 334-337. ↩︎
Branner, Bull. Geol. Soc. Am., Vol. XIII, Stone Reefs of Brazil. Mus. of Comp. Zool., Bull. 44, pp. 27-53. ↩︎
Ameghino, L’age des Formations Sedimentaries de Patagonia, Anales de la Sociedad Crentipica Argentina, 1903. ↩︎
Hill, Geological History of the Isthmus of Panama and Portions of Costa Rica. Bull. Mus. of Comp. Zool., Cambridge, 1898. ↩︎
Neumayr, Erdegeschichte, Bd. II. ↩︎
Penhallow. Tertiary Plants of British Columbia, Can. Rept. of M 1908. ↩︎
Geikie, Text-book of Geology, 3d ed., p. 974. ↩︎
For references to important literature on the American Tertiary Mammalia, see the authors’ larger work, Vol. Ill, p. 228. ↩︎
Osborn, Bull. 361, U. S. Geol. Surv. ↩︎
Mono. XXI, U. S. Geol. Surv., 1893, p. 1. ↩︎
W. H. Dall, Trans. Wagner Free Inst. Sci., Vol. Ill, 1890-1900; Bull. 141. U. S. Geol. Surv., 1896; W. B. Clark, Md. Geol. Surv., Eocene volume, 1901; R. M. Bagg, Bull. 141, U. S. Geol. Surv., 1896; A. Heilprin, Proc. Acad. Nat. Sci. Phil., Vol. XXXI, 1879; Vol. XXXII, 1880; and Vol. XXXIII, 1881; Jour. Acad. Nat. Sci. Phil., Vol. IX, 1884; T. W. Vaughan, Bull. 141, U. S. Geol. Surv., 1896; Mon. XXXIX, U. S. Geol. Surv., 1900; T. W. Stanton, 17th Ann. Rept., U. S. Geol. Surv., Pt. I, 1895-6; G. D. Harris, Am. Pal. Bull., Vols. I and II. ↩︎
Arnold, Jour. GeoL, 1909. ↩︎
Knowlton, Tacoma, Wash. Folio. ↩︎
For table of Oligocene formations, see Dall, 18th Ann. Rept., U. S. Geol. ■v., Pt. II. ↩︎
The classification of the Grand Gulf formation is in dispute. Some of he beds described under this name are probably younger than Oligocene; ee Smith and Aldrich, Science, N. S„ Vol. XVI, p. 836, and Vol. XVIII, p. 26. ↩︎
Penrose, Geol. Surv. of Texas, 1st Ann. Rept. ↩︎
Hill, Geology and Physical Geography of Jamaica, and Geological History of the Isthmus of Panama and portions of Costa Rica. Bull, Mus. Comp. Zo6l., Vols. XXVIII and XXXIV respectively. ↩︎
Darton, Camp Clarke, Scotts Bluff, Edgemont, and Oelrichs folios, U. p. Geol. Surv. ↩︎
Adams, Am. Geol., Vol. XXIX, p. 303. ↩︎
Fraas, Science, Vol. XIV, N. S., p. 212, and Matthew, Am XXXIII, p. 403, 1899. ↩︎
Darton, 19th Ann. Rept., U.S. Geol. Surv., Pt. IV; 21s< Ann ! U. S. Geol. Surv., II. ↩︎
Dall, 19th Ann. Rept., U. S. Geol. Surv.; Merriam, Jour. Geol., V pp. 71-72, and Bull. Univ. of Cal., Vol. II, pp. 270 et Beq. ↩︎
Arnold, Bull. 321, U. S. Geol. Surv. ↩︎
See references to the writings of Hill under Eocene. ↩︎
Willis. Carnegie Institution Year book 4, 1905. ↩︎
Hatcher, See references to this region under Eocene, and especially Geol. Mag., 1902, p. 136. ↩︎
The Tertiary Insects of North America, U. S. Geol. Surv. Ter., Vol. XIII, 1890; Mon. XXI and XL, U. S. Geol. Surv., 1893 and 1900. ↩︎
A. Smith Woodward, Vert. Pal., p. 360. ↩︎
Maury, Bull. Am. Pal., No. 15, Cornell Univ., 1902. ↩︎
Arnold, Jour. Geol., Vol. XVII. ↩︎