| XXIX. El carbón y su presencia en la naturaleza | Página de título | XXXI. El Pérmico y su clima glacial |
[ p. 405 ]
A mediados del Paleozoico, y especialmente en el Devónico, cuando las tierras secas quedaron completamente cubiertas de plantas —el sustrato del que dependen todos los animales para su alimentación— se estableció un hábitat capaz de sustentar la vida animal, destinado a ser dominado por gusanos, caracoles, arañas, insectos y otros invertebrados, y finalmente por diversos grupos de vertebrados. Entre estos últimos, los primeros en encontrar la tierra habitable fueron los anfibios ancestrales, quienes dominaron sus diversos entornos desde finales del Devónico hasta bien entrado el Pensilvánico. De estos animales surgieron otros más complejos, una maravillosa variedad de reptiles, que no solo se convirtieron en amos de las tierras, mares y océanos antes del final del Pérmico, sino que poco después invadieron el aire, y por su dominio dieron al mundo mesozoico o medieval el título de la Era de los Reptiles.
¶ Anfibios, vivos y fósiles
Los anfibios se diferencian de los peces en que tienen patas en lugar de aletas, y cada pata cuenta con dedos. Casi todos respiran por branquias en su juventud y pueden conservar estos órganos durante toda su vida; sin embargo, los pulmones y las fosas nasales funcionales casi siempre están presentes en el adulto, aunque entre las salamandras, las extremidades pueden estar reducidas a vestigios o completamente suprimidas (véase lámina, pág. 407, figs. 1-4). Tienen un corazón de tres cámaras y una lengua muscular móvil.
Anfibios vivos. — Existen alrededor de novecientas formas de anfibios vivos, la mayoría de las cuales pertenecen al género rana (Fig., pág. 406). Ranas, sapos, tritones, sirenas, cachorros de lodo, perros de agua y salamandras terrestres son tipos de anfibios vivos (Lám., pág. 407). Todos son animales de sangre fría. El nombre de clase Anfibios, que significa que viven una doble vida, se les dio porque muchos viven tanto en la tierra como en agua dulce. Otros, sin embargo, viven completamente en el agua y, por regla general, todos los anfibios en sus etapas más jóvenes se limitan por completo a este elemento.
Los sexos siempre están separados y, en la gran mayoría de las especies, los pequeños huevos se fecundan en el agua y se [ p. 406 ] desarrollan allí %%0%% sin necesidad de cuidados adicionales por parte de los padres. Por lo tanto, el desarrollo es muy similar al de los peces y muy diferente al de los vertebrados superiores. Los huevos de ranas y sapos, por ejemplo, se convierten en pequeños animales conocidos como renacuajos o renacuajos, con una cabeza y un cuerpo más o menos grandes y redondeados que terminan en una cola larga y muy flexible, que mueven al nadar, al igual que los peces (véase la figura inferior). Estos renacuajos tienen branquias que al principio sobresalen de los lados de la cabeza, pero que posteriormente quedan cubiertas por un opérculo. En unos dos meses alcanzan una etapa equivalente a la de los peces pulmonados. Luego experimentan una metamorfosis pronunciada: primero aparecen las patas traseras y, posteriormente, las delanteras, que quedan ocultas bajo el opérculo. La larga cola se acorta por absorción interna y las branquias también se absorben o se desprenden. Luego aparecen los pulmones, y durante un tiempo las crías son completamente anfibias, respirando agua por las branquias y extrayendo el oxígeno libre, además de tomar aire por los pulmones; pero pronto las pequeñas ranas o sapos se instalan en tierra y respiran únicamente aire (véase la fig. superior). En esta metamorfosis, que requiere desde unas pocas semanas hasta un máximo de unos meses, vemos una recapitulación de la historia paleozoica que consumió milenios de selección de los más aptos para su entorno. En las formas que permanecen permanentemente en el agua, la transformación no es tan grande, y algunas se asemejan a los peces en su forma corporal durante [ p. 407 ] [ p. 408 ], entre ellas los llamados cachorros de barro y perros de agua, comunes en los ríos de los valles de Ohio y Alississippi (véase la lám., pág. 407, fig. 1).
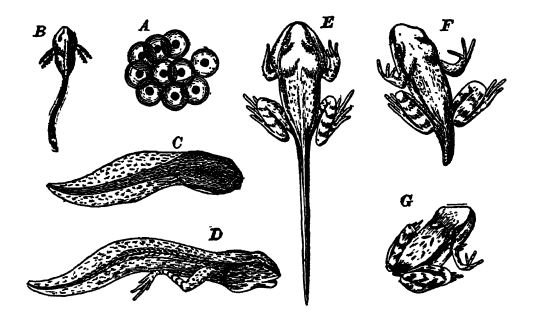
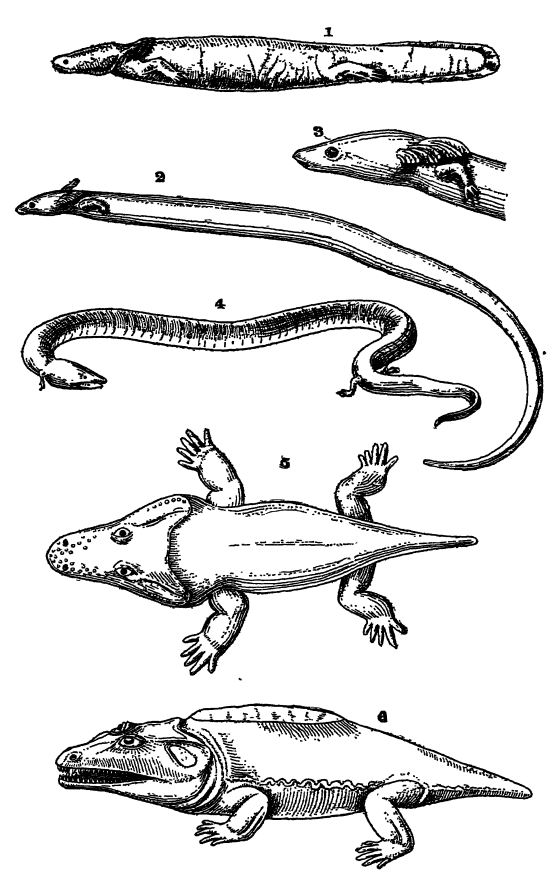
Fig. 1, mud-puppy (Necturus maculaius) of the Ohio and Missisippi rivers; 2 and 3, Siren lacertina, a small salamander, note the three pairs of gills and the absence of hind legs; 4, Amphiuma tridactyla, with both gills and lungs; 5, Trematops milleri, a stegocephaJian, 3 feet long, from Texas; 6, Cacops aspidephorus, a stegocephalian, 20 inches long, from Texas. Figs. 1 and 4 from Parker and Haswell; Figs. 2 and 3 after Lankester. British Museum Guide Book; Figs. 5 and 6 after
Respiración. — Todos los anfibios, en su juventud, presentan dos o tres pares de branquias externas o internas con hendiduras branquiales externas, excrecencias suaves y plumosas situadas en la parte posterior de la cabeza y ricas en vasos sanguíneos (véase lámina, pág. 407, figs. 1-3). Estas branquias también se encuentran en los peces pulmonados, y en las sirenas y los cachorros de lodo pueden persistir durante toda la vida, aunque esta condición es bastante excepcional. En las salamandras terrestres y en todos los anfibios sin cola (ranas, etc.), las branquias desaparecen y la respiración adulta se realiza completamente mediante extremidades, como en los vertebrados superiores. Mientras las branquias están presentes, las vías respiratorias que conducen el aire a los pulmones a través de la nariz no desembocan en la boca, sino que adoptan esta posición tan pronto como las branquias desaparecen y los pulmones se vuelven funcionales. El aire se aspira por las fosas nasales y, en las ranas, se introduce en los pulmones mediante la deglución. Muchos anfibios pueden vivir mucho tiempo sin alimento, y en las zonas más frías, todas las formas terrestres hibernan durante el invierno en el lodo del fondo de arroyos y estanques, respirando durante este tiempo únicamente a través de la piel.
Audición. — Ningún pez posee oídos funcionales, aunque sí internos; sin embargo, los anfibios poseen órganos auditivos externos diferenciados. Estos se aprecian mejor en las ranas, donde la membrana timpánica del tímpano es un disco circular más o menos grande incrustado en la piel externa a los lados de la cabeza. La cavidad situada detrás del disco, o tímpano, se conecta con la parte posterior de la boca mediante un conducto conocido como trompa de Eustaquio, presente también en todos los vertebrados superiores.
Clasificación. — Las formas vivientes se dividen en dos subclases según su forma corporal: cuando carecen de cola en la etapa adulta, se denominan Anura (sin cola, véase Fig., pág. 406), mientras que las formas primitivas que conservan la cola se denominan Urodela (con cola distintiva, véase Lám., pág. 407, Figs. 1-4). Los anfibios del Paleozoico, que se analizarán más adelante, se incluyen bajo el término Stegocephalia.
Los Anuro incluyen las ranas y sapos tan comúnmente conocidos. Siempre tienen cuatro patas, las traseras son largas y poderosas, y los adultos carecen de branquias externas o hendiduras branquiales. Son casi cosmopolitas, presentes en todos los países excepto en las frías regiones polares. Esta subclase es de origen relativamente moderno, ya que la forma fósil más antigua se conoce a finales del Jurásico, aunque en los estratos cenozoicos [ p. 409 ] se encuentran con mucha mayor frecuencia. Son los anfibios vivos más diversificados, y también los más especializados, pero no son de especial interés para la geología histórica.
Los urodelos o anfibios con cola son de menor importancia en este sentido, y aunque no se conocen formas fósiles anteriores al Jurásico, la subclase debió tener su origen en la Stegocephalia paleozoica. Los urodelos, ahora prácticamente restringidos a las zonas templadas del hemisferio norte, son anfibios que, en su etapa adulta (salamandras, tritones, sirenas, cachorros de lodo y ajolotes), suelen tener una apariencia sorprendentemente parecida a la de los peces (véase la lámina, pág. 407, fig. 1). En los cachorros de lodo y los ajolotes existe una aleta impar en la cara dorsal, que se extiende alrededor de la cola difícercal y a lo largo de la parte posterior de la cara ventral, como en los peces pulmonados. Sin embargo, esta aleta no presenta radios esqueléticos ni óseos, como en los peces. Aunque los Urodelos tienen una apariencia similar a la de los peces, se distinguen fácilmente por el hecho de que sus apéndices pares no son aletas, sino patas completamente desarrolladas, aunque pequeñas y débiles, con todas las características esqueléticas esenciales de las extremidades de los vertebrados superiores (estudio Lám., pág. 407, Figs. 1-4). Por lo general, hay dos pares de estas extremidades: la anterior o pectoral y la posterior o pélvica. Sin embargo, en algunas formas solo está presente el par anterior, habiéndose perdido el posterior, y en unas pocas especies faltan todas las extremidades; estas últimas son las Csecilias excavadoras, similares a gusanos, de las tierras tropicales. El número de dedos en cada pata varía entre cinco, el número habitual, y dos. Algunas formas, como las salamandras terrestres, al alcanzar la madurez pueden vivir completamente en tierra y perder todo rastro de branquias, mientras que otras, como las salamandras de barro, permanecen en el agua y conservan las branquias durante toda su vida. El urodelo gigante japonés y chino es el más grande de todos, alcanzando una longitud de 1,5 metros. Un ejemplar vivió en cautiverio durante más de cincuenta años.
Stegocephafia, los anfibios del Paleozoico. — El término Stegocephalia significa cabeza cubierta o acorazada y hace referencia a que la superficie superior del cráneo estaba cubierta por huesos dérmicos más o menos gruesos. También se les conoce como anfibios de cráneo sólido y acorazado, y es esta armadura la que distingue a las formas del Paleozoico y Triásico de todas las actuales. El pecho también estaba provisto de una armadura dérmica, compuesta por tres grandes placas esculpidas que representaban parte de los huesos del arco pectoral de los peces y la cintura escapular de los vertebrados superiores. En algunas formas, todo el cuerpo estaba cubierto de pequeñas escamas superpuestas, siendo las de la cara ventral las más gruesas, o las escamas solo en la cara ventral, mientras que otras (Branchiosauria) [ p. 410 ] estaban esencialmente desnudas y carecían de escamas. Casi todos los anfibios actuales están desnudos y rara vez tienen escamas (Csecilios). Un grupo de formas antiguas se ha llamado Labyrinthodonts, porque la dentina de los dientes grandes y cónicos estaba muy plegada en ondulaciones laberínticas como las que se vieron por primera vez en los peces (p. 298).
El primer rastro de un vertebrado con extremidades, probablemente similar a un estegocéfalo, es una huella de pie (Thinopus) del Devónico Superior (Fig., pág. 331), pero no se conocen esqueletos de estos anfibios acorazados en Norteamérica hasta el Pensilvánico. Se conocieron entonces y en el Pérmico en su apogeo. Toda la población desapareció antes del final del Triásico.
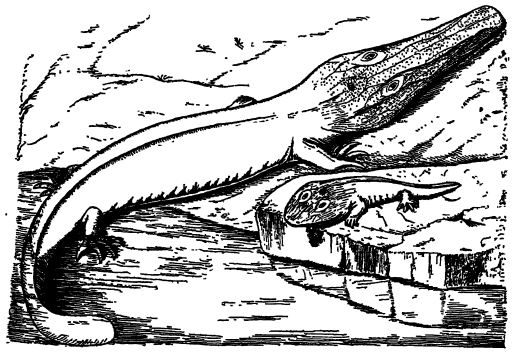
Los anfibios más primitivos tenían cabezas pequeñas y extremidades débiles en cuerpos largos que terminaban en colas propulsoras como las de las salamandras modernas, pero a diferencia de estas, conservaban prácticamente la armadura corporal que heredaron de sus antepasados peces, escamosos y acorazados. Las formas de cabeza grande, observadas principalmente a finales del Pensilvánico y principios del Pérmico, eran descendientes precoces de formas primitivas. La marcada evolución de los antiguos anfibios coincide con su despliegue en todos los diversos entornos, pasando de hábitats acuáticos y pantanosos a hábitats de transición, y [ p. 411 ] finalmente a los de tierras secas, diversificándose aún más al excavar bajo la superficie y ascender a los árboles.
Se conocen muchos tipos de estegocéfalos; tan solo en las mediciones de carbón de Norteamérica se encontraron noventa especies. Su tamaño varía desde aproximadamente una pulgada hasta más de diez pies (Fig., pág. 360). Algunos eran parecidos a lagartos, pequeños y activos, con patas bien desarrolladas para caminar y colas relativamente cortas; otros eran nadadores activos, con colas largas; otros, de tamaño mediano, eran corpulentos y de hábitos lentos, como los cocodrilos (Fig., pág. 410); mientras que algunos tenían la apariencia de renacuajos gigantescos, casi sin patas, que se retorcían en el agua. Invariablemente presentaban cola.
En la gran mayoría de los Stegocephalia existían dos pares de extremidades, aunque también se conocen formas sin patas, como las serpientes (véase Fig., p. 360). Las extremidades estaban bien desarrolladas, pero eran cortas y rechonchas (véase Fig., p. 410, y Lám., p. 407, Figs. 5, 6). Los huesos de las patas delanteras eran similares a los de las salamandras actuales en número, forma y disposición, y las manos contaban con cuatro, o raramente cinco, dedos generalmente cortos. Las extremidades traseras eran casi siempre más largas y pesadas, y presentaban cinco dedos, de los cuales el segundo o el tercero eran los más largos.
Las cabezas solían ser anchas y planas, con bocas muy anchas que abarcaban casi todo el ancho del cráneo, como en ranas y salamandras; pero en algunas formas, las cabezas eran más o menos alargadas e incluso puntiagudas, como en los cocodrilos. Sin embargo, las cabezas de estos últimos también son anchas en las etapas jóvenes, por lo que la cabeza ancha debe ser el tipo primitivo. Del primer tipo, el Eryops del Pérmico de Texas tenía una cabeza de 60 cm de largo y 45 cm de ancho y se asemejaba a un enorme renacuajo con una cabeza ancha y plana, sin cuello, un cuerpo grueso y pesado, patas cortas y una cola pesada y aplanada (véase Branchiosaurus, Fig., p. 410). Como dijo Huxley, «se paseaban con mucha barriga y pocas piernas, como Falstaff en su vejez, entre los bosques de carbón».
Es probable que todos los Stegocephalia vivieran en agua dulce o en tierra firme, y algunos se alimentaban en las playas de los mares del Triásico. Algunos de los tipos pequeños y activos se encuentran en troncos huecos del Pensilvánico de Nueva Escocia. Probablemente la gran mayoría de los Stegocephalia eran carnívoros y se alimentaban de mariscos, gusanos y otros invertebrados acuáticos, pero sobre todo de peces, reptiles y pequeños miembros de su propia tribu.
El tercer ojo de la estegocefalia. — En todos los cráneos estegoefálicos bien conservados, los huesos de la coraza o techo y los del cráneo verdadero estaban perforados no solo por las grandes órbitas laterales donde se ubicaban los ojos pares y por el par de aberturas nasales anteriores, sino también por un único y [ p. 412 ] pequeño orificio a través del hueso sobre el cerebro (véase la fig., pág. 410). Esta abertura es de gran interés, ya que en ella se encontraba un tercer ojo conocido como ojo pineal. Dicha abertura también se encuentra en muchos reptiles fósiles y algunos reptiles actuales (Sphenodon), y si bien el órgano que contiene difícilmente puede considerarse como un ojo verdadero en los reptiles actuales, su gran importancia reside en que se trata de un órgano vestigial cuyo origen se remonta al menos a la época de Pensilvania. Los rudimentos de este ojo están presentes en el cerebro de todos los vertebrados actuales, incluido el hombre.
Ascendencia de los esfegocéfalos. — Cada vez es más difícil distinguir a los reptiles del Paleozoico tardío de sus parientes estegocéfalos. Los reptiles de cabeza sólida más antiguos (Cotylosauria y Pareiasauria) son muy similares a los estegocéfalos de cabeza sólida. Sobre este tipo, Osborn afirma: «Hueso por hueso, sus partes indican una descendencia común del tipo de cráneo de los peces con flecos (Crossopterygia)». En otras palabras, se acumula rápidamente evidencia que demuestra que los estegocéfalos no fueron los vertebrados terrestres con extremidades más primitivos, sino que surgieron de un tronco más antiguo (Protopoda), que dio origen, por un lado, a los anfibios acuáticos y, por otro, a los reptiles, que se adaptaron completamente a la tierra firme.
¶ Reptilia, viva y fósil
En todos los vertebrados estudiados hasta ahora —peces y anfibios— hemos visto que su hábitat es completamente acuático o que, como mínimo, los huevos pequeños se ponen y fecundan allí, y que las crías también nacen y pasan sus días de juventud en este medio. Todos los vertebrados superiores se alejan cada vez más de este hábitat y ninguno se desarrolla en él directamente a partir del huevo. En otras palabras, los reptiles, por regla general, son ovíparos: ponen huevos grandes como los de las aves y reciben una cantidad mayor o menor de alimento (yema). Estos huevos se fecundan antes de ser depositados en tierra, donde eclosionan bajo el calor del sol. Esta es la diferencia más importante y fundamental entre los vertebrados inferiores —peces y anfibios—, por un lado, y los vertebrados superiores —reptiles, aves y mamíferos—, por otro.
Reptiles Vivos. — Todos los animales vivos conocidos como tortugas, lagartos, serpientes, caimanes y cocodrilos, y los tipos extintos, los dinosaurios gigantes del Mesozoico y los reptiles voladores llamados pterodáctilos, pertenecen a la clase Reptilia. La palabra reptil significa reptante o que se arrastra y se refiere a un animal que se mueve boca abajo como la serpiente, o que se mueve con dificultad sobre patas cortas y extendidas, como el caimán. Sin embargo, hay muchos reptiles que no son en ningún sentido animales reptantes, como, por ejemplo, muchos de los lagartos de patas ligeras, algunos dinosaurios con sus patas en forma de pilar, y los pterodáctilos alados.
Todos los reptiles vivos son animales de sangre fría como los peces y los anfibios, y su piel nunca es suave, sino siempre más o menos [ p. 413 ] [ p. 414 ] endurecida por material córneo u óseo que se presenta con más frecuencia como escamas que como placa de armadura. Aunque muchos reptiles pasan gran parte del tiempo en el agua, son esencialmente animales terrestres, ya que nacen en la tierra y respiran exclusivamente a través de pulmones. Cada animal tiene un par de pulmones, pero en las serpientes alargadas el pulmón izquierdo es rudimentario y casi perdido. En las serpientes y en algunos lagartos, las patas están totalmente ausentes o son meros vestigios enterrados en la carne, y en tales reptiles la locomoción se realiza mediante un movimiento de serpenteo ya sea sobre el suelo o a través del agua. Dondequiera que estén presentes las patas, los dedos de los pies tienen garras, una característica que es muy rara entre los anfibios.
Muchos reptiles presentan párpados, y en la mayoría de los lagartos y reptiles fósiles también se encuentra la glándula pineal. Los órganos del olfato y el oído también están bien desarrollados, pero en los reptiles la membrana vibratoria, o membrana timpánica, del tímpano ya no es externa, como en las ranas, sino que se encuentra en una depresión. Con excepción de las tortugas, casi todos los reptiles tienen dientes; en las formas vivas, estos suelen ser puntiagudos y a menudo retraídos para sujetar a sus presas. Sin embargo, en muchas formas fósiles, los dientes estaban adaptados para cortar y, más raramente, para masticar alimentos. En las serpientes y algunos lagartos, la lengua es delgada y bifurcada, y sobresale de forma rápida. En otros reptiles, la lengua es plana e inmóvil, como en los cocodrilos, y se adhiere al suelo de la boca.
Desarrollo de los Reptiles. — En algunos lagartos y serpientes actuales, y raramente entre las formas extintas, las hembras son vivíparas, es decir, dan a luz crías vivas y completamente formadas. Sin embargo, la gran mayoría de los reptiles son ovíparos. Los huevos se parecen a los de las aves, pero suelen ser más redondos y tener una cubierta dura, porosa y apergaminada, o en algunos casos, una cáscara calcárea. Dado que estos huevos difieren mucho en su desarrollo de los de los peces y anfibios, es necesario describirlos con más detalle.
Dado que los huevos de los reptiles son grandes y se desarrollan en tierra firme, su estructura interna debe ser muy diferente a la de los pequeños huevos de los anfibios que eclosionan en el agua. En estos últimos, los embriones poseen branquias funcionales para su uso en el agua, pero en los reptiles se ha originado un órgano completamente diferente para proporcionar a las crías en desarrollo dentro del huevo el oxígeno necesario. Este órgano embrionario, con forma de saco, conocido como alantoides, transmite al embrión el oxígeno recibido a través de la cáscara porosa y, por el proceso inverso, elimina el gas carbónico; en otras palabras, su función es respiratoria (véanse las figuras AE, pág. 415).
[ p. 415 ]
El reptil no nacido respira mediante una capucha vascular que se extiende bajo la cáscara del huevo y absorbe el aire seco del exterior. Es interesante que esta capucha vascular, llamada alantoides, esté representada en los anfibios por una vejiga sin importancia que crece desde el extremo posterior del canal alimentario. Un gran paso en la evolución estuvo implícito en el origen de esta capucha prenatal o membrana fetal y otra —de importancia protectora— llamada amnios, que forma una bolsa de agua sobre el delicado embrión. Este paso significó la emancipación total del agua y de la respiración branquial, y las dos membranas fetales, el amnios y la alantoides, persisten no solo en todos los reptiles, sino también en aves y mamíferos. (J. A. Thomson en Outlines of Science).
Los huevos de reptil son grandes y contienen una gran cantidad de vitelo, en el que vive y crece el embrión. En un polo del huevo se encuentra la vesícula germinal fertilizada que se desarrolla en el embrión (Fig. A, lado opuesto). Durante su crecimiento más temprano se forma en cada extremo del embrión alargado un pliegue de dos capas, en forma de media luna, llamado pliegue amniótico, que se arquea sobre el embrión y finalmente se une para cubrirlo con una capucha protectora [ p. 416 ]. Este último es el amnios, y entre él y el embrión hay una cavidad poco profunda que contiene un líquido amniótico acuoso, que baña y protege la superficie exterior del embrión (Fig. B, pág. 415). Como el amnios se produce solo en los huevos de reptiles, aves y mamíferos, estos grupos se conocen colectivamente como Amniota, y los peces y anfibios, que carecen de él, se llaman Anamnia, que significa sin amnios.
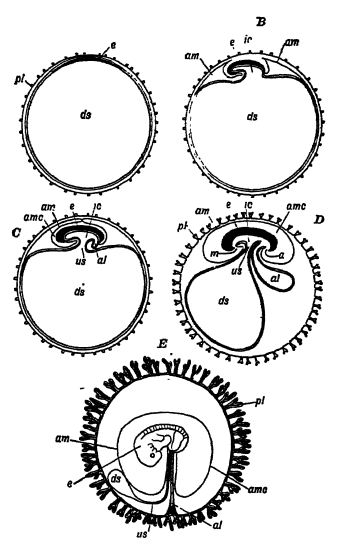
a. gnus; at, allantois; am, amnion; amc, amniotic cavity, ds, cavity of embryonic vesicle, later the yolk sac; e, embryo; e, intestinal cavity; m, mouth; placenta, an organ found only in mammals, formed Of walls of the uterus and embryonal membrane; both are abundantly supplied with blood-vesels, and while there is no direct continuity in the two blood streams, nutrient materials and respiratory gase are exchanged by osmoäs; u, umbilical stalk. After Köniker.
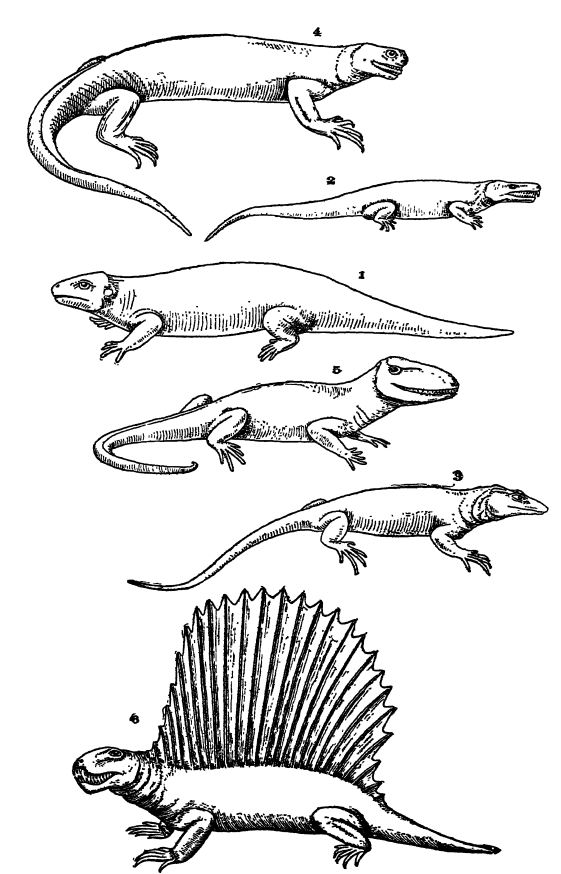
Reptilia Paleozoica. — La mayoría de los reptiles del Pensilvánico y del Pérmico temprano eran animales regordetes, perezosos y más o menos despatarrado, que a menudo se asoleaban en la tierra bajo el sol abrasador. En muchos aspectos, aún se parecían a los estegocéfalos, sus parientes, pero presentaban una marcada tendencia a la reducción del tamaño del cráneo y a la pérdida de la armadura corporal. En todas las formas, los pies terminaban en cinco dedos. Pocos parecen haber sido ágiles de pies, y algunos, los «lagartos de barco» o «lagartos de aleta dorsal», presentaban una curiosa cresta dorsal media muy alta (Pelycosauria, Pl., p. 413, Fig. 6). «Estas estructuras», afirma Osborn, «pueden haberse desarrollado mediante la competencia y selección social o racial dentro de esta familia de reptiles, más que como órganos ofensivos o defensivos en relación con otras familias de reptiles». La mayoría de las formas del Paleozoico tardío eran animales de aspecto feroz debido a sus grandes dientes curvados y de agarre. Todos los representantes americanos parecen haber sido carnívoros, alimentándose de insectos, peces y anfibios acorazados, y otros reptiles. Otros tenían dientes trituradores, lo que indica una dieta de mariscos y crustáceos.
Osborn afirma que el entorno que transformó a los primitivos anfibios del Paleozoico tardío en los reptiles del Pérmico fue una región cálida, terrestre y semiárida, propicia para el desarrollo de un sistema nervioso sensible, movimientos ágiles, armazón escamoso, extremidades delgadas, una cola vibrante y la captura de alimento tanto mediante dientes curvados y puntiagudos como con las garras de una mano y un pie de cinco dedos. Esta evolución es tan maravillosa y extrema como la posterior de los mamíferos. Antes del final del Pérmico, los reptiles dominaban todas las partes de la tierra y se adueñaban también de las aguas de la tierra y los mares.
Parece probable que los reptiles surgieran incluso antes del Pensilvánico más temprano, pues en la última parte de este período aparecieron no solo reptiles verdaderos, sino también formas altamente especializadas. Del Pensilvánico y del Pérmico más antiguo de Texas, Oklahoma y Nuevo México, Williston y Case nos han dado a conocer muchos tipos diferentes de anfibios primitivos, y asociada a ellos existe una sociedad aún mayor y más [ p. 417 ] compleja de reptiles primitivos, animales que alcanzaron una longitud de 2,4 metros (véase lámina, pág. 413).
El orden Cotylosauria abarca a los reptiles de cráneo sólido, llamados así porque la cabeza estaba recubierta de placas esculpidas (Lám., pág. 413, Figs. 1, 2). Eran los reptiles más primitivos. De complexión baja y robusta, de movimientos lentos y torpes al caminar, pero gracias a sus colas bastante largas, nadaban bien. Todos eran carnívoros, aunque algunos se alimentaban parcialmente de plantas. Su área de distribución se extendió desde finales del Pensilvania hasta finales del Triásico, y se conocen principalmente en Norteamérica y Europa, aunque algunos se encuentran en Sudáfrica.
El orden Theromorpha (que significa bestia salvaje), que incluye a los lagartos de aleta dorsal Pelycosauria, un grupo altamente especializado de formas terrestres. Eran activos y similares a los lagartos, con colas bastante largas, y algunos alcanzaban una longitud de 2,4 metros. Los lagartos de aleta dorsal eran regordetes y presentaban una cresta o aleta dorsal ósea más o menos alta. Todos eran ferozmente carnívoros. Su área de distribución se extendió desde finales del Penns III hasta finales del Pérmico, y su mayor abundancia se encontraba en Norteamérica, siendo menos común en Europa y África.
Sudáfrica es otra región donde se conservan numerosos esqueletos excelentes de antiguos anfibios y reptiles. Se encuentran comúnmente en la extensa serie Karoo (hasta 2900 metros) de lutitas y areniscas continentales, y en épocas Pérmicas y Triásicas. Según Osborn, la mayor cantidad de formas se encuentra en el Pérmico temprano y medio, cuando el mundo ya contaba con una maravillosa sociedad de reptiles.
Este conjunto de reptiles africanos incluía grandes formas herbívoras de cuerpo redondo (pareiasaurios), con extremidades enormes y cabezas redondas y sólidas; ágiles, con grandes cuencas oculares y adaptados para correr velozmente (dromosaurios = reptiles sin dientes), «reptiles terribles» con cabezas parecidas a las de los mamíferos (Theriodontia = dientes carnívoros) y muchos animales depredadores de extremidades delgadas con afilados colmillos. También existían reptiles carnívoros gigantes (dinocéfalos = «cabeza terrible»), animales muy grandes con lomo muy arqueado, frente ancha y abultada, y mandíbulas cortas y [ p. 418 ] anchas provistas de dientes marginales. Superando incluso a estos en tamaño estaban los «reptiles dentados sin ley» (anomodontos), en los que el cráneo medía entre un par de pulgadas y un metro de largo, y las mandíbulas estaban revestidas de cuerno y ganchudas como las de las tortugas. Finalmente, a principios del Triásico aparecieron los reptiles «con dientes de perro» (cinodontos), los más parecidos a los mamíferos de todos.
Progenitores de los reptiles superiores. — Como se ha dicho, los reptiles más primitivos son los cotilosaurianos. Lull nos indica que este orden, como se observa en Limnoscelis (Fig. 2, p. 413), bien pudo haber sido el tronco central del cual evolucionaron, directa o indirectamente, los lagartos, caimanes y dinosaurios. Los cotilosaurianos eran reptiles que vivían en pantanos, perezosos, de cola larga y cuatro patas de proporciones iguales.
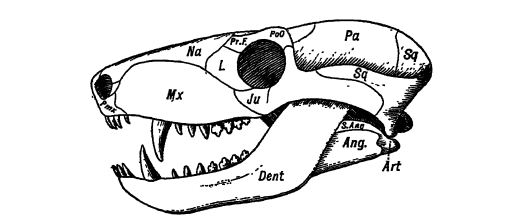
Progenitores de los mamíferos. — En el Pérmico y el Triásico de África y Norteamérica se encuentran reptiles carnívoros del suborden Theriodontia. Su tamaño varía desde formas pequeñas hasta las más grandes que un tigre. Son de gran interés porque se considera que las formas africanas dieron origen a los mamíferos inferiores o ovíparos, mientras que las formas americanas dieron origen a los reptiles superiores. Sus dientes estaban diferenciados y localizados, al igual que en los mamíferos, en incisivos, caninos y molares (véase la fig., pág. 417).
¶ Lectura colateral
Caso EC, Los lechos rojos del Pérmico-Carbonífero de Norteamérica y su fauna de vertebrados. Institución Carnegie de Washington, Publicación n.° 207, 1915.
Caso CE, El entorno de la vida en el Paleozoico Tardío en América del Norte: un estudio paleogeográfico. Ibíd., Publicación n.° 283, 1919.
R. L. Moodie, The Coal Measures Amphibia of North America. Ibíd., Publicación n.° 238, 1916.
H. F. Osborn, El origen y la evolución de la vida, págs. 177-233. Nueva York (Scribner), 1917.
S. W. Williston, Vertebrados del Pérmico Americano. Chicago (University of Chicago Press), 1911.
S. W. Williston, Reptiles acuáticos del pasado y del presente. Chicago (University of Chicago Press), 1914.
S. W. Williston, La filogenia y clasificación de los reptiles. Revista de Geología, vol. 25, 1917, págs. 411-421.
| XXIX. El carbón y su presencia en la naturaleza | Página de título | XXXI. El Pérmico y su clima glacial |