| XVII. Braquiópodos o carcasas de lámparas | Página de título | XIX. Época Champlainiana y el reinado de los animales invertebrados. |
[ p. 220 ]
(Véase láminas 9, 12, 22, 44)
A menudo se habla de los moluscos como fl.niTn fl.ls con concha, aunque muchos de ellos carecen por completo de una concha externa dura. Originalmente, todos tenían concha, pero a lo largo de las eras geológicas, muchas especies abandonaron la rígida armadura exterior para poder desplazarse con mayor facilidad. Los moluscos son una gama muy diversificada de animales con simetría bilateral, y se estima que actualmente viven unas 45.000 especies. Entre ellas se encuentran las almejas, las ostras, los caracoles y el nautilus perlado. Su mayor diversidad y abundancia se encuentra en los mares y océanos. Son menos variados y comunes en aguas dulces, mientras que los caracoles que respiran aire solo viven en tierra firme donde hay vegetación.
El término molusco significa blando, ya que sus cuerpos son muy blandos y carecen de esqueleto interno. Viven en conchas para protegerse de sus enemigos. Sus cuerpos no están segmentados como los de los gusanos y, por lo general, estos animales son muy perezosos. Aunque carecen de extremidades para arrastrarse o nadar, la mayoría se desplaza mediante el llamado pie, una planta rastrera situada bajo el vientre. Generalmente, el cuerpo está cubierto por un manto, una fina membrana que secreta la concha cuando esta está presente. Dentro de la cavidad del manto se encuentran las branquias, órganos de diversa estructura que generan corrientes de agua, de las cuales se extrae no solo el oxígeno para la respiración, sino también, a menudo, el alimento necesario para la vida. Los caracoles terrestres carecen de branquias, y en estos casos, la cavidad del manto se ha adaptado a una especie de pulmón para la respiración aérea, ya que respiran aire. En todos los moluscos, excepto en los bivalvos, la boca está provista de una cinta quitinosa flexible conocida como radvla, una especie de lengua, generalmente provista de numerosos dientes diminutos, afilados y duros, y accionada por músculos especiales para desmenuzar el alimento. Los sexos están separados o unidos, siendo en este último caso los individuos hermafroditas.
Naturaleza de la Concha. — Las conchas suelen constar de tres capas: una capa exterior, muy fina, de color marrón y córnea, siempre [ p. 219 ] ausente en los fósiles; una capa media, gruesa y calcárea, denominada capa porcelana, la que se conserva con mayor frecuencia en las rocas; y una capa interior, generalmente gruesa, de nácar, que a menudo se destruye en los fósiles, especialmente entre los moluscos paleozoicos, ya que consiste en una forma fácilmente soluble de carbonato de cal, conocida como aragonito.
Clasificación de los Moluscos. — La clase más primitiva de moluscos son los Lamelibranquios, bivalvos como las almejas y las ostras. Sin embargo, son primitivos solo porque son degenerados, pues en el curso de su evolución perdieron la cabeza y los ojos, al estar completamente encapsulados entre dos pesadas conchas, lo que los ha convertido en animales más o menos sedentarios. Originarios de aguas marinas, se han extendido a aguas dulces, pero ninguno ha logrado sobrevivir en tierra firme.
El tronco primigenio que dio origen a los moluscos aún no se ha encontrado en el Proterozoico, pero en el Cámbrico los gasterópodos o caracoles son los moluscos comunes. De esto y de la embriología de los seres vivos, es evidente que este clan, en su estructura, es el más cercano al tronco primigenio de los moluscos. A lo largo de las eras geológicas, los gasterópodos, en su evolución, constituyen un grupo ascendente de organismos y, en la actualidad, están más diversificados que nunca. Incluyen los caracoles y los taladros.
De los gasterópodos surgieron los moluscos más altos y, por lo tanto, los más complejos e inteligentes: los cefalópodos, como el nautilus perlado, los amonites, los calamares y los peces diablo. Los dos primeros grupos mencionados poseen conchas, las cuales siempre presentan abundantes cámaras, mientras que los calamares y los peces diablo carecen de ellas, aunque pueden presentar una especie de esqueleto interno.
Importancia de la Evolución de los Moluscos. — El vasto reino de los Moluscos tuvo un buen comienzo, con independencia física, plenamente equipado con poderes locomotores y una excelente nutrición. Sin embargo, todos, excepto los cefalópodos, vendieron su derecho de nacimiento a cambio de comodidad y bienestar. Incluso los cefalópodos, con el marcado grado de independencia observado en los calamares y los peces diablo, no dieron origen a una estirpe superior. Los gasterópodos, y especialmente los lamelibranquios, se volvieron dependientes del movimiento del agua y esperaron a que las olas les trajeran alimento. Seguros de su protección y adaptación, estos tipos de vida se han extendido a lo largo de los siglos en una variedad indescriptible, pero de ellos no puede surgir nada mejor. Tuvieron su oportunidad al comienzo de su ascenso, pero la desaprovecharon, y durante incontables millones de años no han logrado mejorar. (J.M. Clarke, 1917. Véase también su obra Organic Dependence and Disease, 1921).
[ p. 221 ]
¶ Lamelibranquios o bivalvos
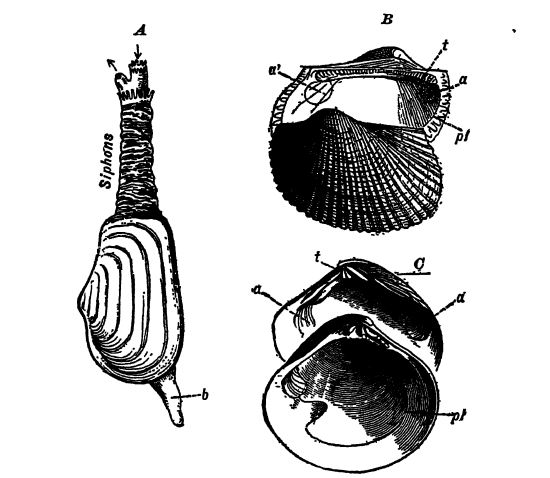
El nombre Lamellibranchia proviene de lamella, que significa hoja, y branchia, que significa branquias, y fue elegido porque en estos moluscos las branquias son similares a las hojas. La clase también es ampliamente conocida como Pel_ecypoda, que significa pie de hacha, porque el pie a menudo sugiere la forma de un hacha (Figs. A, abajo, y A, p. 222). El grupo incluye todos los moluscos sin cabeza y, por lo tanto, degenerados. El cuerpo generalmente está comprimido de lado a lado y siempre cubierto por dos conchas o valvas, una a la derecha y la otra a la izquierda (Figs. A y B, abajo). Por lo tanto, a menudo se les llama bivalvos, y popularmente se les conoce como mejillones, almejas, ostras, berberechos y vieiras. Su hábitat es principalmente marino, en las aguas poco profundas cerca de las costas, donde su alimento, las algas microscópicas, es más abundante. Casi todas las aguas dulces, sin embargo, tienen algunos mejillones, pero ninguno ha logrado sobrevivir fuera del agua, en tierra, como lo hacen los caracoles. El alimento se filtra del agua mediante las branquias (en realidad, los palpos labiales), que también son los órganos respiratorios. Los lamelibranquios son animales muy perezosos, muchos de los cuales permanecen enterrados en el lodo y el fango del mar (Fig. A, p. 222). Las vieiras solo pueden nadar un poco, batiendo sus valvas y forzando el agua alternativamente de un lado a otro.
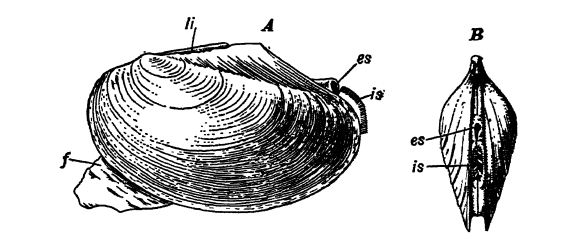
Características de la concha. — Las dos conchas están unidas en el lado dorsal del cuerpo por una bisagra, un área más o menos engrosada provista de dientes y alvéolos, que permite que las valvas se muevan una sobre la otra (Figs. B y C, p. 222). Ya sea en el exterior o dentro de la concha engrosada (majgg) del área de la bisagra, hay un ligamento elástico unido a ambas valvas que actúa como un resorte y mantiene las conchas abiertas cuando los animales están en reposo (Fig. A, arriba). En el interior de las valvas [ p. 222 ] hay cicatrices musculares, generalmente una en cada extremo de la concha, que se deben a los músculos aductores anterior y posterior, aunque en muchas formas, como en las ostras, solo hay una de estas cicatrices situada cerca del centro de la valva (Figs. B y C, abajo). Estos músculos cierran la concha, y cuando está cerrada, el ligamento elástico está bajo tensión listo para abrir las valvas de nuevo tan pronto como los aductores dejen de tirar. El pie en forma de cuña de los lamelibranquios es grande en las formas rastreras, pero más o menos reducido en tamaño por el desuso, según que los animales vivan en agujeros, como la almeja blanda, o tengan una concha firmemente cementada a algún objeto duro en el fondo del mar, como en las ostras.
En contraste con los braquiópodos, que también poseen dos conchas, se observa que en este grupo las valvas se encuentran en las caras dorsal y ventral de los animales, mientras que en los lamelibranquios se encuentran en los lados derecho e izquierdo. Los braquiópodos poseen un complejo sistema [ p. 223 ] muscular para abrir y cerrar las conchas, mientras que los lamelibranquios solo lo tienen para cerrarlas; se abren mediante el ligamento elástico. En los braquiópodos existe un pedúnculo que los mantiene sujetos al suelo, mientras que los bivalvos casi siempre tienen libertad de movimiento.
Sifones. En la mayoría de los lamelibranquios, el manto se extiende más allá de la concha en dos tubos o sifones por los que el agua de la respiración entra y sale de la cavidad del manto (Figs. A, pág. 221 y A, pág. 222). Cuando los sifones son grandes, se trata de una adaptación para vivir enterrados en la arena o el lodo, y todos los lamelibranquios con estos sifones tienen las conchas marcadas en el interior por una línea marcadamente curva, el seno paleal, que se observa en conexión con el músculo aductor posterior.
Distribución geológica. — Los lamelibranquios no se establecieron bien hasta el Champlainiano, época en la que se volvieron cada vez más abundantes y son muy prolíficos en todas las aguas someras de los océanos actuales. Si bien las conchas de estos bivalvos suelen ser abundantes en las rocas del Paleozoico, no suelen estar bien conservadas hasta el Pensilvánico, una condición aparentemente relacionada con una capa porcelánica bastante delgada y una de nácar comparativamente gruesa, siendo esta última fácilmente destructiva. Solo son ocasionalmente útiles como marcadores estratigráficos en el Paleozoico, pero, a partir del Mesozoico, se conservan abundantemente y a menudo sirven como fósiles índice.
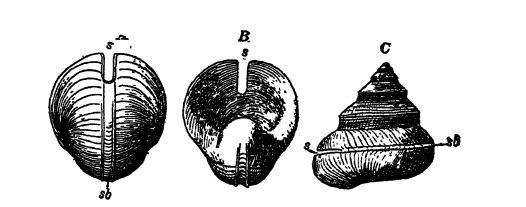
¶ Gasterópodos o caracoles
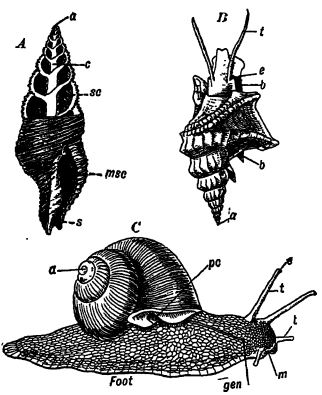
Características generales. — El nombre Gastropoda significa «pies estomacales» y se refiere a que estos animales se desplazan mediante contracciones de la planta del pie, que se encuentra en la cara ventral (Figs. B y C, pág. 224). Esta clase de moluscos, que por lo general son lentos, aunque menos que los lamelibranquios, [ p. 224 ] es extremadamente variada y abarca no solo formas con concha como lapas, drils, bígaros, buccinos, caracoles y caracoles, sino también babosas marinas desnudas y babosas terrestres. Son más abundantes y variados en mares poco profundos, donde se arrastran por el fondo. Son mucho menos variados en agua dulce. En tierra firme, están representados por los caracoles vegetarianos, aerofíticos o pulponatos (llamados así porque la cavidad del manto actúa como un pulmón en la respiración, véase la Fig. C, lado opuesto), que se encuentran en casi todos los lugares donde crecen plantas y son muy variados en todo el mundo. La cavidad del manto en las formas acuáticas suele tener una sola branquia, la del lado derecho, ya que la del lado izquierdo se ha perdido debido a la forma espiral del cuerpo. Por lo general, son herbívoros, y con menos frecuencia carnívoros y carroñeros. La boca está provista de una rádula o proceso similar a una lengua. Las formas carnívoras, conocidas como taladros, perforan agujeros en sus víctimas, como las almejas, con la rádula y la ayuda de una secreción débil de ácido sulfúrico. La cabeza de la mayoría de los gasterópodos suele estar claramente diferenciada del resto del cuerpo y tiene un par de ojos y uno o dos pares de órganos sensoriales o tentáculos (Figs. B y C, arriba).
Concha espiral. — La forma usual de la concha o cubierta protectora es un cono dextral, retorcido en espiral, con el ápice dirigido hacia arriba (Fig. C, arriba); es secretado por el manto situado en el lado dorsal del animal. Debido a esta concha única, a los gasterópodos también se les llama moluscos univalvos. Sin embargo, en muchas formas, hay un opérculo quitinoso o calcáreo, [ p. 225 ] una placa unida al lado del pie, que cierra la abertura de la concha cuando el animal se ha retirado a su concha o está retraído. Aunque los gasterópodos pueden tener dos conchas, estas juntas no representan las dos valvas de los bivalvos. Es solo el cono espiral el que corresponde a las valvas, porque el opérculo es una estructura secundaria que no está presente en todos los univalvos, es secretado por el pie y no tiene nada que ver con el manto dorsal o la secreción verdadera de la concha.
Distribución geológica. — Las formas primitivas de gasterópodos estuvieron presentes en el Cámbrico Inferior, donde prácticamente todos eran más o menos cónicos, no retorcidos, sino con forma de capuchón. El tipo de cono retorcido aparece con mayor abundancia en el Cámbrico Superior y, desde el Champlainiano, ha sido común en fósiles. Actualmente, se conocen más de 20 000 especies, una representación mayor que en cualquier otro momento del pasado geológico. En el Palezoico, por regla general, son fósiles defectuosos, y solo a partir del Mesozoico su abundancia en especímenes bien conservados adquiere valor en la geología histórica. Los gasterópodos aerodinámicos aparecen con poca frecuencia en el Pensilvánico y parecen no haber sido comunes hasta finales del Jurásico, donde también se observan los primeros caracoles de agua dulce. Se conocen unas 500 especies de caracoles pulmonados fósiles y unos 6000 tipos de caracoles terrestres.
¶ Cefalópodos
Los cefalópodos (que significa con cabeza y patas) son los moluscos más altamente organizados e incluyen animales como el nautilo, los amonites, los pulpos, las sepias y los calamares. Todos son exclusivamente marinos. Los dos primeros tipos de cefalópodos tienen conchas con cámaras en las que se encuentran cinco, mientras que los segundos carecen de ellas. Para los fines presentes, solo se describirán los nautilos, ya que son claramente característicos del Paleozoico, y el resto se considerará al estudiar los animales del Mesozoico.
El nautilus perlado (véase la fig., pág. 226) recibe su nombre del hecho de que sus hermosas conchas, bilateralmente simétricas, se utilizan a menudo como adornos y para botones de nácar, tras retirar la fina capa exterior, porcelánica y rayada, de la capa interior, más gruesa y de nácar. La palabra nautilus es una forma poética del término griego para marinero, y al igual que sus parientes lejanos, los argonautas, se creía que estos animales podían extender algunos de sus brazos al viento y así surcar los mares. Sin embargo, esta idea es errónea, ya que las cuatro especies vivas del género Nautilus que habitan actualmente en Oceanía, desde la región malaya hasta [ p. 226 ] las islas Filipinas y Fiyi, nunca han sido vistas en la superficie, excepto en estado de agonía, aunque sus conchas vacías y selladas se desplazan a Japón, África y Nueva Gales del Sur. Los naútilidos son animales gregarios de aguas profundas y cálidas, entre 90 y 180 metros, a una temperatura de 20 °C, aunque se han capturado a 300 metros de profundidad. Son feroces y decididamente carnívoros, y capturan la carne que se usa como cebo en las nasas a profundidades de hasta 128 metros. Los sexos siempre están claramente diferenciados.
Los cefalópodos también se conocen como conchas con cámaras, porque el cono, sea cual sea su forma, está regularmente dividido por delgadas particiones transversales perladas llamadas septos. Estos septos son cóncavos hacia la abertura de la concha y están perforados por un orificio circular. A través de estas perforaciones pasa un tubo carnoso conocido como sifón (Fig., arriba, ss), que se origina en la cara posterior redonda del animal y continúa a través de todas las cámaras hasta la última, diminuta, en el ápice del cono, donde se fija a la cara interna de la pared. En muchas formas fósiles, el sifón ha secretado un tubo calcáreo, conocido como sifúnculo, que a veces es muy pequeño. En vida, las cámaras están llenas de un gas que las hace más o menos flotantes, [ p. 227 ] y así, el peso de la casa de piedra apenas supone un estorbo. Los nautilidos se arrastran por el fondo del mar o se sujetan a él mediante sus tentáculos retráctiles, y pueden nadar rápidamente hacia atrás de forma espasmódica, expulsando el agua de la cavidad del manto a través del hipónomo o embudo ambulatorio. A los lados del cono, los septos están firmemente unidos, pero en los fósiles suele ocurrir que se pierda la concha exterior, en cuyo caso los bordes de las particiones se muestran claramente como líneas en el cono relleno de roca. La naturaleza de estas líneas es de gran importancia para la clasificación de los cefalópodos con concha, y se denominan líneas de sutura. Por lo general, estas líneas de sutura son rectas, pero hay formas en las que están decididamente curvadas en ondulaciones. Estas, cuando son convexas hacia el vértice del cono o dirigidas hacia atrás, se denominan lóbulos, y cuando se curvan hacia el extremo más grande o la abertura de la concha, se denominan sillas de montar.
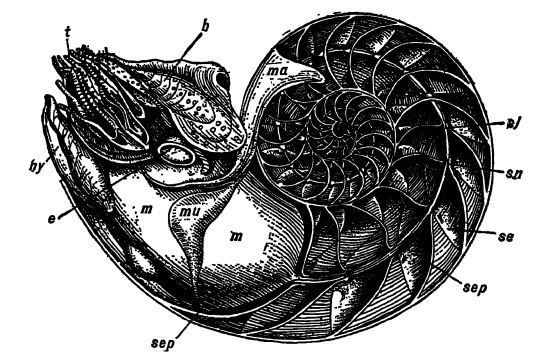
Debajo del saco visceral se encuentra la gran cavidad del manto, con sus plumas branquiales, donde el agua entra y sale libremente a través del embudo. Sin embargo, cuando el animal desea nadar, las dos aletas del manto se aprietan, formando el embudo, y mediante una rápida contracción del manto o cámara branquial, el agua inhalada sale disparada con gran fuerza. El animal entonces se lanza hacia atrás o hacia los lados, según la dirección en la que expulsa el agua. Son nadadores expertos y rápidos. Los ojos, a ambos lados de la cabeza, son grandes, de estructura primitiva y se apoyan en cortos pedúnculos. La boca está rodeada de tentáculos y cuenta con dos picos potentes, protuberantes y homicidas, similares a los de un loro, el superior de los cuales, entre las formas fósiles, solía estar rematado con un pico de carbonato de cal.
Presencia geológica. — Los nautilidos son la división más antigua y primitiva de los cefalópodos. Aparecieron con el Cámbrico, fueron bastante abundantes en el Cámbrico tardío y aumentaron en número en el Champlainiano. Los más primitivos eran conos rectos y ahusados de contorno circular u ovalado, y las numerosas familias de ellos se llaman Orthoceracones (del género Orthoceras, que significa cuerno recto, Pl., p. 236, Fig. 19). Estos ortocéridos fueron comunes a lo largo del Paleozoico y particularmente en el Champlainiano y el Silúrico, cuando se conocen individuos que originalmente tenían una concha de hasta 15 pies de largo (Endoceras). Con el Devónico, estas formas primitivas de concha recta comenzaron a disminuir lentamente, pero algunas todavía estaban presentes en el Triásico.
Los nautilidos rectos y enroscados persistieron hasta el Mesozoico y fueron más abundantes en la primera mitad del Mesozoico, tras lo cual fueron disminuyendo gradualmente. En los océanos actuales solo existen las cuatro formas relictas, que nos proporcionan el único conocimiento de las partes blandas de los miles de especies que poblaban los mares antiguos.
[ p. 228 ]
Tipos de conos de nautílidos. — Todos los tipos superiores de nautílidos se originaron en los Orthoceracones de concha recta. Los primeros descendientes tenían sus conchas ligeramente curvadas y, por lo tanto, se denominan Cyrtoceracones (de Cyrtoceras, que significa horri curvado, Lám., pág. 236, Fig. 21); los posteriores se enrollaban en una espiral suelta en un plano y se conocen como Gyroceracones (de Gyroceras, que significa hcrm redondo); otros están enrollados apretadamente, con los verticilos abrazándose entre sí más o menos estrechamente, como en Nauiilus, y estos se denominan Nautilicones. En los lados de estos se puede ver más o menos de los verticilos internos de las conchas, y el área de estos verticilos se conoce como el ombligo. Es pequeño en Nautilus y ancho o abierto en las formas Champlainian (Lám., pág. 236, Figs. 23, 24). La curvatura de los tubos se debe a una secreción más rápida de cal a lo largo del lado ventral del cono, y cuanto mayor es el crecimiento desigual, más rápidamente se enrolla el cono.
Clasificación. — Los cefalópodos se dividen en dos subclases, los tetrabranquios y los dibranquios, según el número de branquias en forma de pluma que utilizan para oxigenar la sangre. En la primera subclase hay cuatro branquias (tetra, cuatro), y la segunda división tiene solo dos. Los tetrabranquios, todos con concha externa, aparecieron en el Cámbrico y prácticamente han desaparecido de la faz de la tierra, mientras que los dibranquios, que en casi todos los casos están desnudos, pero pueden tener vestigios internos de la concha externa ancestral, surgieron en el Triásico y están abundantemente representados entre los cefalópodos actuales por los calamares, los argonautas y los pulpos.
Hábitos de los Nautilidos Fósiles. — En sus estudios sobre los nautilidos paleozoicos, Ruedemann afirma que, a partir de la naturaleza de las bandas de color preservadas, parece que, entre las formas con conchas rectas sostenidas horizontalmente, algunas se arrastraban lentamente hacia adelante sobre el fondo del mar, o cuando se encontraban en apuros, retrocedían rápidamente gracias a la ayuda del sifón. Entre los nadadores más activos, las conchas más pequeñas y curvadas se sostenían oblicuamente hacia arriba, y las enroscadas, verticalmente. En el nautilus actual, la concha de la hembra tiene una forma ligeramente diferente y un tamaño mayor que la del macho, y esta diferenciación sexual puede distinguirse en algunas de las formas paleozoicas. Las más grandes se consideran hembras y las más pequeñas, machos.
¶ Lectura colateral
Fechas AM, Introducción a la Paleontología. Londres (Murby and Co.), 1920.
Zittel-Eastman, Libro de texto de paleontología, vol. 1, 2.ª ed. Nueva York (Macmillan), 1913.
James Hall, Lamellibranchiata. Paleontología de Nueva York, Vol. 5, Pt. 1, 1884-1885.
C. O. Dunbar, Fases de la adaptación de los cefalópodos. En «Adaptación orgánica», próxima publicación de Yale University Press.
R. Ruedemann, Cefalópodos de las formaciones Beekmantown y Chazy de la cuenca de Champlain. Museo Estatal de Nueva York, Boletín 90, 1906.
R. Ruedemann, Observaciones sobre el modo de vida de los cefalópodos primitivos. Boletín de la Sociedad Geológica de América, vol. 32, 1921, págs. 315-320.
| XVII. Braquiópodos o carcasas de lámparas | Página de título | XIX. Época Champlainiana y el reinado de los animales invertebrados. |