| XV. Época Cámbrica y el predominio de los trilobites | Página de título | XVII. Braquiópodos o conchas de lámparas |
[ p. 207 ]
(Véase láminas 4, 7 y 12)
Historia. — Los trilobites fueron los primeros fósiles que atrajeron la atención de los naturalistas y han sido de interés popular durante mucho tiempo. Fue Lhywd, conservador del Museo Ashmolean de Oxford, Inglaterra, quien, en 1698, llamó la atención por primera vez sobre un espécimen completo. No sabía cómo clasificarlo, pero pensó que era el esqueleto de un pez desconocido, y durante mucho tiempo se los consideró escarabajos, orugas, insectos sin alas, mariposas fósiles e incluso moluscos. El gran naturalista sueco, Linneo, fue el primero en reconocer correctamente su relación con los crustáceos, animales como camarones, cangrejos y langostas. En 1771, el profesor Waleh, de Jena (Alemania), les dio el nombre genérico de Trilobites, término que desde entonces se ha elevado al rango de subclase Trilobita de la clase Crustacea del filo Arthropoda, que abarca una gama extremadamente variada de animales con cuerpos y extremidades articulados o segmentados, entre ellos crustáceos, insectos, escorpiones y arañas. Actualmente se conocen más de dos mil tipos de trilobites.
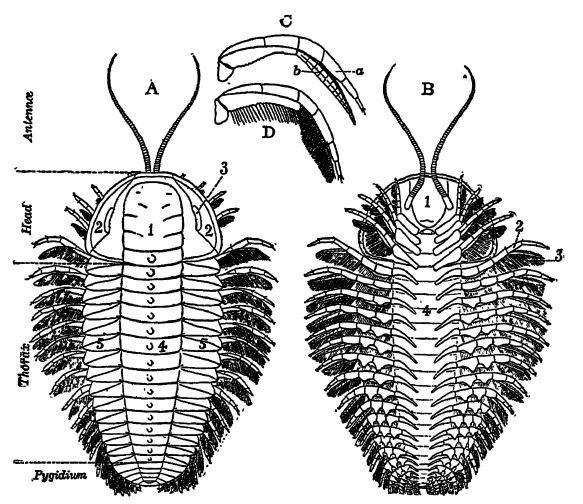
Definición. — La palabra trilobite significa similar a tres lóbulos y hace referencia a los tres lóbulos longitudinales (el eje central y la pleura lateral) que se observan en la cara dorsal o superior de la mayoría de los trilobites (véase la fig., pág. 208). Eran animales sexuados. Sus cuerpos estaban compuestos por segmentos, muchos de los cuales se articulaban entre sí; y estos segmentos se agrupaban en tres dhisiones, como puede observarse en cualquier ejemplar completo del caparazón superior o caparazón, la parte que suele conservarse. La cara inferior, o ventral, con las extremidades, tenía una capa exterior muy fina y solo se conservaba en condiciones excepcionales, normalmente en pizarras negras, más o menos sulfurosas. El caparazón era más grueso y estaba compuesto de quitina (muy similar a la del cuerno, el pelo, etc.), una sustancia nitrogenada a la que se añaden sales de cal. La quitina resiste en gran medida la alteración química, y esta es la razón por la que los trilobites se conservan tan a menudo como fósiles.
Hábitos y hábitat. — Los trilobites habitaban únicamente en aguas marinas, principalmente en aguas poco profundas, y sus testas o conchas externas se encuentran [ p. 208 ] en todo tipo de sedimentos marinos, pero principalmente en lutitas y calizas. En general, eran animales bastante perezosos, flotaban con facilidad, pero probablemente nadaban a sacudidas, y particularmente hacia atrás, ya sea con la parte ventral o dorsal hacia arriba. Sobre el fondo marino se arrastraban lentamente con la ayuda de numerosas patas robustas. Algunos excavaban en la arena en busca de alimento vivo, metiéndose de cabeza, mientras que otros entraban primero con la cola y así, ocultos, esperaban a su presa, lanzándose tras ella repentinamente como peces. Otros se arrastraban por el lodo. Las especies pequeñas de formas muy espinosas podrían haber pasado su vida flotando y nadando en el plancton, mientras que aquellas con ojos grandes podrían haber habitado en las partes más profundas y oscuras de los mares, subiendo por la noche a la superficie en busca de alimento. Se dice que muchos trilobites, y en particular los del Cámbrico [ p. 209 ] de la provincia Atlántica, eran ciegos, sin e 3 'es, o ciertamente sin ninguno en la cara dorsal. Debido a esta ceguera, se cree que vivieron generalmente en aguas muy profundas o incluso abisales, pero los sedimentos en los que se encuentran y los fósiles asociados a ellos indican que también vivieron en mares poco profundos.
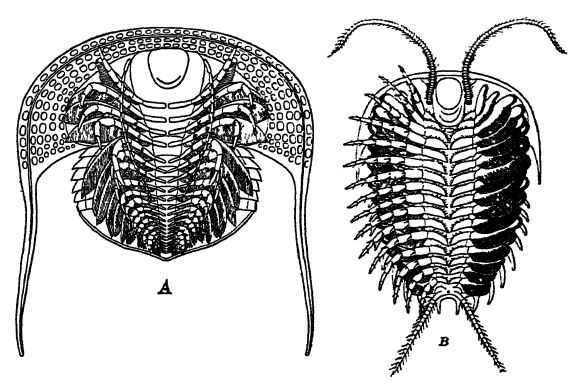
La mayoría de los trilobites podían enrollar sus cuerpos como las cochinillas de la humedad o las cochinillas de nuestros sótanos, animales que no son insectos, sino crustáceos (isópodos) y parientes lejanos de los trilobites. Este enrollamiento protegía las partes más blandas y delicadas de la parte ventral, ofreciendo así al enemigo su duro y grueso caparazón, una armadura eficaz contra otros trilobites, pero ineficaz contra los cefalópodos y los peces. La capacidad de enrollarse apareció por primera vez en el Cámbrico Superior, y con el Champlainiano todas las formas adoptaron este método de protección.
Por regla general, los trilobites eran carroñeros y, como carroñeros, mantenían los fondos marinos limpios de animales muertos; algunos eran omnívoros, otros probablemente totalmente vegetarianos y unos pocos eran «comedores de barro», es decir, el tracto digestivo asimilaba la materia orgánica del barro para su sustento corporal.
A principios del Cámbrico, los trilobites con pigidios grandes, o piezas caudales, comenzaron a ser comunes. Este mayor tamaño, logrado mediante la consolidación de un mayor número de segmentos, se cree que se debió al uso de la cola agrandada como órgano natatorio, moviéndola rápidamente hacia abajo y hacia adelante, lo que les hacía lanzarse hacia atrás como los cangrejos de río y las langostas actuales. El máximo crecimiento pigidial y la mayor abundancia de formas que lo poseen se alcanzaron en el Champlainiano. Esta tendencia hacia una cola más grande parece haberse desarrollado un poco antes de la primera abundancia de cefalópodos, y como estos animales probablemente se alimentaban de los trilobites, se cree que estos últimos desarrollaron, en defensa propia, no solo un modo de natación más rápido, sino también uno hacia atrás, una nueva adición locomotora a las abundantes patas.
Tamaño. — El tamaño de los trilobites en la madurez varió considerablemente, con longitudes que oscilaban entre 0,95 y 70,9 cm, pero el tamaño promedio era de unos 3,8 cm. Muchas especies alcanzaron longitudes de 7,62 a 10,16 cm e incluso 15,24 cm, pero se trataba de individuos grandes, y los de más de 30 cm eran gigantes. Los ejemplares representados de entre 45,72 y 70,92 cm se basan, en casi todos los casos, en restauraciones realizadas a partir de fragmentos grandes.
Duración geológica. — Los trilobites fueron característicos de la era Paleozoica, comenzando con una variedad considerable en el Cámbrico Inferior (alrededor de 37 géneros y 125 especiales) y dominando los mares del [ p. 210 ] Cámbrico (alrededor de 95 + 625) y el Champlainiano (más de 120 + 1050). En el Silúrico, aunque todavía eran comunes, los trilobites estaban, sin embargo, en declive (35 + 500), y este reflujo de su fuerza vital se muestra aparentemente en muchas formas pintorescas repletas de protuberancias, espinas y exageración de partes. Como regla general, en la evolución, uno encuentra que cuando un tronco orgánico está perdiendo su vitalidad surge en él una exageración de partes, como si se estuvieran haciendo esfuerzos heroicos para mantener la raza. La espinosidad en los animales es a menudo la profecía de la muerte tribal. En el Devónico, la variedad y el número de los trilobites se redujeron considerablemente (30 + 120), en un momento en que los antiguos Los peces, que sin duda se alimentaban de estos crustáceos, comenzaron a ser comunes en los mares. En los mares del Paleozoico tardío, los trilobites eran reliquias, o animales sobrevivientes de una época más adaptada a sus necesidades, y uno a uno desaparecieron, hasta que poco antes del final de la era Paleozoica no quedó ninguno. En su apogeo, dominaron la vida de los mares, ya que fueron los principales carnívoros y carroñeros del Cámbrico y el Champlainiano. Sin embargo, con el auge de los cefalópodos y los animales parecidos a peces en el Silúrico y los peces auténticos en el Devónico, su sentencia de muerte fue anunciada por la llegada de una mayor mentalidad y alerta de estos tipos superiores.
Definición de las Partes (estudio con Fig., pág. 208). — El caparazón de los trilobites se divide transversalmente en (1) una porción cefálica, llamada cefalón; (2) una serie de segmentos que forman el tórax; y (3) una cola o pigidio, que forma el abdomen. El eje es el área de los órganos vitales, mientras que las regiones pleurales son las cubiertas protectoras de las extremidades ventrales.
El cefalón o cabeza es la parte más importante del animal y, en la clasificación del grupo, presenta las características más fiables para la distinción ordinal y genérica. Está compuesto por una glabela medial y mejillas libres externas, donde se encuentran los ojos compuestos. En muchas formas, las mejillas libres están separadas de la glabela por una sutura facial que generalmente se aprecia como una línea tenue en relieve. En todas estas formas, las tres partes tienden a separarse tras la muerte, dando lugar así a fósiles imperfectos. En algunos trilobites no se conocen ojos compuestos, por lo que estas formas se denominan ciegas; sin embargo, normalmente presentan dos ojos compuestos más o menos grandes. En la mayoría de los trilobites, cada ojo compuesto está cubierto por una sola córnea, y las lentes individuales que se encuentran debajo pueden o no ser visibles. En otros, cada lente tiene su propia córnea. El número de lentes en un ojo compuesto puede variar desde catorce hasta la asombrosa cifra de quince mil. ¡Imagina un animal con treinta mil ojos!
[ p. 211 ]
El tórax consta de un número variable de segmentos articulares (en el trilobites más primitivo, Naraoia, no hay ninguno). Este número es constante en los adultos de cada especie, pero variable en las crías de cada forma y en los géneros. Con una excepción, nunca hay menos de dos, como en Agnostus (Lámina, pág. 201, Fig. 4), ni más de cuarenta y cuatro. En general, se puede decir que, cuando hay muchos segmentos torácicos, el pigidio es pequeño, y cuando hay pocos, el abdomen es grande. Cabe señalar que, mientras estos animales crecen, los nuevos segmentos torácicos añadidos durante las etapas de muda se derivan de la descomposición del pigidio.
El pigidio o abdomen de los trilobites puede ser pequeño o grande, y consiste en una sola pieza de segmentos unidos, cuyo número varía entre dos y veintiocho. Generalmente, las regiones axial y pleural están claramente delimitadas, y algunas formas muestran claramente el número de segmentos fusionados.
En Neolenus, desde el pigidio se extienden dos cercópodos que probablemente se utilizaban como órganos sensoriales, al igual que las antenas (Fig. B, arriba). Estos cercópodos podrían tener su origen en las patas reptantes. Si bien solo se conocen [ p. 212 ] cercópodos en este género, es posible que otros trilobites de cola pequeña los tuvieran, pero en las formas de cola grande se perdieron cuando el pigidio se convirtió en un órgano natatorio.
Lado ventral. — Debajo de la glabela y articulado a su margen posterior curvado y engrosado se encuentra el hipostoma o labio superior que cubre la boca (Fig. 5, pág. 208). A cada lado de esta placa hay un par de órganos sensoriales o sensitivos, con antenas cortas o largas y multiarticulados. Todos los miembros son pares, y cada miembro es doble o birrámeo, con cada una de las dos ramas, a su vez, compuesta por múltiples articulaciones.
La cabeza posee cuatro pares de extremidades birrames que también sirven para masticar el alimento y llevarlo a la boca. En la parte posterior de la cabeza, cada segmento torácico y abdominal posee su par similar de extremidades birrames. Ambas ramas de las extremidades surgen de una única articulación basal unida a los apendíferos, que son pliegues de la concha dorsal del raquis. La rama inferior, más robusta y con menos articulaciones, sirve para reptar, mientras que la otra, más delicada y articulada, se utiliza para nadar y también está repleta de largos apéndices pilosos (setas), que son delicados conductos por los que la sangre circulante extrae del agua el oxígeno necesario para su sustento (Fig. D, p. 208). Actualmente se conocen extremidades en doce especies y nueve géneros de trilobites, que representan tres órdenes y siete familias.
Muda de Trilobite. — A medida que los trilobites crecían, mudaban periódicamente su caparazón quitinoso, al igual que los cangrejos, y esto sucedía muchas veces antes de alcanzar el crecimiento completo y probablemente ocasionalmente durante la madurez. Durante el tiempo de muda, los animales crecían y las características detalladas cambiaban, junto con la introducción de nuevos segmentos. Estos nuevos segmentos se añadían justo delante del anal, y en el momento de la muda uno o más de los segmentos anteriores del pigidio pueden haberse desprendido y añadido al tórax. Al principio, el trilobite era un animal muy diminuto, aparentemente no segmentado, todo cabeza (Lám., pág. 242, Figs. 14, 15); después consistió en cabeza y pigidio, este último añadiendo rápidamente más segmentos; estas etapas fueron seguidas por otras mudas que mucho antes de alcanzar la madurez introdujeron rápidamente el número completo de segmentos torácicos y pigidiales según las características de cada especie. Las mudas posteriores simplemente agrandaron a los individuos.
Evolución. — Los trilobites son los crustáceos más primitivos y generalmente se asume que se desarrollaron a partir de un anélido multisegmentado en el Proterozoico. Sin embargo, recientemente, Raymond ha postulado que el progenitor de los trilobites fue un animal de cuerpo blando, deprimido, capaz de flotar y nadar, similar a un gusano, compuesto de unos pocos segmentos, probablemente con ojos marginales simples o incluso ciegos, con una boca bajo el margen anterior, órganos táctiles en uno o ambos extremos, forma ovalada y un intestino recto y estrecho que se extiende desde la boca anterior hasta el ano terminal.
Probablemente fue el tipo de animal mencionado el que dio origen, en algún momento del Proterozoico, a los trilobites, de los cuales surgieron algunos de los crustáceos superiores. Por lo tanto, podemos afirmar, con Raymond, que los trilobites dieron origen, directa o indirectamente, a todos los demás artrópodos.
Para una discusión completa de la tendencia evolutiva entre los artrópodos, el estudiante puede consultar el libro de Raymond.
[ p. 213 ]
¶ Lectura colateral
P. E. Raymond, Los apéndices, la anatomía y las relaciones de Trilo Memorias de la Academia de Artes y Ciencias de Connecticut, vol. 7, 19
C. D. Walcott, Apéndices de trilobites. Lecciones misceláneas del Smithsonian, vol. 67, 1918, págs. 115-217.
C. D. Walcott, Notas sobre la estructura de Neolenus. Ibíd., Vol. 67, págs. 365-457.
| XV. Época Cámbrica y el predominio de los trilobites | Página de título | XVII. Braquiópodos o conchas de lámparas |