[ pág. 186 ]
FIN DEL TERCER INTERGLACIAL. CLIMA TEMPLADO Y ÁRIDO. INDUSTRIA ACHEULÉE — ADVENIMIENTO DE LA CUARTA GLACIACIÓN, PROFUNDOS CAMBIOS EN LA VIDA ANIMAL Y VEGETAL — EL PERÍODO DE LA TUNDRA ÁRTICA EN LA VIDA DE MAMÍFEROS Y VEGETALES — CARACTERÍSTICAS DE LA RAZA NEANDERTAL, DE SU INDUSTRIA DEL SÍLICE MUSTERIANO — SUPUESTAS CAUSAS DE EXTINCIÓN O DISPERSIÓN
Llegamos ahora a una etapa prolongada e importante de la prehistoria de Europa, a saber, el período de la cuarta glaciación, del desarrollo final de la raza humana neandertal, de la industria musteriense, de los comienzos de la vida en cuevas, de la caza del reno y su utilización como alimento y vestido.
En toda Europa, la industria achelense parece haber llegado a su fin durante un período de clima árido, cálido en algunas partes de Europa occidental y fresco o incluso frío en otras. Las variaciones estacionales pudieron haber sido extremas, como en las estepas del sur de Rusia, donde veranos extremadamente calurosos pueden dar paso a inviernos intensamente fríos, con fuertes vientos y tormentas de nieve destructivas para la vida.
Es esta alternancia estacional, así como la recurrencia, ya sea estacional o secular, de un clima más cálido, lo que explica la supervivencia o el regreso de la fauna asiática incluso después del cierre de la industria achelense y cuando la industria musteriense estaba muy avanzada.
A partir de los yacimientos hallados en Grimaldi, en la Gruta de los Niños y en la Gruta del Príncipe, se ha afirmado durante mucho tiempo que los hombres del Musteriense temprano vivieron simultáneamente con el hipopótamo, el elefante de colmillos rectos y el rinoceronte de Merck en el clima benigno de la Riviera Mediterránea. Más recientemente, se han encontrado los mismos animales tan al norte como el valle del Somme en las corrientes fluviales de Montières-les-Amiens.1 Aquí, de nuevo, encontramos restos de 186 [ p. 187 ] del hipopótamo, el elefante de colmillos rectos y su compañero, el rinoceronte de Merck, en yacimientos del Musteriense, un descubrimiento sorprendente, ya que siempre se había supuesto que un período climático frío se había establecido en toda Europa occidental incluso antes del fin de la cultura achelense. Pero también hay evidencia de un clima templado que todavía prevalecía en el valle del Támesis en el período de los ‘pisos’ Musterienses. 2 De nuevo, a lo largo del valle de Vézère, Dordoña, encontramos que en la estación de La Micoque, donde la industria marca la transición entre el Achelense tardío y el Musteriense temprano, el rinoceronte de Merck se encuentra en las capas más bajas asociado a restos del alce (Alces).
Existe evidencia de que el rinoceronte de Merck y el elefante de colmillos rectos persistieron en Europa occidental durante todo el período del desarrollo temprano de la industria musteriense. Como se mencionó anteriormente, estos animales eran más resistentes que el mamut del sur, que fue el primero de los mamíferos asiáticos en desaparecer, seguido pronto por su compañero, el hipopótamo. Incluso después de la aparición de la pareja de la tundra estrechamente asociada, el mamut lanudo y el rinoceronte lanudo, el rinoceronte de Merck persiste, como, por ejemplo, en los yacimientos de Rixdorf, cerca de Berlín, donde este antiguo tipo se encuentra en los mismos yacimientos que el mamut lanudo, el rinoceronte lanudo, el reno y el buey almizclero, así como con las formas forestales, el alce, el ciervo, el lobo y el caballo de bosque. La extrema latitud septentrional de este yacimiento explica la ausencia del elefante de colmillos rectos, que en aquella época podría haber habitado más al sur. La misma mezcla de mamíferos del sur y el norte de Asia se encuentra en Steinheim, en el valle del río Murr, algunos grados al oeste y al sur de Rixdorf, no lejos de Gotinga, donde encontramos el rinoceronte de Merck® y el elefante de colmillos rectos en asociación con el mamut lanudo, el rinoceronte lanudo, el ciervo gigante y el reno.
Así, las razas neandertales entraban en la etapa cultural musteriense durante el final de la Tercera Etapa Interglaciar y durante el período inicial del avance de los campos de hielo desde los grandes centros de Escandinavia y los Alpes. A medida que estos campos de hielo se acercaban lentamente desde el norte y el sur, debió transcurrir un largo período de tiempo durante el cual todos los mamíferos del sur de Asia abandonaron Europa occidental o se extinguieron, con la excepción de los leones y las hienas, quienes se adaptaron bien al clima extremadamente severo que prevaleció en Europa durante la cuarta glaciación, e incluso durante la larga Etapa Postglaciar que le siguió. Los grandes carnívoros se adaptaron fácilmente a los climas fríos, ya que subsisten de la vida animal dondequiera que se encuentre. Se han encontrado tigres de la misma estirpe que los de la India tan al norte como el río Lena, a 52° 25’ de latitud, donde el clima es más frío que el de Petrogrado o Estocolmo, mientras que el león prosperó en la fría atmósfera de la cordillera del Alto Atlas. Así, el león cavernario (Felis leo spelaea) y la hiena cavernaria (H. crocuta spelaea) desarrollaron sin duda una capa interna de pelaje, así como una capa externa de pelo largo, como los mamíferos de la tundra. En tamaño, el león de este período en Francia a menudo igualaba y, en ocasiones, superaba a sus parientes actuales, el león africano y el león de Asia occidental; figura con frecuencia en el arte de los artistas del Paleolítico Superior y sobrevivió en Europa occidental hasta el final de dicho período.
¶ La Cuarta Glaciación
Penck4 ha estimado que el primer máximo de la cuarta glaciación en los Alpes se alcanzó hace 40.000 años, y que, tras el período de recesión, el segundo máximo finalizó hace no menos de 20.000 años. Esto extendería la industria musteriense durante un período muy largo, pues no cabe duda de que la cultura musteriense fue prácticamente contemporánea a la cuarta glaciación, incluso si se le atribuyera un período más breve a este gran evento natural.
Se cree que la cuarta glaciación, al igual que la primera, fue contemporánea en Europa y Norteamérica, 5 hecho de especial importancia para los antropólogos estadounidenses en relación con la fecha de llegada del hombre primitivo a América. En ambos países, la glaciación alcanzó un máximo temprano, seguido de un período de retroceso de los campos de hielo, durante el cual prevaleció un clima algo más templado, pero que a su vez dio paso a un segundo avance de la misma severidad que el primero.[1]
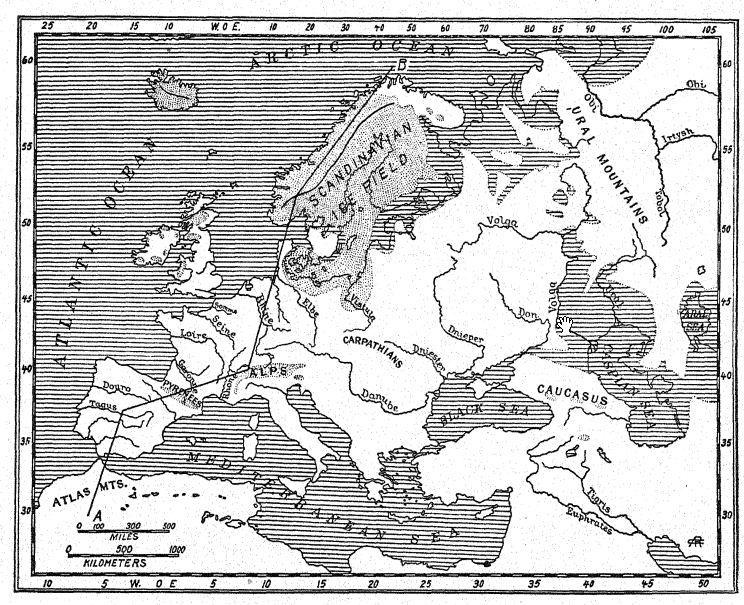
En el norte, Escandinavia y Finlandia volvieron a quedar cubiertas de hielo, y un gran mer de glace ocupó la cuenca del mar Báltico, enviando sus morrenas terminales hacia Dinamarca y Schleswig-Holstein [ p. 190 ] y sobre las provincias septentrionales de Alemania, pero este gran campo de hielo no volvió a confluir con el de Gran Bretaña.7 Al comienzo de la cuarta glaciación, grandes glaciares descendieron sobre los valles montañosos escoceses y llenaron muchos de ellos incluso hasta el mar; la costa se hundió al menos 40 metros en esta región. En el sur de Gran Bretaña, a lo largo del valle del Támesis, se extendió una flora ártica, con el sauce polar (Salix polaris) y el abedul enano (Betula glandulosa); una planta ártica [ p. 191 ] También se ha descubierto un lecho de tundra en el valle del Lea. Así, el clima de tundra se extendió desde las tierras bajas escocesas hasta el sur de Inglaterra, siendo el terreno árido y casi sin árboles.8 Creemos que este fue también el período de la flora ártica en Hoxne, Suffolk, y del lecho vegetal ártico en el valle del Támesis. En esa época, el valle era frecuentado por el reno, el rinoceronte lanudo y el mamut, cuyos restos se encuentran enterrados en los aluviones de bajo nivel arrastrados por las laderas del valle, de modo que los restos de esta fauna ártica podrían, en algunos lugares, sobrepasar los de la fauna asiática cálida, mucho más antigua y enterrada a mayor profundidad, de la época chelense. Al igual que el Somme, el Támesis® se encontraba entonces entre 10 y 25 pies por debajo de su nivel actual, habiéndose sedimentado su fondo con tierra aluvial.

Este fue el período de la sedimentación de la «deriva superior» sobre las tierras bajas del norte de Alemania, los Alpes y el norte de Inglaterra, así como de la formación Wisconsin temprana y tardía, también conocida como «deriva superior», que se extiende ampliamente por los estados del este, desde Wisconsin hacia el sur y el este hasta la latitud de Nueva York. También se depositaron las gravas y arenas de algunas de las «terrazas inferiores».
¶ La vida de los mamíferos en el Musteriense
Las tres fases sucesivas del clima y del entorno que rodearon al hombre de Neandertal durante el período de desarrollo de la industria musteriense fueron, en orden descendente, las siguientes:
3. Clima extremadamente frío del Último Gran Avance Glacial. Periodo de la industria del Musteriense Tardío de La Quina. Expansión de todos los mamíferos árticos y de la tundra por Europa occidental, incluyendo el niusk-ox; migraciones del obi y el lemming anillado del extremo norte. Vida e industria de las razas neandertales, principalmente en los refugios, grutas y entradas a las cavernas.
2. Clima frío y húmedo. Periodo de la industria del Musteriense medio o pleno de las razas neandertales. Aparición de la vida de la tundra, incluyendo mamíferos y aves bien protegidos de la región ártica, así como la ascendencia de los tipos alpinos a las estribaciones y riberas fluviales. Primeros precursores de la vida esteparia; la vida eólica eurasiática plena, tanto forestal como campestre, se extendió ampliamente por Europa. La vida y la industria se concentraban principalmente en los refugios, grutas y entradas a las cavernas. Los renos eran muy abundantes.
1. Clima cálido o frío árido. Transición del Achelense a la cultura Musteriense temprana, como se observa en las estaciones de La Micoque y de Combe-Capelle. La fauna denominada «Musteriense cálido», que incluye el hipopótamo superviviente, el rinoceronte de Merck y el elefante de colmillos rectos en el norte y sur de Francia; manadas de bisontes, ganado vacuno y caballos salvajes en el suroeste de Francia. Vida tribal, con la industria en parte en estaciones abiertas y en parte bajo acantilados resguardados.
Este es el comienzo del «Período del Reno», pues este migrante procedente de Escandinavia, junto con sus compañeros de las tundras nororientales, el mamut lanudo y el rinoceronte, se desplazó lentamente hacia el sur ante el avance de los campos de hielo escandinavos, que se vieron enormemente aumentados por el clima cada vez más frío y húmedo. Así, estos animales se encuentran en el norte con pedernales de la cultura musteriense antes de aparecer en la región más benigna de Dordoña. En la estación musteriense achelense de La Micoque, algo más antigua, a orillas del Völztre, los fogones contienen casi exclusivamente restos de caballos y relativamente pocos restos de bisontes y ganado salvaje, pero ningún reno. Una chimenea cerca de la estación de Combe-Capelle contiene numerosos restos de bisonte, solo algunos de caballo y los primeros de reno. Antes de la aparición del reno en el valle del Vézère, podemos imaginar las praderas cubiertas de bisontes y caballos salvajes, estos últimos del tipo que ahora es característico de las altas mesetas del Asia central, mientras que el bisonte de la época parece ser más similar al búfalo americano que a la forma europea sobreviviente.
Gradualmente, los animales de la tundra se extendieron hacia el sur con el clima frío que, por primera vez, azotó toda Europa occidental. El aspecto del país cambió lentamente con la llegada del reno, y la flora septentrional, compuesta por píceas, abetos y sauces árticos, cubrió los valles fluviales y las laderas más resguardadas, mientras que las mesetas y los campos quedaron deforestados parcial o totalmente.
Así, el país se adaptó principalmente a los mamíferos de la tundra; y en los estratos del Musteriense medio, estas manadas, recién migradas desde el extremo norte y desde las estepas nororientales que bordean el río Obi, superan ampliamente en número a las formas esteparias, que se limitan a dos o tres especies. De estas, las principales son el caballo estepario, emparentado con el caballo de Przewalski que ahora habita en el desierto de Gobi, el suslik estepario (Spermophilus rupescens) y el urogallo estepario o gallineta común. Las formas más características de la vida esteparia, como el antílope saiga, el jerbo y el kiang, llegaron más tarde y no aparecieron hasta después del cierre de la industria musteriense y la desaparición de la raza neandertal.

Esto se debió al hecho de que el clima que rodeaba a la raza neandertal en tiempos musterienses era frío y húmedo, con fuertes lluvias en verano y tormentas de nieve en invierno, un clima totalmente adecuado para los mamíferos árticos timdra con su espesa cubierta de pelo que actuaba como protector contra la lluvia y la capa interna de lana que los protegía del clima más severo.
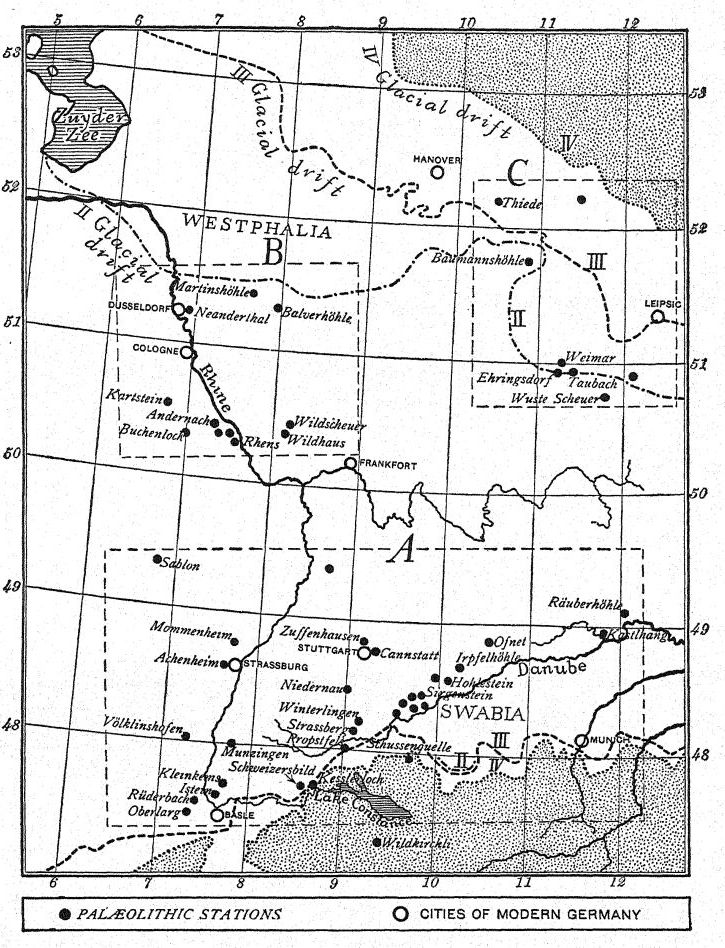
La vida de los mamíferos durante la cuarta glaciación, a medida que se extendía hacia la región del Rin medio y Westfalia, está plenamente documentada en los yacimientos de Toess de Achenheim y en la famosa gruta de Sirgenstein, en el alto Danubio, al noroeste de Múnich. Allí, junto con vestigios de la industria musteriense más primitiva, se encuentran restos del mamut, el bisonte, el reno, una especie de caballo salvaje y el oso cavernario. Tras estos mamíferos, en el mismo yacimiento se registra la llegada del lemming de Obi, procedente del norte de Rusia.
El hecho de que solo se conozcan siete estaciones musterienses en toda Alemania, u ocho si incluimos el yacimiento del entierro neandertal, puede explicarse por la relativa proximidad del gran glaciar escandinavo al norte, que se encontraba a tan solo 560 kilómetros del gran glaciar alpino al sur. Al este se encontraban las llanuras de Bohemia y la vasta región de tierras bajas que se extendía al noreste hasta las tundras y al este hasta las estepas, por donde llegaron las grandes migraciones de la vida de la tundra y la estepa.
[ pág. 195 ]
[ pág. 196 ]
¶ Entorno geográfico y climático de la raza neandertal
Echemos un primer vistazo a Dordoña. Entre las estaciones de la industria musteriense temprana, hemos visto que los neandertales del valle del Vézère, en La Micoque, se encontraban en medio de una fauna compuesta principalmente por bisontes y caballos salvajes, siendo los restos encontrados en los hogares casi exclusivamente de este último animal.[2] En la primitiva estación musteriense de CombeCapelle, cerca de allí, los hogares de los fogones contienen restos de bisonte, pero solo algunos de caballo.
Entre las primeras cuevas habitadas por el hombre10 estaba la de Le Moustier, situada en la orilla derecha del Vézère, y a unos 90 pies por encima de él. Este refugio y cueva fueron examinados ya en 1860-3 por Lartet11 y Christy y posteriormente por de Vibraye,12 Massenat,13 y otros. Además de los depósitos en el suelo de la gruta, se ha encontrado una profunda capa de cultura musteriense bajo el acantilado de enfrente, y esta ha sido seleccionada para nuestra representación de la vida de los hombres de la época musteriense y de la flora del Vézère en este período temprano (véase el frontispicio). Peyrony observa que, aquí como en otros lugares, los campamentos industriales más antiguos y bajos estaban más alejados de los refugios; de hecho, en esta misma región hay evidencias de que los trabajadores del sílex chelense y achelense visitaban ocasionalmente las mesetas superiores; Pero con el paso del tiempo y el clima más severo, los neandertales comenzaron a trabajar más cerca de los acantilados que sobresalían y, finalmente, directamente debajo de ellos. En esta estación clásica de Le Moustier, Hauser desenterró en 1908 uno de los esqueletos más completos del hombre de Neandertal. Aquí residieron de forma continua durante el Musteriense medio y superior, extendiéndose hasta el Auriñaciense inferior del Paleolítico Superior. La fauna contemporánea en estos depósitos incluía el mamut, el reno, el ciervo gigante (Megaceros), el caballo, el bisonte, el rinoceronte lanudo y el oso de las cavernas. Durante la ocupación humana de esta estación típica, el clima era muy frío y húmedo.
En esta región se encuentra el registro completo del curso de la evolución musteriense, tanto en las herramientas como en la aparición de nuevas formas de vida; el número de renos aumenta gradualmente en los estratos ascendentes con el desarrollo de la industria musteriense. Existe una gradación constante desde los tipos industriales acheiles hasta los musterienses; según Gartailhac, esta industria es obra del mismo pueblo, sin líneas divisorias definidas.

Así, en Combe-Capelle, donde se originó la verdadera cultura musteriense, encontramos varios grandes coups de poing que nos retrotraen a los primeros aperos achelenses. Las gradaciones que se muestran aquí en estas capas sucesivas contrastan con el avance de la industria al final del Musteriense en la misma localidad, donde se produce una abrupta transición cultural hacia el Auriñaciense.
El sur de Gran Bretaña describe una secuencia similar, que podemos interpretar de la siguiente manera. Las estaciones musterienses a lo largo del río Lea y cerca de la desembocadura del Támesis en Crayford (Worthington Smith,14 Geikie15), pertenecientes al clima templado [ p. 198 ] del Musteriense temprano, o al período de recesión de la cuarta glaciación, conocido en los Alpes como Laufenschwankung, son las estaciones musterienses a lo largo del río Lea y cerca de la desembocadura del Támesis en Crayford (Worthington Smith,14 Geikie15). Estos suelos paleolíticos del Musteriense están enterrados bajo entre 1,2 y 1,5 metros de arena y marga, y descansan sobre la superficie de gravas fluviales más antiguas. Entre los depósitos fluviales posteriores se han descubierto varias superficies terrestres antiguas; consisten en unos pocos centímetros de grava angular, amontonada en algunos lugares con herramientas y lascas sin desgastar que obviamente se encuentran justo donde fueron dejadas por los trabajadores pakolíticos. En cierto punto, hay evidencia de que el artesano se agachaba sobre su trabajo, con las rodillas ligeramente separadas, pues las astillas se lanzaban a derecha e izquierda en pequeños montones. Aquí y allá, hechas con estos utensilios musterienses, se encuentran formas más arcaicas que podrían haber sido arrastradas desde las superficies terrestres más antiguas.

[ pág. 199 ]
Un piso de este tipo ha sido rastreado por Worthington Smith16 a través de Middlesex y en ambas orillas del Támesis. Los restos de plantas ocurren abundantemente en esta antigua superficie terrestre, incluyendo impresiones de porciones de hojas, tallos de hierba, juncos y juncias. El abedul, aliso, pino, tejo, olmo y avellano han sido reconocidos. El helecho macho común es de ocurrencia frecuente, mientras que el helecho real (Osmunda regalis) se encuentra en profusión. En general, este conjunto de plantas indica un clima templado. Los pedernales descritos y figurados por Worthington Smith son o bien del tipo ‘lasca Levallois’ del Achelense tardío o bien de la edad Musteriense temprana. Este escritor señala la gran cantidad de instrumentos conocidos como lascas recortadas, que se forman en el ‘suelo’ Paleolítico; estas son lascas de gran tamaño, recortadas en forma de herramienta en un lado, mientras que el otro lado se deja perfectamente liso; Los ejemplos se mantienen notablemente constantes en una forma. El tipo de herramienta aquí descrito se asemeja a las lascas de Levallois o Combe-Capelle, o incluso a la típica punta de Le Moustier. Estas lascas, moldeadas en las formas musterienses del racloir, o raspador, son muy comunes en las gravas del Lea y del Támesis.
Aunque aquí se encuentran los restos del mamut lanudo, también hay indicios de la presencia de una flora templada bien marcada. Estas “derivas fluviales” de alto nivel a lo largo del Támesis18 se depositaron ciertamente cuando las condiciones climáticas eran templadas, pero son sucedidas por depósitos que indican un nuevo período frío, que puede representar los tiempos fríos del “Musteriense pleno” de la habitación del Paleolítico Inferior del Támesis. Aquí encontramos las notables láminas de “deriva” retorcidas atribuibles a los movimientos del suelo y el subsuelo congelados al exponerse al calor del sol de verano. Al mismo tiempo, es posible que se hayan depositado a lo largo del Támesis las margas y gravas aluviales, que en ocasiones contienen piedras y rocas, que fueron arrastradas por balsas de hielo; estas gravas de bajo nivel no deben confundirse con las “gravas fluviales antiguas” subyacentes que contienen la fauna de hipopótamos de temperaturas cálidas, ya que se acumularon en condiciones muy frías. Se encuentran restos del rinoceronte lanudo y del mamut. Así, tanto en las zonas altas del Támesis como en las bajas encontramos evidencias de la cultura humana y de la fauna extinta del período de la cuarta gran glaciación.
Las aguas superiores del Rin y del Danubio también fueron frecuentadas por trabajadores del sílex del Achelense tardío y del Musteriense temprano. En un punto muy alejado del sur de Inglaterra se encuentra la caverna de Wildkirchli en los montes Santis, cerca de Appenzell, en Suiza; en la época Musteriense, esta se encontraba en pleno corazón del campo de hielo alpino norte. La fauna presente en este lugar podría indicar que esta caverna estuvo abierta durante el período de recesión entre los dos grandes avances de la cuarta glaciación. Aquí, a una altitud de 1377 metros, Bachler (1903-1906) descubrió pruebas de ocupación por el hombre de Neandertal durante la época Musteriense; los pedernales no están bien formados; la presencia de herramientas de hueso toscas podría apuntar a la época Musteriense tardía; sin embargo, Bachler considera que los pedernales pertenecen a la misma etapa que los de Le Moustier. Se afirma que cuando los neandertales perseguían la caza aquí, el clima era más benigno, ya que entre los animales encontrados se incluyen el ciervo, el lobo alpino (Cyon alpinm fossilis), el oso cavernario, el león cavernario, el leopardo cavernario (Felis pardus spelaea), el tejón, la marta y la nutria, junto con las formas típicas alpinas: la cabra montés, el rebeco y la marmota. Sin embargo, esta fauna por sí sola difícilmente puede considerarse prueba de un clima templado, ya que a esta altitud alpina no cabría esperar descubrir la vida de la tundra de la época; de hecho, está completamente ausente.


De todas las estaciones a lo largo del Danubio, la más importante es, con diferencia, la de Sirgenstein, situada entre las ciudades modernas de Núremberg y Augsburgo. Esta estación fue ocupada inicialmente por los neandertales a principios del Musteriense y continuó siendo visitada por los hombres del Paleolítico Inferior y Superior hasta el final del Paleolítico Superior. La continua sección de vida animal y de cultura humana que esta notable caverna revela proporcionó a Schmidt, quien comenzó sus investigaciones aquí en la primavera de 1906, una clave para la prehistoria de las dieciocho cavernas en la región del Alto Danubio y el Alto Rin. En Sirgenstein se encontró la primitiva cultura del Musteriense de los primeros neandertales, junto con restos de mamut, bisonte, reno, una especie de caballo salvaje y el oso de las cavernas. Esta industria musteriense cerró con un registro de la llegada a esta región del lemming Obi desde el norte de Rusia. Más tarde, la raza cromañona de la época auriñaciense dejó en el suelo de la caverna restos de su industria del sílex y de sus festines, incluyendo huesos de rinoceronte lanudo, mamut, ciervo y reno. Durante el Paleolítico Superior Solutrense la caverna no estaba ocupada; pero a principios del Magdaleniense volvió a ser habitada por el hombre, y coincidiendo con su regreso está la llegada de una gran migración del lemming bandeado (Myodes torquatus) desde las tundras árticas del norte. Finalmente, hacia el final del Paleolítico Superior, a finales del Magdaleniense, otra transición climática está indicada por la aparición de la pika, o liebre sin cola (Lagomys pusillus). Durante la Edad del Bronce, se volvió a acceder a esta gruta predilecta, y también estuvo habitada durante parte de la Edad del Hierro. Los restos de estas diversas culturas, hogares y depósitos de marga cavernaria alcanzan un espesor total de 2,5 metros y sitúan a Sirgenstein como la primera estación paleolítica de Alemania.
¶ Tipos y migraciones de los mamíferos cazados por los neandertales
Esta es la vida del período de la cuarta glaciación, cuando un clima muy frío y húmedo prevaleció en toda Europa occidental, llegando hasta el norte de España y el norte de Italia. Si bien los campos glaciares no eran tan extensos como durante la tercera o la segunda glaciación, el clima era muy severo, como lo indica la migración hacia el sur no solo de la flora ártica, sino también de los mamíferos y aves de la región de la tundra que bordea las costas meridionales del océano Ártico. Dos o tres formas del frío [ p. 203 ] [ p. 204 ] [ p. 205 ] Las estepas del norte de Rusia también se extendieron a Europa occidental, pero este no fue claramente un período estepario debido a la humedad predominante del clima; en lugar de los vientos del oeste y las grandes nubes de polvo de los últimos tiempos del Achelense, frías nieblas y nubes cargadas de humedad inundaron el país, que durante los inviernos a veces quedaba cubierto de nieve y sujeto a rápidos cambios de temperatura. Estas condiciones climáticas[3] parecen demostrarse por el predominio de la vida de la tundra ártica, mamíferos adaptados únicamente al clima severo y atraídos por la flora del norte.

Los veranos eran indudablemente cálidos, como los actuales veranos árticos, pero mucho más largos en estas latitudes meridionales. No es improbable que hubiera migraciones estacionales, hacia el norte y el sur, de mamuts, rinocerontes y renos, y también que las canteras de sílex del norte, a lo largo del Somme y el Marne, fueran visitadas principalmente durante el cálido verano. Los mamíferos asiáticos habían desaparecido por completo de las regiones de Francia y Alemania durante el primer máximo de la cuarta glaciación, pero hay quienes sostienen que durante la mejora climática posterior, un intervalo en la región alpina denominado Laufenschwankung por Penck, el elefante de colmillos rectos y el rinoceronte de Merck volvieron a migrar al norte de Francia. Es cierto que ocasionalmente encontramos huesos de estos animales en estrecha asociación con los del mamut y el rinoceronte lanudos. Es posible explicar dicha mezcla como consecuencia del avance de la Cuarta Glaciación o como consecuencia de la migración hacia el norte y el sur de las respectivas manadas de mamíferos durante el verano y el invierno. A medida que avanzaba la Cuarta Glaciación, es seguro que estos mamíferos asiáticos desaparecieron por completo.
Al mismo tiempo, los neandertales habían pasado por la primera etapa de desarrollo de la industria musteriense y habían alcanzado lo que se conoce como el Tull o Musteriense ‘alto’, que, [ p. 206 ] con pocas excepciones, se llevaba a cabo al abrigo de los acantilados salientes o en el interior de las grutas.
La vida de los mamíferos de estos tiempos Musterienses “completos”, tal como se encuentra a lo largo de las cabeceras del Danubio, el Rin y los brazos del Dordoña y el Vère, se divide entre los diversos grupos faunísticos de la siguiente manera:
Vida en el Musteriense Medio
- Vida en la tundra.
- Mamut lanudo.
- Rinoceronte lanudo.
- Reno escandinavo.
- Zorro ártico.
- Liebre ártica.
- Lemming bandeado.
- Perdiz nival ártica.
- Vida alpina.
- Marmota alpina.
- Cabra montés.
- Perdiz nival alpina.
- Vida en la estepa.
- Caballo de estepa.
- Suslik de estepa.
- Gallina de agua.
- Vida asiática.
- León cavernario.
- Hiena de las cavernas.
- Leopardo de las cavernas.
- Vida en el bosque.
- Ciervo, lince, lobo, zorro, rata de agua, oso pardo, ciervo gigante.
-Oso de las cavernas.
- Ciervo, lince, lobo, zorro, rata de agua, oso pardo, ciervo gigante.
- Vida en el prado.
- Bisonte.
- Ganado salvaje.
Parecería que el reno, el mamut lanudo y el rinoceronte lanudo ya estaban ampliamente distribuidos por Europa occidental, acompañados por el zorro ártico (Cams lagopus), la liebre ártica (Lepus variabilis) y el lemming bandeado (Myodes torquatus). No hay pruebas de que el buey almizclero hubiera alcanzado en ese momento su distribución más meridional, y parecería que la llegada del segundo tipo de lemming del norte procedente de la región del río Obi (Myodes obensis) no se produjo hasta finales del Musteriense,21 porque la gran migración de estos animales está registrada por sus abundantes restos en la llamada «capa inferior de roedores» de todas las estaciones a lo largo del Rin y el Danubio, como Sirgenstein, Wildscheuer y Ofnet, tras la etapa final de la industria musteriense. De hecho, este notable roedor pequeño parece marcar el segundo máximo o el final de la cuarta glaciación con su migración por toda Europa occidental, y dondequiera que se encuentren sus restos en los depósitos de las grutas, proporcionan una de las fechas prehistóricas más importantes y positivas, a saber, la del estrato inferior de roedores. Los lemmings superan a todos los demás mamíferos en las grandes distancias recorridas por sus migraciones, y parece que esta especie septentrional se extendió por toda Europa occidental al mismo tiempo, dejando sus restos no solo en las cavernas a lo largo del Danubio, sino también en las de Bélgica y en Thiede, cerca de Braunschweig. Esta última estación, Thiede, no estaba lejos del límite sur del glaciar escandinavo; Estaba sometido a un clima ártico muy severo, ya que los únicos compañeros del lemming Obi eran el lemming anillado, el zorro ártico, la liebre ártica, el reno, el mamut y el buey almizclero.
El mamut lanudo alcanza ahora la cima de su evolución y especialización; tal como se preserva en las tundras heladas del norte de Siberia, y como se representa en numerosos dibujos y grabados de los artistas del Paleolítico Superior, es el fósil de mammaha más completamente conocido.22 Sus proporciones, como se muestra en la figura adjunta, que representa la información recopilada de todas las fuentes, son completamente diferentes a las del elefante indio o africano. La cabeza es muy alta y está rodeada por una gran masa de pelo y lana; detrás de esta, una depresión aguda separa la parte posterior de la cabeza de la gran joroba en la espalda; la parte posterior de la espalda cae muy rápidamente y la cola es corta; el manto de pelo largo casi llega al suelo, y debajo de este hay una cálida capa interna de lana. No es improbable que las jorobas en la cabeza y la espalda fueran depósitos de grasa. El color del pelo era un marrón amarillento, que variaba del marrón claro al marrón puro; El pelo lanoso, de entre dos y tres centímetros y medio de largo, cubría todo el cuerpo; entre los pelos más cortos se intercalaban numerosos pelos más largos y gruesos, que formaban melenas en las mejillas, la barbilla, los hombros, los flancos y el abdomen. Una amplia franja de este largo pelo se extendía a los lados del cuerpo, como se representa en las obras de los artistas del Paleolítico Superior de la cueva de Combarelles. Nos resulta especialmente interesante el alimento encontrado en el estómago y la boca de los mamuts siberianos congelados, que consiste principalmente en una flora de pradera similar a la que florece durante el verano en el norte de Siberia en la actualidad, incluyendo hierbas y juncias, tomillo silvestre, judías silvestres de la familia oxytropis y la variedad ártica del ranúnculo (Ranunculus acer). Este era el alimento de verano. Los alimentos de invierno incluían sin duda las hojas y los tallos del sauce, del enebro y de otras plantas de invierno.
La vida en el Musteriense tardío
Segundo Máximo de la Cuarta Glaciación
- Tundra, estepa, vida alpina, asiática y de pradera, como arriba.
-Obi lemming. - Buey almizclero.
- Armiño.
- Perdiz nival ártica.
- Comadreja de Eversmann (Comadreja de estepa).

El rinoceronte lanudo era el compañero invariable del mamut, así como el rinoceronte de Merck siempre se asociaba con el elefante de colmillos rectos. Este notable animal está emparentado con el grupo norteafricano de rinocerontes blancos, del cual se desprendió en un período muy remoto. El perfil de su cabeza, muy larga y estrecha, de su enorme cuerno anterior y posterior menor, y su lomo jorobado se asemeja al de la forma africana existente, pero su protección contra el clima ártico le dio una apariencia exterior completamente diferente; el pelo de la cara, de color marrón dorado, con una capa interna de lana, se conserva en el Museo de Petrogrado. Gracias a un descubrimiento en Starunia, en la Galicia oriental, en 1911, este animal es ahora completamente conocido por nosotros, excepto la cola. Sus restos fueron encontrados aquí a una profundidad de 30 pies, e incluían la cabeza, la pata delantera izquierda y la piel del lado izquierdo del cuerpo. El espécimen de Starunia tiene un labio superior ancho y truncado adaptado a los hábitos de pastoreo, ojos pequeños y oblicuos, orejas largas, estrechas y puntiagudas, un cuerno anterior largo con base ovalada y un cuerno posterior más corto, un cuello corto, en cuya parte posterior hay una pequeña joroba carnosa, bastante independiente del esqueleto; las patas son comparativamente cortas. Se diferencia de la forma africana viviente en el hocico algo más estrecho, en sus orejas pequeñas y puntiagudas y en la presencia de una gruesa capa de pelo. Al igual que el rinoceronte blanco, la forma lanuda era un habitante de las llanuras, viviendo de pasto y pequeñas hierbas.23 Este rinoceronte se mantuvo más cerca de los bordes de las grandes capas de hielo que el mamut, deteniendo su migración en Alemania y Francia; Es decir, no emigró tan al sur como el mamut, que vagó hacia Italia hasta llegar a Roma.
El reno fue el precursor de toda la fauna de la tundra ártica; llegó al valle del Vézère a principios del período de la verdadera cultura Musteriense y ya había penetrado mucho más al sur durante la Tercera Etapa Glacial, probablemente migrando a lo largo de los límites de los campos de hielo; de hecho, se encuentra en el norte de Europa incluso durante la segunda glaciación. Es la verdadera especie escandinava o de tierras áridas, que ahora se caracteriza por dos formas del reno del Viejo Mundo (R. tarandus, R. spitzbergensis) y por las formas americanas de tierras áridas existentes. Las astas son redondas, delgadas y largas en proporción al tamaño relativamente pequeño del animal; las puntas de la frente son palmeadas. Hay pocas pruebas de que los neandertales hicieran un gran uso de los huesos de reno, pero hay razones para suponer que utilizaban las pieles, para cuya preparación los raspadores y cepillos musterienses eran especialmente adecuados.
[ pág. 210 ]
En la península Ibérica, la fauna de la tundra no llegó tan al sur como Portugal, aunque el lemming noruego (Myodes lemmus) llegó a las cercanías de Lisboa. El mamut lanudo, acompañado del rinoceronte lanudo, se ha descubierto en dos localidades del extremo norte de España, en la provincia de Santander, junto al golfo de Vizcaya. El reno (Rangifer tarandus) se encuentra en la caverna de Seriha, al sur de los Pirineos; ya en la época achelense llegó a la región de Altamira, cerca de Santander. Por lo tanto, Harlé24 concluye que es seguro que la fauna de la tundra se extendió desde Francia hacia el oeste hasta Cataluña, a lo largo de la costa norte de España, flanqueando los Pirineos. Se cree generalmente que el oso de las cavernas (Ursus spelaus) ocupó muchas de las cavernas antes de que fueran ocupadas por el hombre, y desarrolló ciertas peculiaridades estructurales en estos lugares. Así, las falanges que sostienen las garras están poco desarrolladas, lo que indica que las garras habían perdido parcialmente su función prensil; los dientes anteriores de molienda están muy reducidos, y las cúspides de los dientes posteriores están romas, lo que indica una dieta omnívora. Sin embargo, las patas delanteras eran de un tamaño enorme, el cuerpo era robusto y de proporciones más pesadas que el de los osos europeos más grandes y recientes (Ursus arctos). Por lo tanto, parecería que los neandertales expulsaron de las cuevas a un tipo de oso menos formidable que la especie existente, pero sin embargo un serio oponente para los hombres armados con las armas pequeñas del período Musteriense.

¶ Costumbres de la Caza y de la Vida en las Cavernas
Solo disponemos de medios indirectos para conocer la valentía y la actividad de los neandertales en la caza, a través de los huesos de animales cazados para alimentarse, que se encuentran entremezclados con los pedernales en torno a sus antiguos hogares. Estos incluyen, en los hogares musterienses tempranos, como hemos visto, huesos de bisonte, ganado vacuno salvaje y caballo, a los que sigue en Combe-Capelle la primera aparición de huesos de reno. Tanto los huesos de bisonte como los de caballo salvaje se utilizan en los yunques de hueso de la cultura musteriense final de La Quina. Lo que consideramos el período de la gran vida mamífera de la región del Alto Danubio se encuentra en los niveles musterienses de la gruta de Sirgenstein, de donde parece que los neandertales cazaban mamuts, rinocerontes, caballos salvajes, bisontes y ganado vacuno, además de renos, ciervos gigantes. Debemos tener presente, sin embargo, que cuando estas cuevas quedaron desiertas por un tiempo, las bestias de presa regresaron, y así sucede a menudo que las capas sucesivas proporcionan pruebas de ocupación alternada por el hombre y por bestias de presa de tamaño suficiente para traer los tipos de caza más grandes, mientras que los búhos pueden ser responsables de los depósitos del lemming, como en la “capa inferior de roedores”.
Obermaier25 ha estudiado minuciosamente las vicisitudes de la vida en cuevas durante el Musteriense. Mucho antes de que estas cuevas fueran habitadas por el hombre, sirvieron de guaridas o refugios para el oso cavernario [ p. 212 ] y la hiena cavernaria, así como para muchas aves rapaces. Por ejemplo, la cueva de Echenoz-la-Moline, en las aguas superiores del Saona, contenía los restos de más de ochocientos esqueletos de oso cavernario, y sin duda los neandertales tuvieron que librar muchas y reñidas batallas antes de que las bestias fueran expulsadas y el hombre se adueñara de la gruta. Es posible que el fuego fuera el medio empleado. Se ha cuestionado si las cuevas eran moradas insalubres, pero debe recordarse que, salvo en ciertas cavernas con aberturas naturales en el techo para la salida del humo, no existía una verdadera vida cavernícola, sino una vida de gruta, centrada en la entrada de la cueva. La cueva más pequeña, observa este autor, era considerablemente más grande y mejor ventilada que las pequeñas y humeantes cabañas de algunos campesinos europeos o las chozas de nieve de los esquimales. El obstáculo más serio era la humedad predominante, que variaba periódicamente en las cavernas, de modo que a las estaciones secas sucedía una abundante humedad que se filtraba a través del techo de piedra caliza y por las paredes laterales. En esos momentos, las cavernas eran probablemente inhabitables, y en los huesos, tanto de hombres como de animales, se han observado muchos casos de hinchazones y de inflamación de las vértebras, como las causadas por la humedad extrema. Las ventajas compensatorias eran el refugio que ofrecían contra la lluvia y el frío, una temperatura constante a distancias moderadas de la entrada, y también el hecho de que las cuevas eran muy fáciles de defender, ya que la entrada era generalmente pequeña y el acceso a menudo empinado y difícil. Un alto muro de piedra que cruzara la abertura habría facilitado aún más la defensa, y una tea encendida habría impedido la aproximación de osos y otras bestias de presa. Debido a este refugio contra el clima y las fieras, las grutas y las aberturas más grandes de las cavernas estaban ciertamente llenas de trabajadores musterienses del sílex durante las estaciones inclementes del año.
Sin embargo, la mayor parte de la vida de los neandertales transcurría sin duda al aire libre y en la caza. Durante el Musteriense, las presas más comunes eran el caballo salvaje, el buey salvaje y el reno. Se utilizaban tanto la carne como las pieles, y la médula se obtenía partiendo los huesos más grandes. Así, con frecuencia encontramos en los hogares restos de mamut, rinoceronte lanudo, ciervo gigante, oso cavernario y oso pardo. De estas bestias de presa, los cazadores neandertales obtenían pieles y quizás también grasa para las antorchas que utilizaban para iluminar las cavernas; no hay pruebas de la invención de la lámpara en este período.
El trabajo de las mujeres, sin duda, consistía en preparar las comidas y convertir las pieles en cobertores y ropa. Siempre que era posible, esto se hacía a la luz del día, fuera de las grutas, pero en tiempo frío y lluvioso, o en el crudo invierno, toda la tribu se refugiaba en la gruta, reuniéndose alrededor de los fogones alimentados con leña; los rincones que encontraban servían como almacenes de combustible o carne seca, preservada para los días en que el frío extremo y la nieve cegadora impedían a los cazadores aventurarse.
Parece que la pieza de caza se desmembraba en el lugar donde caía, extrayéndose las mejores partes. El cráneo se abría para extraer el cerebro; los huesos largos se conservaban para la médula ósea; por lo tanto, los huesos del flanco y la espalda de la pieza son frecuentes en depósitos cavernarios, mientras que las costillas y las vértebras son escasas.
La trampa pudo haber formado parte de la técnica de caza conocida por los neandertales. La caza se realizaba con lanzas o dardos con puntas de sílex, también mediante piedras arrojadizas, que se encuentran en gran número en los niveles musterienses superiores de La Quina, en la cueva del Lobo de Yonne, Les Cottes y en varios lugares de España. Si imaginamos, como es muy posible, que la piedra arrojadiza se colocaba en una honda de cuero, en la punta hendida de un palo o sujeta a una larga correa de cuero, podemos ver fácilmente que resultaría un arma muy efectiva.
Los métodos de caza de los neandertales son, sin embargo, un misterio. Existía una marcada disparidad entre el tamaño y la eficacia de sus armas y la fuerza y resistencia de los animales que perseguían. No se conservó ninguno de los pesados instrumentos de la época achelense; las puntas de dardos y lanzas no han mejorado mucho; ciertamente, no podían penetrar las gruesas pieles de los mamíferos más grandes de la tundra ártica, fuertemente protegidas con pelo y lana; incluso la caza de caballos, ganado salvaje y renos aparentemente se realizaba sin la ayuda del arco y la flecha, y antes de la invención de la flecha con púas o la punta de lanza.
¶ Descubrimiento de las razas neandertaloides
La vida al aire libre o nómada de todas las tribus de Europa occidental, desde el Prechelense hasta casi el final del Achelense, fue muy desfavorable para la preservación de restos humanos. Es posible que los cuerpos de los muertos y de los ancianos fueran arrojados a las hienas que rodeaban las estaciones, como entre algunas tribus de África hoy en día, pero es igualmente posible que fueran enterrados de forma sonae. Los esqueletos enterrados cerca de la superficie en las arenas o gravas del río de las terrazas no se habrían conservado. Hemos visto que la preservación de los restos de Heidelberg y Piltdowm se debió enteramente al azar, ya que los huesos fueron arrastrados y mezclados con los de los animales. Tampoco se ha encontrado evidencia alguna en la gruta de Ehapina de enterramientos ceremoniales ni de respeto por los muertos, sino que, por el contrario, existen indicios de prácticas caníbales. Incluso antes del fin del Musteriense temprano, todo esto cambió. Quizás la estrecha relación impuesta por el clima más riguroso generó indirectamente un mayor respeto por los muertos y condujo a la costumbre del entierro o la disposición ordenada de los restos en los suelos de las grutas y cavernas parcialmente protegidas, costumbre a la que debemos nuestro conocimiento actual de la estructura del hombre de Neandertal en el Musteriense.
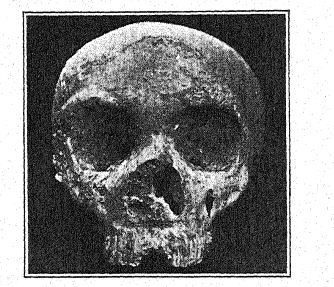
El primer descubrimiento de un neandertal se realizó en 1845, ocho años antes de que se revelara el tipo de la raza neandertal. Se trata del cráneo de Gibraltar26 hallado por el teniente Flint, cerca de la cantera Forbes, en la cara norte del Peñón de Gibraltar. Consiste en un cráneo bien conservado, al que solo le faltan los huesos parietales y la cara y la base del cráneo están notablemente completas. En 1868, Busk lo donó al Museo del [ p. 216 ] Real Colegio de Cirujanos de Londres, donde se encuentra actualmente. El lugar exacto del descubrimiento ya no puede identificarse con certeza. Probablemente se encontró en una cueva aún existente, y aunque no se puede determinar su edad arqueológica, dado que sus características anatómicas son las de la raza neandertal, y como todos los restos de esta raza que se pueden datar con certeza son de la era musteriense, probablemente pertenece al período musteriense. En los últimos años, su gran importancia en la historia de la humanidad se ha revelado en los estudios de Sollas, Keith y Schwalbe. Por lo tanto, se ha clasificado entre los neandertaloides y se considera una forma particularmente primitiva, debido al tamaño extremadamente pequeño del cerebro. Esta característica y el ligero desarrollo de las crestas supraorbitales, tan característico de los neandertaloides, se explican por la teoría de que el cráneo perteneció a una mujer.
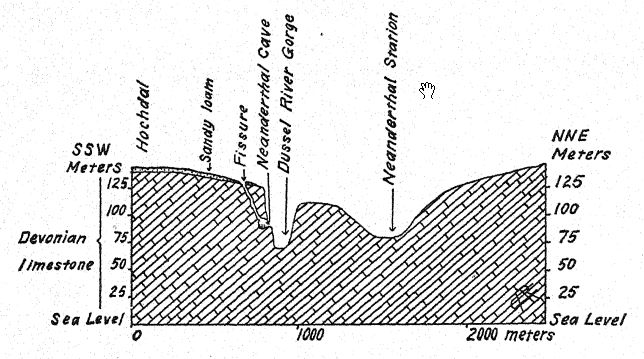
Sera27 considera que el cráneo de Gibraltar es el más simiesco de todos los fósiles humanos y opina que no debería clasificarse con los neandertaloides, sino que debería considerarse preneandertaloides; Keith comparte esta opinión. Boule, sin embargo, cree que este cráneo tiene la misma edad geológica que los de Spy, La Chapelle, La Ferrassie y La Quina; todo nos lleva a creer,28, que el cráneo de Gibraltar es un cráneo femenino de tipo neandertal. En otro lugar, se refiere a los cráneos de Gibraltar, [ p. 217 ], de La Quina y de La Ferrassie II como probablemente los de mujeres neandertales.
El cráneo tipo de esta gran raza extinta de hombres es el del neandertal —sin duda el más famoso y el más controvertido de todos los restos antropológicos—, apreciado por Lyell y Huxley, pero ignorado por Darwin, y finalmente establecido por Schwalbe como el eslabón perdido más importante entre la especie humana actual (Homo sapiens) y los simios antropoides. En 185629, unos obreros estaban trabajando en la limpieza de una pequeña cueva cubierta de marga, de unos dos metros de altura, la llamada Gruta Feldhofner, en la caliza cretácica del valle conocido como el Neandertal, en el pequeño arroyo Düssel, que fluye entre Elberfeld y Düsseldorf. Descubrieron algunos huesos humanos, probablemente un esqueleto completo que representaba un enterramiento, que, por desgracia, se dejó esparcir y triturar. El doctor Fuhlrott rescató las partes restantes, incluyendo el ahora famoso casquete craneal, ambos fémures, el húmero derecho, partes del antebrazo, huesos de ambos lados, la clavícula derecha y fragmentos de la pelvis, el omóplato y las costillas. Todos los huesos se conservaron en perfecto estado y ahora se encuentran en el museo provincial de Bonn.

El descubrimiento causó gran sensación, pero al principio la edad de estos fósiles permaneció en duda; a unos 150 pasos de la gruta, en una pequeña cueva similar, se encontraron huesos de oso cavernario y rinoceronte. En 1858 se publicaron las memorias de Schaaffhausen30, en las que ofreció la primera descripción detallada de estos restos como pertenecientes a una raza primitiva original que difería en todo del hombre actual, y nunca dudó de esta postura. [ p. 218 ] En 186331 Busk, Huxley y Lyell también situaron este esqueleto en su verdadera posición intermedia entre el hombre y los simios antropoides. La firme opinión de Virchow de que éste no era un tipo normal de hombre ejerció una influencia tan grande que no fue hasta el trabajo clásico de Schwalbe,32 entre 1899 y 1901, que este esqueleto asumió su importancia dominante para todos los tiempos, e incluso esto fue posterior al descubrimiento de otras dos razas neandertaloides.
Al principio, erróneamente, se asoció con la llamada raza de Cannstatt, pero mucho antes del trabajo de Schwalbe, King,33, en 1864, lo reconoció como una especie distinta de hombre (Homo neanderthalensis), «el hombre del valle de Neander». Poco después del descubrimiento de los taloides de Meander de Spy, en Bélgica, Cope,34, propuso en 1893 el mismo nombre específico de Homo neanderthalensis_. En 1897, Wilser35 sugirió el nombre de Homo primigenius, ampliamente adoptado en Alemania, mientras que entre los autores franceses, la misma especie de hombre se conoce hoy en día como Homo mousieriensis. Esta variedad de nombres sirve al menos para dejar constancia de la opinión unánime de que este hombre del Pleistoceno medio pertenece a una especie distinta.
Dado que la raza estaba muy extendida, podemos hablar de estos pueblos como «neandertales», mientras que las razas similares a la especie neandertal pueden caracterizarse como «neandertaloides». El conjunto de descubrimientos de miembros de esta raza es actualmente muy extenso.
En 1887, los geólogos belgas Fraipont y Lohesy descubrieron en una gruta cerca de Spy, no lejos de Dinant, en el Mosa, los restos de dos individuos que ahora se identifican como Spy I y Spy II. En el mismo estrato que los esqueletos, bajo una capa de caliza tobácea, se incrustaron herramientas de sílex del Musteriense, junto con restos de mamut lanudo, rinoceronte lanudo, oso cavernario y hiena cavernaria. Este descubrimiento es uno de los más importantes en la historia de la antropología, ya que data definitivamente a los hombres de Spy como pertenecientes al período de la industria musteriense, y también porque los autores reconocieron inmediatamente a estos hombres como pertenecientes a la raza de [ p. 219 ] [ p. 220 ] Neandertal y de Cannstatt, que en aquel entonces se suponía que eran idénticos. Aquí se conocieron por primera vez las proporciones del cráneo y el cerebro, los rasgos primitivos de la mandíbula inferior y de los dientes, la baja estatura y varias características simiescas de los huesos de las extremidades; se observaron las prominentes crestas supraorbitales del tipo neandertal, la frente retraída, el perfil craneal inferior al de las razas australianas más bajas existentes, y el cráneo estrecho y dolicocefálico.
DISTRIBUCIÓN DE LOS RESTOS DE LOS NEANDERTALES (Compárese con la Fig. 104)
| I. De tiempos desconocidos del Paleolítico inferior | |||
| 1848. | Gibraltar. | Cantera Forbes. | Cráneo fragmentario. |
| 1856. | Neandertal. | Düsseldorf, Alemania. | Cráneo y fragmentos esqueléticos. |
| 1859. | Arcy-sur-Cure. | Yonne, Francia. | Bajo la mandíbula. |
| 1866. | La Naulette. | Bélgica. | Mandíbula inferior. |
| 1888. | Malarnaud. | Ariège, Francia. | Mandíbula inferior. |
| ?Gourdan. | Altos Pirineos. | Mandíbula inferior. | |
| 1906. | Ochos. | Moravia. | Mandíbula inferior. |
| 2. Con industria del Musteriense tardío | |||
| 1887. | Espía 1, 11. | Cerca de Dinant, Bélgica. | Dos cráneos y esqueletos. |
| 1907. | Petit-Puymoyen. | Char elite, Francia. | Fragmentos de mandíbulas superior e inferior. |
| 1909. | Pech de l’Azé. | Dordoña, Francia. | Cráneo de un niño. |
| 1910. | La Ferrassie II. | Dordoña, Francia. | Yo esqueleto (hembra). |
| 1911. | La Cotte de St. Brelade. | Isla de Jersey. | 13 dientes humanos. |
| 1911. | La Quina II. | Charente, Francia. | Cráneo y fragmentos de esqueleto. |
| 3. Con la industria del Musteriense Medio | |||
| 1982. | Sipka. | Moravia. | Mandíbula de un niño. |
| 1908. | La Chapelle-aux-Saints. | Corrèze, Francia. | Cráneo y esqueleto casi completos. |
| 1909. | La Ferrassie I. | Dordoña, Francia. | Porciones de un esqueleto. |
| 1910. | La Quina I. | Charente, Francia. | Huesos del pie. |
| 4. Con la industria musteriense temprana | |||
| 1908. | El Moustier. | Valle de Vézère, Dor Dogne, Francia. | Esqueleto de un joven. |
| 1914. | Ehringsdorf. 37 | Cerca de Weimar. | Mandíbula inferior. |
| 5. Con industria musteriense o achelense | |||
| 1899. | Krapina. | Croacia, Austria-Hungría. | Fragmentos de numerosos esqueletos de adultos y niños. |
| 1892. | Taubach. | Cerca de Weimar. | Diente de leche. |
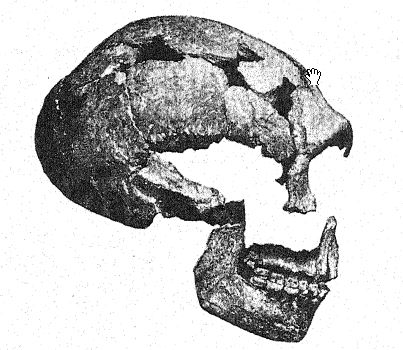
Se descubrió que las extremidades conservaban la desproporción antropoide entre el fémur y la tibia, y se hizo el importante descubrimiento de que este hombre de Neandertal, de baja estatura, complexión robusta, frente poblada y rostro opaco, no podía mantenerse completamente erguido, ya que la estructura de la articulación de la rodilla le permitía mantenerlas ligeramente flexionadas constantemente. En otras palabras, el Hombre Espía aún no había adquirido plenamente la posición erguida de las extremidades inferiores.
Este descubrimiento puede decirse que estableció a los neandertales en todos sus caracteres como una raza inferior muy distinta, pero transcurrieron veintidós años antes de que esto se confirmara aún más con el hallazgo de otro tipo aún más temprano de neandertaloides en Krapina, en el norte de Croacia, Austria-Hungría, como se describe al final del Capítulo II (pág. 181 arriba); un tipo que con sus variaciones locales [ pág. 221 ] pronto se determinó como perteneciente incuestionablemente al mismo grupo que el hombre de Neandertal y los hombres de Spy.
Muchos años antes, concretamente en 1866, el antropólogo belga Dupont38 había descubierto los restos de otro miembro de esta raza en una gruta a orillas del río Lesse, cerca de La Naulette, no lejos de Furfooz, en el norte de Bélgica. Esta, conocida actualmente como mandíbula de La Naulette, se considera de tipo neandertal. Se asoció con huesos de mamut lanudo, rinoceronte, reno y algunos fragmentos de otros huesos humanos.
De nuevo, en 1882, Maska39 encontró en una cueva cerca de Sipka, en Moravia, al sur de Sternberg y a seis millas al este de Neutitschein, fragmentos de la mandíbula inferior de un niño, extraordinariamente fuertes, gruesos y grandes, que mostraban la aparición de los dientes permanentes. De esta misma región proviene la mandíbula de Ochos, Moravia, hallada por Rzehak40 alrededor de 1906. Solo se encontró la parte alveolar de la mandíbula, pero sirvió para demostrar la amplia distribución geográfica de la raza neandertal.
En esa época, la región de Dordoña, conocida desde hacía tiempo como un centro intensivo de industria musteriense, desde el descubrimiento de Le Moustier por Lartet en 1863, no había aportado ni un solo esqueleto ni evidencia anatómica del tipo de hombre que la habitó en la época musteriense. Sin embargo, a partir de la primavera de 1908 se sucedieron varios descubrimientos similares, principalmente de entierros ceremoniales, en La Chapelle-aux-Saints, en la estación tipo de Le Moustier, en La Ferrassie, otra estación en el bajo Vézère, y en La Quina.
En octubre de 1910 se descubrió el cráneo conocido como La Ferrassie II, de edad Musteriense tardía; probablemente se trata de una mujer, y los restos fueron dispuestos en lo que presumiblemente fue una forma especial de entierro ceremonial, porque los huesos, en lugar de estar dispuestos rectos en una dirección determinada, estaban en una posición agachada o flexionada (ver Apéndice, Nota X).
El esqueleto de Le Moustier fue hallado por Hauser en la gruta inferior de Le Moustier, en el valle del Vézère, en la primavera de 1908, y extraído cuidadosamente con la ayuda del profesor Klaatsch.[ p. 222 ] Pertenecía a un joven de unos dieciséis años. La característica más interesante del descubrimiento fue la disposición del esqueleto.[ p. 222 ] La cabeza descansaba sobre varios fragmentos de sílex cuidadosamente apilados, una especie de almohada de piedra; el cadáver yacía dormido, con la cabeza apoyada en el antebrazo derecho. Un coup de poing excepcionalmente fino estaba cerca, y numerosos huesos carbonizados y partidos de ganado salvaje (Bos primigenius) estaban colocados alrededor, lo que indicaba una ofrenda de comida. Se creía que los pedernales pertenecían al Achelense, que subyace al estrato de auténtica industria musteriense, conocida desde hace tiempo en esta localidad; sin embargo, los arqueólogos franceses y Schmidt consideran que estos instrumentos pertenecen al Musteriense más temprano, en el que es bien sabido que aún persistía el coup de poing achelense. Desafortunadamente, el esqueleto no estaba muy bien conservado y, si bien Edaatsch tenía toda la razón al clasificarlo con los neandertaloides, no debería considerarse una especie distinta (Homo mousteriensis hauseri), sino más bien un miembro de la auténtica raza neandertal (Homo newnderthalensis). También resulta ser un individuo bastante robusto, robusto y de baja estatura: los brazos y las piernas son relativamente cortos, especialmente el antebrazo y la tibia.

[ pág. 223 ]
Al mismo tiempo que se desenterraba el esqueleto de Le Moustier, los abades A. y J. Bouyssonie, y L. Bardon43 exploraban la cultura musteriense de la gruta cercana a La Chapelle-aux-Saints, a pocos kilómetros al este de Le Moustier, y encontraron un esqueleto que ha demostrado ser, con diferencia, el mejor de todos los fósiles neandertaloides, incluyendo un cráneo notablemente bien conservado, casi toda la columna vertebral, veinte costillas, huesos del brazo y de la mayor parte de la pierna, y varios huesos de las manos y los pies. Este fue también un entierro ceremonial de un individuo de entre cincuenta y cincuenta y cinco años de edad, cuidadosamente dispuesto en dirección este-oeste en una pequeña depresión natural. Junto con él se encontraron pedernales típicos del Musteriense, también varias conchas y restos principalmente de rinoceronte lanudo, caballo, reno y bisonte. El hallazgo de un cráneo maduro con los huesos faciales en su posición y en un estado de conservación relativamente perfecto, sin distorsión de todo el cráneo, brindó por primera vez la oportunidad de determinar finalmente no solo todas las características y proporciones esqueléticas del hombre de Neandertal, sino también el tamaño y las proporciones reales del cerebro. Este magnífico ejemplar fue enviado al Museo de París, y las descripciones preliminares de Boule44 y, finalmente, su monografía casi impecable45 despertaron el interés mundial por la raza neandertal.

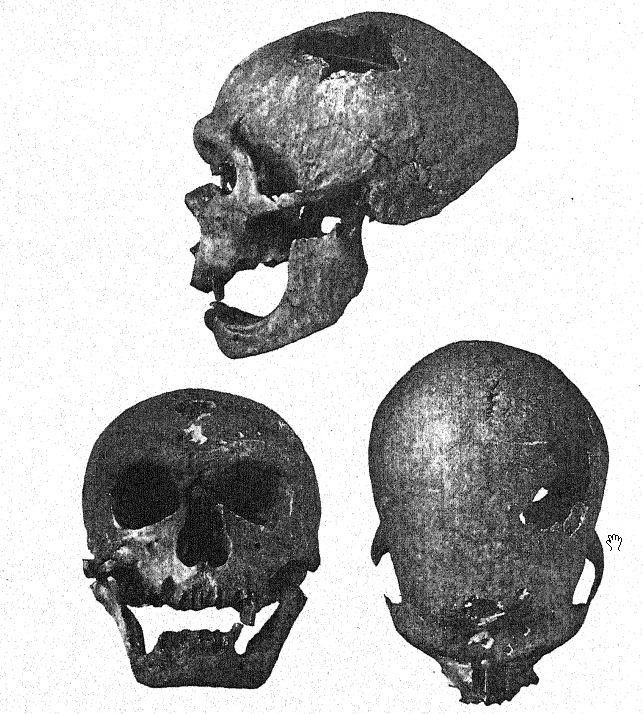
Un año después, Peyrony descubrió un tercer esqueleto neandertal en la cueva de La Ferrassie, cerca de Le Bugue, Dordoña. Los huesos estaban muy fragmentados y las pruebas de un entierro ceremonial no eran del todo claras, pero a simple vista, las características del cráneo, y en particular las de la frente, permitían reconocer al esqueleto como perteneciente a la raza neandertal.
En el año siguiente, 1910, en la caverna de La Quina, departamento de Charente, al norte de la región de Vézère46, se hallaron los huesos del pie de un hombre que se asemejaba exactamente al tipo de La Chapelle, y de nuevo en 1911 se descubrieron varias partes del esqueleto de otro miembro completamente típico de la raza neandertal en los primeros estratos musterienses. Los huesos del cráneo estaban algo separados en las suturas. Ciertamente, no se trataba de un entierro ceremonial. Al igual que el cráneo de Gibraltar, se supone que corresponde a una mujer.

De especial interés geográfico es el descubrimiento por Nicolle y Sineh47 de trece dientes humanos en una caverna musteriense en la bahía de St. Brelade, en la isla de Jersey48, lo que prueba la extensión de la raza neandertal a las Islas del Canal, cuando estas, con toda probabilidad, aún formaban parte del continente. Los dientes se asociaban con huesos de rinoceronte lanudo, de reno y de dos variedades de caballo, así como con evidencias de hogares musterienses e instrumentos de sílex. Las características distintivas de los dientes de molienda neandertales son el tamaño robusto, la implantación profunda y la forma expandida de las raíces, que, junto con la mandíbula pesada, indican la dureza de los alimentos y la fuerza muscular ejercida durante la masticación. [ p. 226 ] Las raíces, en lugar de estrecharse hacia abajo, como en el hombre moderno, forman una columna ancha y robusta que sostiene la corona, adaptada al movimiento de barrido de la mandíbula. Esta característica especial, por sí sola, excluiría a los neandertales de la ascendencia de las razas superiores.
Así, a través de una larga serie de descubrimientos, iniciados en 1848 y multiplicándose rápidamente durante los últimos años, hemos encontrado los materiales para un conocimiento completo de la estructura esquelética de los hombres, mujeres y niños de la raza neandertal; conocemos el desarrollo relativo del cerebro, así como la estatura de los sexos; hemos determinado que esta raza, y sólo ésta, se extendió por toda Europa occidental durante el Achelense tardío y todo el período del Mbusteriano, y también hemos aprendido que era una raza imbuida de reverencia hacia los muertos y, por lo tanto, probablemente animada por la creencia en alguna forma de existencia futura.
¶ Personajes de la raza neandertal
Los cráneos y esqueletos49 de Neandertal, Spy, Krapina, Le Moustier, La Chapelle, La Ferrassie y Gibraltar comparten tantas características distintivas que es indudable que deben clasificarse en un grupo estrechamente relacionado. Las características distintivas de este grupo son:
En primer lugar, características que se encuentran también en las diferentes razas existentes de hombres, pero nunca en los simios antropoides y, por tanto, en los humanos; en segundo lugar, características que nunca se han encontrado combinadas en ninguna raza de hombre reciente, por lo que el grupo representa una especie distinta de hombre; en tercer lugar, características fuera de los límites de variación de las razas recientes de hombres, e intermedias entre ellas y los límites de variación de los simios antropoides.
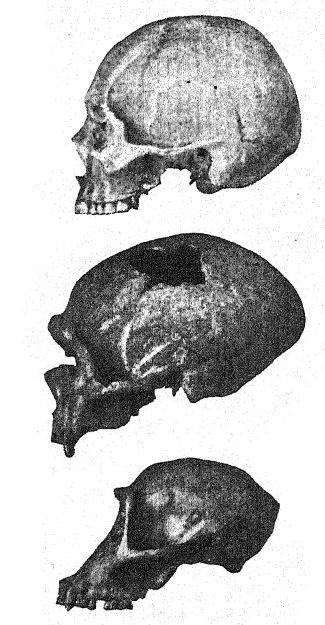
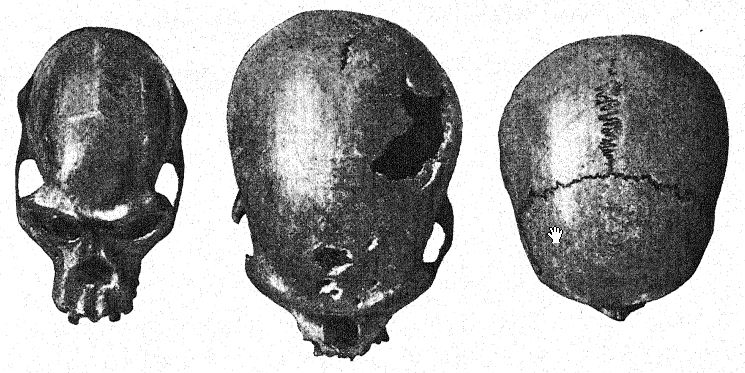
Antes de analizar al hombre de Neandertal en su conjunto, podemos centrar nuestra atención especialmente en varios de estos rasgos peculiares de la raza. Todos los primeros observadores quedaron impresionados por las cejas pobladas y prominentes y la frente hundida. En el hombre reciente, suele existir una marcada prominencia sobre los ojos, desde la glabela o punto medio por encima de la nariz hacia afuera, pero generalmente el tercio exterior del borde de estas prominencias [ p. 227 ] gira hacia arriba bajo la línea exterior de las cejas. En los neandertales, por el contrario, estas prominencias bajo las cejas rodean todo el borde superior de la cuenca ocular, extendiéndose hacia afuera alrededor de los bordes externos de la frente, por lo que pueden denominarse «tori supraorbitales»; la extensión de esta prominente cresta por encima y a los lados forma un verdadero techo sobre las cuencas oculares, que parecen dos profundas cavernas laterales. Estas prominencias laterales aparecen, aunque raramente, en el hombre reciente; se observan, por ejemplo, en ciertos australianos.
La vista frontal del rostro neandertal, como se observa en el cráneo femenino de Gibraltar, en el que las crestas superciliares no son tan prominentes como en los cráneos masculinos, es igualmente notable por la gran altura del rostro en comparación con la planitud de la frente. Al comparar el cráneo con el del australiano,50 observamos de inmediato la enorme diferencia en las proporciones del rostro y el cráneo entre estos dos tipos, aunque el australiano representa una de las razas más bajas existentes de Homo sapiens; observamos en el cráneo de Gibraltar el amplio espacio entre los ojos y el gran tamaño de la abertura nasal, lo que indica una nariz ancha y aplanada; existe un espacio correspondientemente largo entre la base de la abertura nasal y la línea de inserción de los incisivos, lo que indica un labio superior muy largo.
[ pág. 228 ]
La mandíbula es menos potente que la del hombre de Heidelberg. Hemos visto que la mandíbula de Heidelberg se distingue por su fuerza y torpeza, y por la ausencia de mentón, o mejor dicho, un mentón sin la más mínima prominencia. En el interior de esta placa mentoniana, muy gruesa y redondeada, falta la característica espina mentoniana (spina mentalis); en su lugar, se observa un doble surco como punto de inserción de los músculos que conectan el mentón y la lengua con el hueso hioides. El proceso ascendente para la inserción de los músculos mandibulares es inusualmente ancho, de 60 mm, en contraste con los aproximadamente 37 mm de la mandíbula reciente; finalmente, el cóndilo para la inserción con el cráneo es particularmente grande.
Al igual que la mandíbula de Heidelberg, la de los neandertales se distingue por su gran grosor y solidez. En general, los contornos son similares; en algunos casos, el proceso del mentón se sugiere por una ligera prominencia, pero en general, el mentón es muy retraído y coincide con la de Heidelberg en la ausencia de la espina mentalis. En otras características, existen marcadas diferencias entre las mandíbulas de Heidelberg y la de los neandertales. La forma de esta última se conoce actualmente a partir de los especímenes de Krapina, Spy, La Naulette, Ochos y Sipka, así como de los ejemplares perfectos de Le Moustier y La Chapelle. El espécimen de Sipka [ p. 229 ] demuestra que incluso en un niño de diez años, la mandíbula era notable por su grosor y fuerza. Boule52 está completamente de acuerdo con Gorjanovic-Kramberger53 en que el mentón en la mandíbula del neandertal estaba sólo en proceso de formación, y a lo largo de la vida no alcanzó más que una forma infantil, y que los neandertales pueden ser clasificados, sin embargo, como Homines mentales, mientras que los Heidelberg, en los que el mentón falta por completo, pueden ser considerados como Homines amentales.
Las proporciones de los dientes en los neandertales son igualmente distintivas, especialmente en el tamaño de los verdaderos dientes de corte y de molienda. Al igual que en la mandíbula de Heidelberg, forman una hilera densa, de la cual el canino no sobresale como en la dentición de Piltdown; de hecho, el contorno de la mandíbula y las proporciones de los dientes son claramente humanos en comparación con la mandíbula anaranjada del hombre de Piltdown. La superficie de molienda de los dientes presenta numerosas capas de esmalte, y las cúspides están bien desarrolladas. A diferencia de los del hombre reciente, los incisivos presentan pliegues de esmalte en las superficies internas o linguales, una condición rara vez observada en los dientes de corte modernos. En los dientes de la mandíbula de Heidelberg, las cavidades pulpares son excepcionalmente grandes, mientras que en los dientes de la raza Krapina se observa la peculiar característica de que los molares carecen de raíces normales, ya que estas han sido más o menos absorbidas, una ocurrencia muy rara en el hombre reciente. La dentición de La Chapelle es también claramente humana, pero extraordinariamente masiva, en correspondencia con la masividad general del cráneo y del aparato masticatorio; en detalle, no es la de las razas civilizadas, sino una forma exagerada del tipo llamado macrodonte.54 El alargamiento de la corona también es similar a lo que se denomina hipsodonte.
No todos los dientes de molienda presentan este tamaño masivo y forma columnar, ya que aproximadamente el cincuenta por ciento de los dientes de Krapina tienen raíces bien definidas y se asemejan más a los dientes de molienda modernos. En los neandertaloides de Spy, los dientes son pequeños y las raíces son de tamaño moderado.
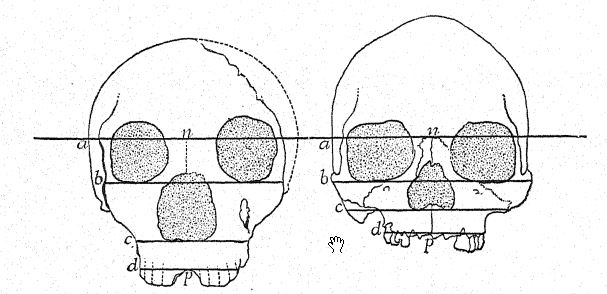
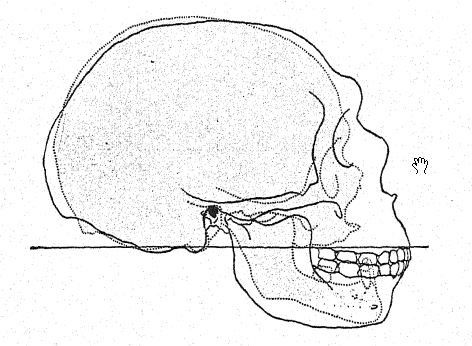
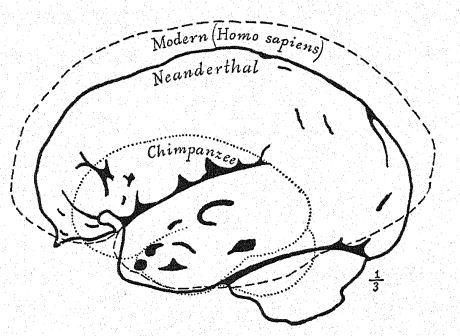
Este estudio de la frente y de los arcos superciliares, de la gran profundidad del rostro y de la forma peculiarmente alta y cuadrada de las cuencas oculares nos prepara para una vista de perfil del cráneo de [ p. 230 ] La Chapelle en contraste con el del tipo europeo más desarrollado e intelectual, es decir, el perfil del distinguido paleontólogo estadounidense, el difunto profesor Edward D. Cope, quien legó su cráneo y esqueleto para fines de estudio y comparación científica. En La Chapelle, observamos de inmediato la platicefalia (aplanamiento del cráneo), la frente retraída, la gran prominencia de las cejas, similar a la de los simios antropoides, el alargamiento del rostro en comparación con el aplanamiento del cráneo, la gran prominencia o prognatismo del rostro en su conjunto, la especial prominencia de las hileras de dientes cortantes en comparación con la línea vertical o hacia adentro, y la recesión de la hilera de dientes en el perfil de Cope. Esta comparación también resalta el marcado contraste entre la alta prominencia del mentón del Homo sapiens y el mentón profundamente hundido de los neandertales. El contraste es igualmente notable en la vista superior del cráneo, donde se observa que el tipo neandertal es extremadamente dolicocéfalo, con la parte posterior del cráneo relativamente ancha y la parte frontal estrechándose en la región del prosencéfalo hasta expandirse repentinamente en los prominentes procesos supraorbitales.
Como se muestra en el diagrama de la página 8, Fig. 1, la mayor longitud del cráneo neandertal se encuentra en la línea horizontal que atraviesa la cavidad cerebral, conocida como línea glabela-inión. Esta línea se traza desde una prominencia entre los arcos superciliares hasta un punto en la parte posterior del cráneo conocido como protuberancia occipital externa o inión. Esta línea también es la más larga en los cráneos de Spy y La Chapelle, así como en los simios antropoides,< … La sección media del cráneo del chimpancé, del neandertal y del norte de Australia muestra de forma muy impactante la generalización de Schwalbe en 1901, según la cual el cráneo del neandertal es en realidad una forma intermedia entre el de los simios antropoides y el del Homo sapiens. En esta esclarecedora sección, observamos el crecimiento de la cúpula craneal, es decir, la gran cavidad que alberga el cerebro por encima de la línea glabela-inión (g-i), observando el contraste entre la longitud de la línea vertical de la altura craneal y el espacio por debajo de dicha línea, indicado por las letras. Esta importante línea vertical termina en la abertura donde la médula espinal penetra en la base del cerebro (véase la figura 1).
[ pág. 232 ]

En muchas características, el cráneo neandertal se asemeja más al de los simios antropoides que al del Homo sapiens. Esta conclusión, a la que llegó Schwalbe en 1901,57 ha sido más que confirmada por el magistral estudio de Boule58 del cráneo muy completo de La Chapelle. Después de su revisión detallada, concluye: En cuanto a la unidad de la forma de la cabeza neandertal, estas características no son exclusivas del cráneo de La Chapelle; en todos los casos también se encuentran en los cráneos de Neanderthal, Gibraltar, Spy, Krapina, La Ferrassie, que dan testimonio de la homogeneidad de ese tipo fósil humano llamado neandertal. Estas características muestran una afinidad estructural entre los hombres fósiles del período Musteriense y los simios antropoides. Debe notarse que muchas de estas características también pueden encontrarse en cráneos humanos recientes de las razas inferiores, pero que son muy raras, muy dispersas, muy aisladas y ocurren solo como aberraciones. Es la acumulación de todas estas características en cada cráneo de una serie completa lo que constituye un conjunto enteramente nuevo y de gran importancia. En el cráneo, como en otras partes de la anatomía de los neandertales, no deberíamos esperar encontrar todos los caracteres intermedios entre los antropoides y el hombre reciente. El largo rostro neandertal es algo similar al del esquimal y contrasta con el rostro muy corto de los australianos y tasmanos actuales. La depresión en la raíz de la nariz, justo debajo de la glabela, es muy marcada en todos los neandertales; hay menos puente nasal que en cualquier raza reciente, excepto en los australianos varones, pero la nariz no es aplanada sino algo arqueada o aguileña. Este rasgo no es característico de todos los simios antropoides, y en este sentido los neandertales, australianos y tasmanos son más diferentes de los simios antropoides que algunas de las razas blancas; Así, la nariz neandertal, lejos de parecerse a la de los antropoides, difiere de ella más que la de algunos tipos humanos recientes.59 Muchos anatomistas, siguiendo a Huxley, han descrito los cráneos australianos y tasmanos como más o menos neandertaloides, y algunos autores han llegado incluso a considerar a estas razas como neandertales supervivientes. Es cierto que algunos cráneos de estas razas existentes son extraordinariamente platicefálicos y muestran una frente retraída, que otros presentan crestas supraorbitales casi tan prominentes como en los neandertales, que a veces la prominencia del occipital es muy marcada, y que ciertas mandíbulas muestran un mentón muy retraído. Así, se ha observado uno u otro de estos rasgos neandertales en estas razas inferiores existentes, pero todas estas características nunca se han combinado en una raza como rasgos constantes e invariablemente asociados, como en todos los cráneos de neandertales que conocemos.
En resumen, el tipo de cabeza australiano no tiene nada en común con el de los neandertales, salvo en un pequeño número de características en la región de la frente y la nariz. Los rasgos distintivos de la cabeza y el rostro neandertales son la platicofagia (frente retraída), el aplanamiento del occipucio (la porción inferior del cráneo), la prominencia de las crestas supraorbitales, el mentón retraído o ausente, la proyección de todo el rostro debido a la peculiar forma de la mandíbula superior y el tamaño relativamente pequeño de los lóbulos trontales del cerebro. De hecho, concluye Boule: «Todos estos llamados ‘neandertaloides’ modernos no son más que variedades de individuos del Homo sapiens, notables por la exageración accidental de ciertos rasgos anatómicos que se desarrollan normalmente en todos los especímenes del Homo neanderthalensis. La explicación más simple de estos accidentes, en la mayoría de los casos, es el atavismo o la reversión. No podemos afirmar que nunca haya habido una infusión de sangre neandertal en los grupos pertenecientes a la especie Homo sapiens, pero lo que parece bastante seguro es que dicha infusión pudo haber sido solo accidental, pues no existe ningún tipo reciente que pueda considerarse siquiera un descendiente directo modificado de los neandertales».
Esta opinión es confirmada por las últimas y más exhaustivas investigaciones de Berry y Robertson,60 quienes concluyen que ni los australianos ni los tasmanos tienen ninguna relación directa con el Homo neanderthalensis ; los puntos superficiales de semejanza craneal son explicables únicamente sobre la base de la lejanía de la ascendencia. Los australianos y los tasmanos son descendientes no del linaje neandertal sino de un linaje del Plioceno tardío o del Pleistoceno temprano, que, siguiendo a Sergi, puede llamarse Homo sapiens tasmanianus, del cual el aborigen tasmano, ahora extinto, fue el descendiente casi inalterado. Con respecto a los caracteres ‘bajos’, como se muestra en el diagrama. Fig. 117, los cráneos de Spy-Neanderthal se encuentran bastante cerca de los tasmanos y australianos, y el cráneo de Gibraltar se encuentra a medio camino entre este tipo y Pithecanthropus con respecto a doce caracteres diferentes de comparación.
Es interesante notar[4] que los tasmanos fueron encontrados en una etapa de la industria del sílex muy similar a la practicada por los neandertales en tiempos musterienses; sus pedernales fueron hechos de lascas producidas artificialmente, incluyendo unos pocos ejemplos61 que exhibieron una pulcritud en el recorte de los bordes y una regularidad resultante en el contorno, mientras que la mayor parte se caracterizaban por un recorte torpe y un contorno irregular; el bajo estatus de los implementos tasmanos puede ser descrito más correctamente con la palabra Pre-Auriñaciense, es decir, del Musteriense o de una etapa anterior, pero de ninguna manera ‘Eolítico’.
[ pág. 235 ]
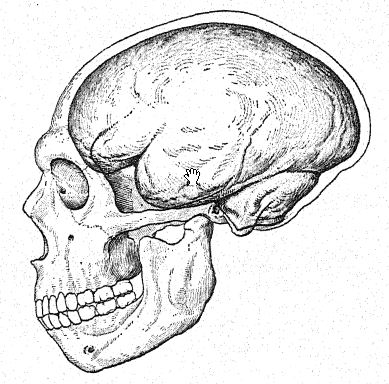
Se sabía que el cerebro del hombre de Neandertal era de gran tamaño, incluso al estimarlo a partir del tipo thal. Darwin se vio obligado a admitir que el famoso cráneo del hombre de Neandertal estaba bien desarrollado y era espacioso, y Broca ofreció una ingeniosa explicación del hecho, por lo demás inexplicable, de que la capacidad media del cráneo del antiguo cavernícola es mayor que la de muchos franceses modernos: que la capacidad media del cráneo en las naciones civilizadas debe verse reducida por la preservación de un número considerable de individuos, débiles mental y físicamente, que habrían sido rápidamente eliminados en estado salvaje, mientras que entre los salvajes el promedio incluye solo a los individuos más capaces que han logrado sobrevivir en condiciones de vida extremadamente duras. Los cráneos de La Chapelle y de Spy brindaron la oportunidad de resolver este interesante problema, y los resultados confirman plenamente las estimaciones previas de Schaaffhausen y de Broca sobre la gran capacidad cúbica del cerebro neandertal. Las estimaciones, en orden descendente, son las siguientes:
[ pág. 236 ]
| Cráneo de Spy II (Fraipont) | ? 1723 c. cm. |
| “ La Chapelle (Boule, Verneau y Rivet) | 1626 “ |
| «Espía I (Fraipont) | 1562» |
| «Neandertal | 1408» |
| “ La Quina, hembra (aproximación Boule) | 1367 “ |
| Gibraltar, hembra (estimación de Boule) | 1296 |
El tamaño del cerebro en las razas actuales de Homo sapiens varía de 950 cm³ a 2020 cm³. Así, en cuanto al volumen de materia cerebral, el cerebro del hombre de Neandertal es sin duda humano, pero en su forma carece de las proporciones características de la organización cerebral superior del hombre reciente. En otro aspecto importante, es humano: el mayor tamaño del hemisferio izquierdo, lo que indica el desarrollo del uso de la mano derecha. En su forma general, el cerebro se asemeja más al de los simios antropoides en el tamaño relativamente menor de la porción frontal, la simplicidad y longitud de las circunvoluciones, y en la posición y dirección de las grandes fisuras laterales conocidas como «fisuras de Silvio» y «fisuras de Rolando». Según lo estudiado por Boule y Anthony64, existen muchas características primitivas en el cerebro de los neandertales. La parte frontal del prosencéfalo, la llamada área prefrontal, sede de las facultades superiores, no está completamente desarrollada, pero presenta una protuberancia similar a la del cerebro de los antropoides. El lóbulo frontal izquierdo, en particular, asociado con la capacidad del habla, no está muy desarrollado en su parte inferior, por lo que se infiere un desarrollo limitado de la facultad del habla. La fisura lateral de Silvio es relativamente ancha y abierta, y esta y otras características sugieren el cerebro del antropoide. El cerebro del cráneo de La Quina, que se cree que es femenino, también muestra muchas características primitivas.65 La capacidad cúbica absoluta del cerebro es menos significativa para la inteligencia que el desarrollo relativo de las porciones cerebrales implicadas en los procesos superiores de la mente.
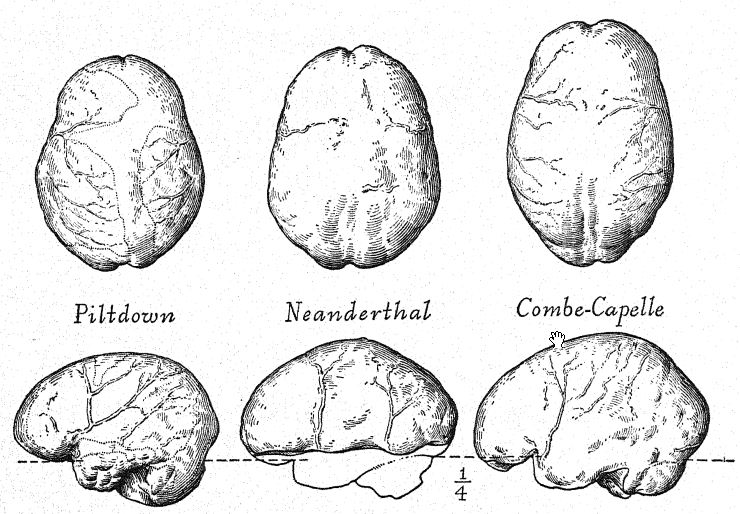
La estatura de los diversos ejemplares de la raza neandertal es estimada de forma algo diferente por Boule y por Manouvrier, y también varía según el sexo:
| Neandertal (Boule) | 1.55 m. | 5 pies | 1 pulgada |
| “ (Manouvrier) | 1.632 m. | 5 pies 4 1/5 pulg. | |
| La Chapelle (Boule) | 1.57 m. | 5 pies | 1 4/5 pulg. |
| “ (Manouvrier) | 1.611 m. | 5 pies | 3 2/5 pulg. |
| Spy (Maniobrador) | 1.633 m. | 5 pies | 4 3/10 pulgadas. |
| La Ferrassie I (Manouvrier) | 1.657 metros. | 5 pies | 5 1/5 pulg. |
| Promedio de los supuestos neandertales varones | 1.633 m. | 5 pies | 4 3/10 pulgadas |
| La Ferrassie II (hembra) | 1.482 metros. | 4 pies | 10 3/10 pulg. |
La cabeza neandertal es muy grande en proporción a su cuerpo bajo y robusto, que rara vez supera los 1,65 m en los hombres y los 1,38 m en las mujeres. Las proporciones del cuerpo y las extremidades de los neandertales arrojan una luz sorprendente sobre su historia ancestral, así como sobre sus defectos como raza dependiente de la caza. En proporción a la longitud del muslo, la parte inferior de la pierna es mucho más corta que en cualquier raza humana existente. La tibia representa solo el 76,6 % de la longitud del fémur, mientras que en las razas actuales con la tibia más corta, como los esquimales y la mayoría de las razas amarillas, nunca es inferior al 80 % de la longitud del fémur. En este aspecto, el hombre de Neandertal no es como los simios antropoides, sino que tiene una tibia relativamente más corta, ya que los gorilas tienen un índice del 80,6 por ciento, los chimpancés del 82 por ciento y los orangutanes y gibones de más del 83 por ciento; de este modo, todos los simios antropoides y las razas inferiores del hombre tienen una pierna relativamente más larga desde la rodilla hacia abajo que la de la raza Neandertal.
La brevedad de la tibia en comparación con la longitud del fémur es una prueba de que los neandertales eran muy torpes y lentos de pies, porque esta proporción es característica de todos los animales de movimientos lentos, mientras que una tibia larga y un fémur corto indican que una raza es naturalmente veloz de pies.
De modo similar, el hombre de Neandertal tiene un antebrazo muy corto, sólo el 73,8 por ciento de la parte superior del brazo; se aproxima a las proporciones observadas en los esquimales, lapones y bosquimanos.66 Aquí, nuevamente, el hombre de Neandertal se diferencia de los simios antropoides, entre los cuales el antebrazo más corto es el del gorila, con una proporción del 80 por ciento.
Existen otras características que tenderían a indicar que los ancestros de los neandertales habitaban el suelo en lugar de los árboles desde tiempos geológicos muy remotos; los brazos son mucho más cortos que las piernas, mientras que en los arborícolas son mucho más largos. Así, hemos observado en los simios antropoides que el brazo es muy largo en proporción a la pierna; en el chimpancé, que tiene los brazos relativamente más cortos entre los simios antropoides, el índice es del 104 %, es decir, los brazos son ligeramente más largos que las piernas. Por el contrario, en los neandertales la longitud del brazo es solo el 68 % de la longitud de la pierna; por lo tanto, está muy alejado del tipo de simio antropoide y se acerca más a los tipos de negro australiano y africano.
Así, para resumir las proporciones corporales de los neandertales:
Brazo corto en proporción a la pierna, índice medio 68 por ciento.
Antebrazo corto en proporción al brazo superior, índice medio 73,8 por ciento.
Hueso de la tibia corto en proporción al fémur, índice medio 76,6 por ciento.
Estatura extremadamente baja en proporción al tamaño de la cabeza.
La estructura del hombro y del tórax resulta muy interesante. Todas las costillas son notablemente robustas y de gran volumen, y, mientras que en las razas actuales presentan una sección aplanada, en los neandertales la sección presenta una forma claramente triangular. Esto implica un torso muy musculoso y robusto, en consonancia con la cabeza gigantesca y las extremidades robustas. Las clavículas son correspondientemente largas, presentando una proporción con el húmero superior al 54%, mucho mayor que la del promedio de las razas actuales; esto indica un hombro muy ancho. El omóplato también es muy diferente al de las razas superiores de humanos, e incluso al de los primates superiores; es extremadamente corto y ancho.
Si bien, como se mencionó anteriormente, el brazo de los neandertales es relativamente corto y, por lo tanto, no antropoide, presenta una mezcla de características humanas y simiescas. El húmero, o brazo superior, es verdaderamente de tipo humano, con un ángulo de torsión de 148° sobre su eje, mientras que en los simios antropoides el ángulo de torsión nunca supera los 141°. Entre los huesos del antebrazo, el más significativo es el radio, con el cual se correlaciona el movimiento de giro de la mano; la estructura de la cabeza del radio se asemeja más a la de los simios antropoides que a la de las especies humanas actuales. La estructura del otro hueso del antebrazo, el cúbito, también es muy primitiva y presenta ciertas características simiescas.
La estructura de la mano es de sumo interés en relación con la capacidad de los neandertales para fabricar herramientas. La mano es notablemente grande y robusta, comparable en tamaño a la de hombres de gran estatura en razas actuales. En cuanto a la capacidad de oposición del pulgar contra los demás dedos mediante el músculo oponente, una característica distintivamente humana, el desarrollo neandertal es claramente inferior al de las razas actuales, ya que la articulación del metacarpiano que sostiene el pulgar tiene una forma peculiar, convexa, y presenta un auténtico cóndilo convexo, mientras que en las razas humanas actuales la superficie articular de la parte superior de la articulación del pulgar tiene forma de silla de montar, es decir, cóncava desde dentro hacia atrás y convexa desde fuera hacia dentro. Así, el altamente perfeccionado [ p. 241 ] Los movimientos del pulgar del Homo sapiens no se alcanzaron en el Homo neanderthalensis. Dos falanges conservadas en el esqueleto de Chapelle-aux-Saints muestran que los dedos eran relativamente cortos y robustos.
En la estructura de la cintura escapular, nuestro hombre fósil es completamente humano; sin embargo, algunos de sus caracteres son muy primitivos y distintivos.
De modo similar, el fémur muestra varios caracteres primitivos que sólo raramente se ven en las razas actuales, como el tercer trocánter y la fuerte curvatura general hacia adelante.
La estructura de la articulación de la rodilla en relación con la tibia es muy peculiar, ya que muestra que esta siempre estaba retrovertida o doblada hacia atrás. Otras dos características de la tibia son su extrema abreviatura en comparación con el fémur y la ausencia de aplanamiento o platicnemismo. En la unión de la tibia con el tobillo (astrágalo) se observan dos facetas, que solo se conservan en las razas humanas actuales que han conservado el hábito de agacharse o la posición plegada de las extremidades; estas facetas no se encuentran en las razas que tienen el hábito de sentarse. Indican que la posición de descanso de los neandertales durante el trabajo industrial era en cuclillas, como se muestra en nuestra restauración de uno de los neandertales en Le Moustier.
Asociada a estas extremidades poderosas y de forma peculiar se encuentra la columna vertebral, particularmente corta y gruesa, cada hueso de la cual destaca por su contracción. El cuello, en particular, presenta una constitución completamente diferente a la de las razas humanas actuales. Parece que la curvatura cóncava de la espalda en los neandertales se extendía directamente hacia arriba y continuaba hasta la curvatura cóncava del cuello, como en los simios antropoides, y especialmente en el chimpancé. Las vértebras del cuello, especialmente la quinta, sexta, séptima y la primera dorsal, se asemejan mucho más a las del chimpancé que a las del europeo moderno; las apófisis espinosas se dirigen hacia atrás en lugar de hacia abajo. Esto provocó la habitual inclinación del hombre de Neandertal a la altura del cuello y los hombros, impidiéndole mantener la cabeza completamente erguida. Mientras que en la columna vertebral de las razas actuales la posición erguida se mantiene por cuatro elegantes curvaturas, dos hacia el frente y dos hacia la espalda, en los neandertales, al igual que en los miembros recién nacidos de las razas superiores, observamos sólo tres curvaturas, dos cóncavas hacia el frente, a saber, la curvatura de la espalda y el cuello, recién descrita, y una curvatura sacra o pélvica; también hay una curvatura lumbar convexa en la parte inferior de la columna vertebral neandertal, que, sin embargo, es menos pronunciada que en las especies de hombres actuales.

Resumiendo los caracteres de la columna vertebral en los neandertales, algunos de ellos son muy primitivos, como la estructura de las vértebras del cuello y el robusto desarrollo de las apófisis espinosas, la ausencia de curvatura marcada en la parte inferior de la columna vertebral y la curvatura muy suave de los huesos del sacro.
El aspecto general del hombre de Neandertal puede caracterizarse de la siguiente manera: una cabeza enorme colocada sobre un tronco corto y grueso, con extremidades muy cortas y robustas; hombros anchos y encorvados, con la cabeza y el cuello habitualmente inclinados hacia adelante en la misma curvatura que la espalda; brazos relativamente cortos en comparación con las piernas; la parte inferior de la pierna, en comparación con la superior, más corta que en cualquiera de las razas humanas existentes; la rodilla habitualmente inclinada hacia adelante sin la capacidad de enderezar la articulación ni de mantenerse completamente erguido; las manos extremadamente grandes y sin el delicado juego entre el pulgar y los dedos característico de las razas modernas; la postura en cuclillas al dedicarse a la fabricación de pedernal y otras industrias. Por lo tanto, las actitudes comunes características del Homo neanderthalensis serían bastante diferentes de las nuestras y muy desgarbadas. La cabeza pesada, el enorme desarrollo de la cara y la posición retrasada del foramen magnum, a través del cual la médula espinal se conecta con el cerebro, tenderían a arrojar la parte superior del cuerpo hacia adelante, y esta tendencia, con la menor curvatura del cuello, los hombros pesados y la forma aplanada de la cabeza, daría a esta porción del cuerpo un aspecto más o menos antropoide.

¶ Distribución geográfica de las estaciones musterienses
La raza neandertal del Musteriense estableció estaciones por toda Europa occidental, de las cuales ya se han descubierto más de cincuenta, en comparación con las cincuenta y siete o más estaciones achelenses conocidas. En algunos puntos, los antiguos campamentos abiertos de los trabajadores del sílex achelenses aún eran visitados, como a lo largo del Támesis, el Somme y el Marne. Así, Abbeville, Saint-Acheul, Montières y Chelles, en el norte de Francia, muestran una sucesión de industria musteriense tras la achelense, la chelense y, en Saint-Acheul, incluso la prechelense. Es posible que estas fueran estaciones de verano, visitadas en épocas favorables del año debido a su abundante suministro de sílex. A unas 200 millas al este de Saint-Acheul, en Bélgica, en un pequeño afluente del Mosa, se encuentra la gruta de Spy, que, junto con herramientas musterienses, ha revelado dos esqueletos fósiles humanos de la raza neandertal.
En el sur de Devonshire se encuentra la famosa caverna de Kent’s Hole, cerca de Torquay, descubierta en 1825 por MacEnery y descrita en 1840 por Godwin-Austen.68 Es interesante destacar que se han encontrado dientes de tigre dientes de sable (Machcorodus latidens) en esta caverna, lo que llevó a Boyd Dawkins a creer que este animal sobrevivió hasta tiempos geológicos tardíos: será recordado como contemporáneo de los primeros trabajadores del sílex de Chellean en Abbeville. [ p. 245 ] La vida animal de Kent’s Hole, tal como la describió originalmente Godwin-Austen, incluía restos de «elefante, rinoceronte, buey, ciervo, caballo, oso, hiena y un felino de gran tamaño», fauna que ahora se sabe que pertenece al período de la cuarta glaciación.[5]
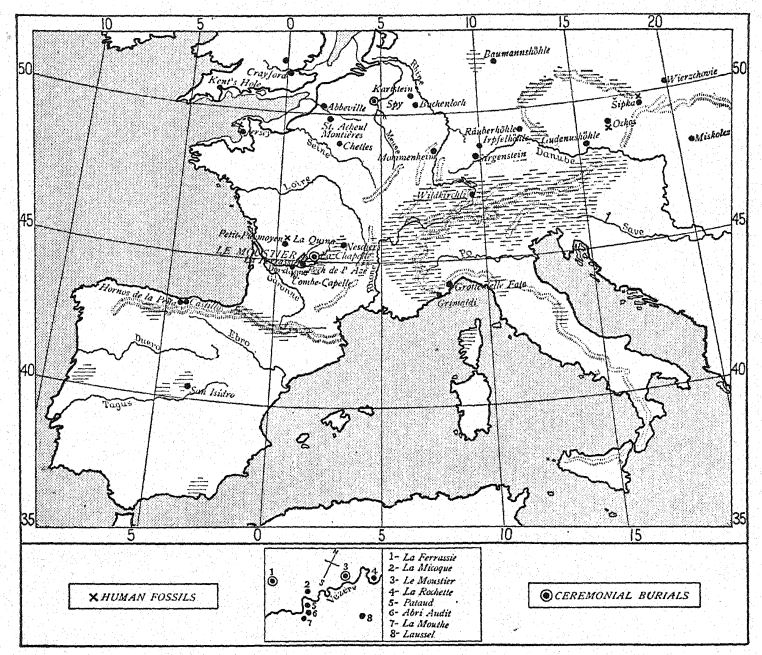
Al sur hay tres estaciones, una de las cuales, La Cotte de St. Brelade, en la actual isla de Jersey, entonces parte del continente, ha proporcionado lascas musterienses y trece dientes humanos de tipo neandertal.
Aún más al sur, en la región de Dordoña, se encuentra la estación tipo descrita en una página anterior, Le Moustier, centro de un grupo de ocho yacimientos a lo largo de las orillas norte y sur del Vézère, que se han hecho famosos por el conocimiento que aportan sobre las sucesivas etapas del desarrollo de los implementos musterienses, comenzando con la estación de cultivo primitivo de La Micoque, e incluyendo La Ferrassie, Le Moustier, La Rochette, Pataud, La Mouthe, Laussel y, finalmente, Abri Audit, que marca la etapa final en el desarrollo de la industria musteriense y, en opinión de muchos arqueólogos, su transición al Auriñaciense. En varios de estos lugares se han realizado importantes descubrimientos, tanto de fósiles humanos como de transiciones notables en el progreso de la invención. Alrededor de este grupo de Vézère se encuentran las estaciones de Petit-Puymoyèn, La Quina, donde se han hallado herramientas de la última etapa de la industria musteriense, así como un fósil humano de tipo neandertal, y La Chapelle-aux-Saints, que ha proporcionado el único esqueleto completo de un hombre neandertal descubierto hasta la fecha.

En España se encuentra la estación de San Isidro, cerca de la cabecera del Tajo, y las grutas bellamente situadas del Castillo y Hornos de la Peña, en la ladera norte de la Cordillera Cantábrica.
[ pág. 247 ]
En contraste con los numerosos yacimientos achelenses de Italia, sorprende observar que hasta la fecha solo se han descubierto dos grutas musterienses en esta región: la Grotte delle Fate, en las montañas de Liguria, y el importante conjunto de aleros de la Riviera, cerca de Mentone, conocido como las Grottes de Grimaldi, cerca de la costa y en el mismo punto donde los Alpes italianos se unen al mar. Cruzando hacia el norte, observamos la magnífica gruta suiza de Wildkirchli, en la cabecera del Rin, a 1524 metros sobre el nivel del mar.

En toda Alemania solo existen unas siete estaciones de edad musteriense incuestionable. De estas, seis son grutas, y la séptima, Mommenheim, es un redepósito fluvial de loess a lo largo de un pequeño arroyo, donde solo se ha encontrado una herramienta.69 Es interesante observar que en Alemania estos yacimientos musterienses ocupan la gran cuña territorial entre los campos de hielo escandinavos al norte y los alpinos al sur, y que Wildkirchli se encontraba de hecho dentro del área de glaciación; mientras que las cuevas de Rauberhöhle y Sipka no estaban lejos de los glaciares que cubrían los Cárpatos, y Baumannshöhle no estaba tan lejos del gran campo de hielo escandinavo. En la región de las cabeceras del Rin y el Danubio, la industria de la [ p. 248 ] Hasta la fecha, la raza neandertal solo se ha rastreado en las estaciones de Irpfelhöhle, Rauberhöhle y Sirgenstein. Esta última caverna es de especial importancia, ya que abarca toda la historia paleolítica de esta región, presentando una serie de estratos culturales sucesivos desde el Musteriense hasta la llegada de la raza neolítica. Más al este se encuentran Gudenushöhle, cerca de Krems, en la Baja Austria, y Ochos y Sipka, en Moravia, mientras que al otro lado de la frontera rusa se encuentran Wierschovie y Miskolcz. Bastante al noroeste de Wildkirchli se encuentran las estaciones de Mommenheim y Kartstein, y al norte la de Baumannshöhle.
¶ La artesanía de los neandertales
La densa vida comunitaria del Musteriense pudo haber favorecido una evolución social, el desarrollo de la imaginación y las tradiciones tribales, y el inicio de las creencias religiosas y ceremoniales, cuyos indicios aparentes se encuentran ampliamente difundidos entre las razas completamente diferentes del Paleolítico Superior. Sin embargo, esta vida no está marcada por el progreso industrial ni la invención.
Las etapas sucesivas de la industria musteriense aún no se han definido con tanta claridad como las del achelense (Schmidt70). En las estaciones y cavernas musterienses abiertas de Bélgica e Inglaterra, Schmidt ha observado las etapas del musteriense temprano, medio y tardío. Breuil y Obermaier consideran que La Micoque pertenece al final del achelense, pero marca la transición al musteriense. Breuil considera que la industria de la estación Combe-Capelle representa la cultura musteriense auténtica más antigua. Las investigaciones realizadas hasta la fecha parecen justificar las siguientes subdivisiones de la cultura musteriense en el suroeste de Francia:
6. Cultura de la auditoría de Abri, que marca la transición de la industria del Musteriense tardío a la del Auriñaciense temprano,
5. Industria Musteriense Tardía. Herramientas tipo La Quina con raspadores y yunques de hueso.
4. Industria musteriense media, con predominio de hermosas y grandes puntas musterienses cuidadosamente ‘retocadas’ en el canto y a veces en un lado, un ‘retoque’ que a veces se acerca a la técnica solutrense superior.
[ pág. 249 ]
3. Industria primitiva del Musteriense temprano, con un inventario limitado de herramientas.
2. Etapa de Combe-Capelle, con golpes de puño en forma de corazón y típicas puntas musterienses. (Llegada de renos).
1. Cultura Micoque, de transición entre el Achelense y el Musteriense. (No hay renos).
La industria del sílex, aunque muy diferente en su apariencia externa, se reconoce como una evolución directa del achelense, con la supresión o dechne de ciertos implementos y la mejora de otros. Es producto del mismo tipo de mentalidad trabajando con los mismos materiales, pero bajo diferentes condiciones climáticas y con nuevas demandas, especialmente de ropa para protegerse del clima severo. Tampoco podemos evitar la sensación de que el abandono de la vida libre y abierta de los tiempos chelenses y del achelense temprano, y el hacinamiento de las tribus neandertales bajo los refugios y en las grutas, tuvieron un efecto de empequeñecimiento tanto en el físico como en la propia industria. Los implementos musterienses, en comparación con los achelenses, impresionan como el trabajo de una raza menos musculosa y vigorosa.
Además de las numerosas y sutiles transiciones que se observan71 entre las industrias achelense y musteriense en Saint-Acheul, los descubrimientos en Laussel, en el Vézère, cerca de Les Eyzies, también aportan pruebas contundentes de una estrecha conexión entre estas culturas. Allí, amplia y profunda ante el abrigo de Laussel, se encuentra la capa musteriense, y justo debajo se encuentra una verdadera capa achelense cerca de las aguas del valle del Beune. Esto demuestra que en la época achelense este valle se profundizó al mismo grado que en la actualidad, y aquí se evidencia nuevamente una estrecha unión entre el achelense y el musteriense. En el valle del Somme, cerca de Saint-Acheul, Commont también ha observado pruebas de una estrecha conexión similar entre estas culturas. Con estos registros en el norte y el sur de Francia, la raza neandertal, conocida hacia el final del achelense y que abarca especialmente todo el período musteriense, se acerca mucho más a nosotros. Si asignamos la industria musteriense a [ p. 250 ] el último período glacial, le damos una duración de unos 30.000 años, y este es más o menos el cálculo que los anatomistas reflexivos ya han asignado para el hombre de Neandertal.
¶ Herramientas Musterienses Especiales
Dos instrumentos son especialmente típicos de la industria musteriense de principio a fin: la punta y el racloir. El primero, puntiagudo y con forma de lanza, mide de 2,5 a 10 cm de largo; el segundo es un raspador ancho, de 2,5 a 5 cm de ancho; y ambos tienen la peculiaridad de estar compuestos por una gran lasca de sílex arrancada de un bulbo o nódulo mayor, y de estar retocados solo por una cara, dejando en la opuesta la superficie concoidea lisa de la lasca. Esta punta y este raspador son muy característicos no solo de las primeras etapas, sino de la industria musteriense a lo largo de toda su trayectoria, incluyendo incluso los tipos tardíos de La Quina, y su método de fabricación es, obviamente, una modificación del descubrimiento achelense tardío de las lascas de Levallois.
0 5 Ip


Un asunto de gran interés para el desarrollo industrial de Europa occidental en aquella época es el descubrimiento de la utilización de la lasca, ya sea en las “lames de Levallois” o en el declive musteriense del coup de poing. Las lascas retocadas, de diversas formas, eran más fáciles de fabricar y reparar, y servían igualmente para desollar y desmembrar la caza, como antes se hacía con el antiguo coup de poing.
En consecuencia, el coup de poing, elaborado a partir del núcleo del nódulo, comienza a desempeñar un papel muy secundario y se presenta con poca frecuencia en los niveles musterienses. Incluso en Saint-Acheul, el centro mismo de su antiguo reinado, empezamos a encontrar formas decadentes y una mano de obra deficiente, lo que dificulta reconocer que estos son los sucesores de los coup de poing achelenses, finamente retocados. Si bien los coup de poing en la estación tipográfica de Le Moustier conservan los antiguos patrones achelenses —el ovalado, el acorazonado, el puntiagudo—, todos son de menor tamaño y presentan retoques bastante toscos. Así, tras miles de años de [ p. 252 ] desarrollo y uso, el coup de poing cae en un período de degeneración y desuso definitivo. La historia de este implemento, que hemos rastreado desde sus prototipos prechelianos, presenta una analogía sumamente interesante con el curso de la evolución observado en tantas formas animales y vegetales. Pasa por numerosas etapas de perfeccionamiento y alcanza un clímax de perfección y adaptación; luego entra en competencia con otra forma que evoluciona según un plan fundamentalmente diferente y superior, y desaparece en la lucha por la existencia gracias a la mayor utilidad del tipo que la reemplaza.
¶ Etapas sucesivas de la industria musteriense
La sucesión de etapas industriales se aprecia mejor a lo largo del río Vézère. La industria musteriense más antigua es la de Combe-Capelle, con sus coups de poing en forma de corazón, toscamente tallados, sin que se observe ninguna evidencia de una superficie preparada para el agarre manual.
En el valle del Somme, Commont74 ha observado las tres etapas siguientes en el avance de la industria musteriense:
3. Una cultura Musteriense tardía que se encuentra en las capas superiores, cerca de la cima del mismo depósito de grava, y que presenta elementos técnicos completamente nuevos. La antigua cultura del coup-de-poing ya no se valora, y todos los utensilios encontrados aquí son lascas trabajadas solo por una cara y con un retoque extraordinariamente fino.
2. Un horizonte Musteriense medio que se encuentra en las capas inferiores de un depósito de grava, perteneciente al loess más nuevo, y que contiene sólo un pequeño coup de poing,
1. Un musteriense temprano, con numerosos coup-de-poing en forma de lanza, se encuentra en la base del loess más reciente, lo que demuestra que la tradición del coup-de-poing aún perdura y que el tipo de coup-de-poing aún se conserva. Con estos se asocian los nuevos tipos de herramientas y, en especial, las puntas de mano, tan típicas de la industria musteriense.
Los niveles más recientes (2, 3) contienen lascas más largas, que ya muestran una tendencia hacia las láminas o “láminas” del Paleolítico Superior.
En los refugios y cavernas de Dordoña se observa la misma secuencia industrial, aunque la sucesión cronológica de los estratos no siempre está claramente definida. En la gruta de Combe-Capelle, los coups de poing en forma de corazón conservan con mayor fuerza las antiguas tradiciones, pero incluso aquí son superados en número por las elegantes «puntas» musterienses, lascadas solo por un lado.
El desarrollo posterior de la industria musteriense se puede observar en la estación tipográfica de Le Moustier, donde los niveles inferiores muestran un musteriense primitivo, compuesto principalmente por lascas muy finas e irregulares, transformadas en pequeños raspadores, puntas triangulares, perforadores y discos. La capa superior incluye puntas musterienses cuidadosamente trabajadas, frecuentemente retocadas por una cara en toda la superficie; aquí la técnica musteriense alcanza su máximo desarrollo, por lo que Schmidt la denomina «museriense superior».^® Por encima de esta capa, se encuentra un nivel de formas típicas del musteriense tardío, muy diferente de las pequeñas lascas primitivas del nivel inferior y similar a las formas características de La Quina, siendo el tipo dominante el racloir de La Quina, finamente tallado. Los escasos coups de poing diminutos que se encuentran en este nivel de Le Moustier constituyen la única distinción entre la industria de este nivel y la de La Quina, donde no se encuentran coups de poing. En Le Moustier también se encuentran los típicos yunques de hueso que se reconocieron por primera vez en La Quina.
La industria musteriense de los neandertales se dedicó, por lo tanto, principalmente al desarrollo de herramientas más pequeñas, en su mayoría retocadas por una sola cara, y con una mejora constante de la técnica. Sin embargo, los principales tipos de herramientas musterienses siguen siendo los mismos que en la época achelense, como se muestra en la tabla adjunta.
El instrumento conocido como punta, o punta de mano, es una forma principal y muy característica del Musteriense, perfeccionada a partir de su etapa achelense. Tiene forma de punta de lanza y está lasqueado solo por un lado, y perdura hasta finales del Musteriense, encontrándose aún en los niveles musterienses de Spy, Bélgica.
La pointe double, una forma de doble punta en forma de lanza, a veces casi alcanza la forma alargada de la pointe solutrense [ p. 254 ] de laurier, aunque nunca su esbeltez, simetría y perfección de técnica.
Existen cinco o seis variedades bien definidas del racloir, o raspador, cuidadosamente elaborado a partir de lascas. La forma principal tiene forma de medialuna, con el borde curvado hacia afuera. Otras formas son serruchadas, con bordes rectos o afilados como cuchillos. Otra forma, con bordes curvados hacia adentro de forma muy pulcra y simétrica, presenta sus bordes retocados con precisión, como si se usara para alisar mangos de hueso o madera. El barrenador también se elabora a partir de una lasca alargada y, a veces, se remata con una punta muy fina en uno de sus extremos. Cabe destacar que el grattoir, o herramienta de cepillado, tan desarrollado en las industrias del Paleolítico Superior, aparece solo esporádicamente en el Musteriense. Por ejemplo, en La Quina, en las últimas etapas de la industria musteriense, de 220 herramientas recolectadas al azar, había 166 raspadores de seis formas diferentes, 45 puntas de mano de cinco formas diferentes y 5 puntas dobles, en comparación con 5 grattatos o herramientas de cepillado. Hay muy pocas formas con forma de cuchillo. Parece que el racloir y el perj oir eran las principales herramientas empleadas en la preparación de pieles para la confección.
Es posible que en los primeros tiempos del Musteriense el coup de poing todavía fuera utilizado por los neandertales en la caza, y la fina punta de lanza y la más rara punta doble pueden haber sido desarrolladas en respuesta a las necesidades de los cazadores, que ahora se aventuraban a cazar bisontes, uros, caballos salvajes y renos.
Industrial.
- Coup de poing (decadente), piedra de mano,
- ovoide.
- en forma de corazón.
- de punta afilada.
- Hachette, helicóptero.
- Grattoir, herramienta de cepillado.
- Perforadora, taladro, perforadora.
- Couteau, cuchillo.
- Rascador, raspador,
- afilado como un cuchillo,
- borde curvado hacia afuera,
- con bordes de sierra,
- de doble filo,
- en forma de pico,
- de múltiples filos.
- Pointe, ‘punto de la mano’.
- ¿Percutor? ¿Martillo-piedra?
Guerra y persecución.
- Pointe, ‘punto de la mano’.
- Punta doble, punta de lanza ?
-Coup de poing, golpe de mano.
-Pierre de jet, tirando piedra. - Couteau, cuchillo.
[ pág. 255 ]
Las características más llamativas de todos los instrumentos que pudieron haberse utilizado en la caza son: primero, la ausencia de cualquier prueba definitiva de su fijación a un asta o mango; y segundo, la ausencia de cualquier punta con púas o cabeza. El uso de la púa, como veremos, parece ser un descubrimiento relativamente reciente de las culturas posteriores del Paleolítico Superior.
La transición del Musteriense al Auriñaciense se observa en el Abri Audit, también situado en el valle del Véfére. Aquí aún encontramos coup de poing de formas irregulares, sucesores decadentes de los tipos con forma de corazón de la industria musteriense más temprana; esta es prácticamente la última fase del declive de la antigua manufactura de coup de poing. Mientras que el coup de poing con forma de lanza del Achelense tardío nunca aparece en ninguna industria musteriense auténtica, el tipo de Combe-Capelle, más corto y con forma de corazón, atraviesa todo el Musteriense y, tras posteriores etapas de degeneración, pasa a la cultura del Abri Audit e incluso persiste en el Auriñaciense temprano. En esta última etapa, las típicas «puntas» musterienses casi desaparecen.
[ pág. 256 ]
El Musteriense, observa Schmidt,75 que conserva las tradiciones de la cultura del coup-de-poing del Paleolítico Inferior, es una de las fases más interesantes en el desarrollo de la industria paleolítica, ya que sus etapas sucesivas exhiben las últimas fases de la gran industria del coup-de-poing, de la cual solo aparecen los tipos almendrado y ovalado de raspador, y esto muy raramente, en el Auriñaciense temprano. Por otro lado, en el Musteriense tardío observamos una tendencia hacia la industria de la hoja (lámina) del Paleolítico Superior. El estudio y la observación minuciosos de las subdivisiones de la cultura Musteriense se han limitado hasta ahora al centro y sur de Francia, y aún no se han rastreado en España; pero en las grutas de Bélgica e Inglaterra se sabe que existen los tipos Musterienses temprano, medio y tardío.
Yunques de hueso, fabricados con las superficies duras de los huesos de las patas delanteras y del pie del bisonte y el caballo, fueron descubiertos en La Quina en 1906. Presentan una superficie aplanada con incisiones cruzadas demasiado regulares para ser accidentales y demasiado alejadas de la articulación como para ser resultado de un intento inexperto de seccionar la articulación. 76 Sin embargo, este no fue el único uso del hueso en la época musteriense, ya que ocasionalmente se encuentran en Dordoña herramientas puntiagudas primitivas de hueso, mezcladas con pedernales musterienses. También se encuentra una variedad de herramientas de hueso de fabricación rudimentaria en Wildkirchli, Suiza.
¶ Desaparición de la raza neandertal
Hemos visto que los neandertales habitaron Europa durante mucho tiempo, miles de años, durante los cuales sin duda experimentaron una considerable evolución de tipos inferiores a superiores, y a variedades, bajo la influencia modificadora del clima, la alimentación y los hábitos raciales. En consecuencia, los restos conocidos de neandertales muestran una marcada variación en la forma de la cabeza, así como en la dentición: algunos son más primitivos y simiescos; otros, como Spy II, se asemejan más a las razas modernas. La variedad Krapina tiene la cabeza más ancha que la variedad neandertal típica. La variedad Gibraltar es, en muchos aspectos, de tipo bajo. El individuo conocido como Spy II es de tipo superior al de los demás neandertales. Las variaciones de estatura conocidas son leves.
Por estas y otras razones, Hrdlicka,77 quien recientemente^ realizó un amplio estudio comparativo de los principales restos neandertales de Europa, opina que los neandertales evolucionaron parcialmente hacia las razas inferiores del Homo sapiens; siendo no solo en cierta medida ancestros de formas tan primitivas como la raza Brünn o Predmost del Paleolítico Superior, sino que incluso contribuyeron a la raza superior de los cromañones. También sostiene que no faltan rastros de sangre y fisonomía neandertales ni siquiera entre los europeos modernos.
En el presente volumen se plantea una perspectiva contraria: los neandertales representan una rama secundaria de la raza humana que se extinguió por completo en Europa occidental. El autor comparte esta opinión con Boule y Schwalbe. Ciertamente, la evidencia aportada por los yacimientos funerarios conocidos del Paleolítico Superior no respalda la teoría de la pervivencia de los neandertales. Sin embargo, es posible que los esqueletos del Paleolítico Superior descubiertos en Predmost, y ahora a la espera de la descripción de Maska, modifiquen esta conclusión y demuestren la teoría de Hrdlicka de que los neandertales sobrevivieron y dejaron descendientes, o hombres, de raza mixta neandertal y Homo sapiens, a lo largo del valle del Danubio.
Cualquiera que haya sido su destino en otras regiones, sin duda el cambio racial más repentino que conocemos en toda la prehistoria de Europa occidental es la desaparición de la raza neandertal al final de la etapa cultural Musteriense, que fue el último período industrial del Paleolítico Inferior, y su reemplazo por la raza cromañona. A partir de la evidencia geológica, se cree que la fecha de este reemplazo fue entre 20.000 y 25.000 años antes de nuestra era. Hasta donde sabemos actualmente, los neandertales fueron completamente eliminados; no se ha encontrado ningún rastro de la supervivencia del tipo neandertal puro en ninguno de los yacimientos funerarios del Paleolítico Superior; tampoco se han confirmado los supuestos casos de supervivencia de la cepa neandertal ni de personas con caracteres craneales neandertales. Nos inclinamos a estar de acuerdo con Boule y Schwalbe en que los supuestos casos entre las razas modernas de transmisión de caracteres neandertales son simplemente tipos bajos o reversionales, que, tras un análisis más minucioso, nunca presentan la combinación altamente distintiva y peculiar de características neandertales.
Hay razones para creer que los neandertales se estaban degenerando física e industrialmente durante las durísimas condiciones de vida de la cuarta glaciación, pero la consiguiente inferioridad y disminución numérica no explicaría su extinción total, y nos inclinamos a atribuir esto a la llegada a toda la Europa occidental neandertal, hacia finales del Paleolítico Inferior, de una nueva raza muy superior. Los arqueólogos encuentran vestigios de una nueva cultura e industria en ciertas estaciones musterienses, anteriores a la desaparición de la industria musteriense típica. Dicha mezcla se encuentra en el valle del Somme, en el norte de Francia.
De esta escasa evidencia, podemos inferir que la nueva raza compitió durante un tiempo con los neandertales antes de que los desposeyeran de sus puestos principales y los expulsaran del país o los mataran en batalla. Los neandertales, sin duda, lucharon con armas de madera y con dardos y lanzas con punta de piedra, pero no hay evidencia de que poseyeran arco y flecha. Por el contrario, existe la posibilidad de que la recién llegada raza cromañona estuviera familiarizada con el arco y la flecha, ya que una flecha con púas o una punta de lanza aparecen en dibujos de una etapa posterior de la historia cromañona, el llamado Magdaleniense. Por lo tanto, es posible, aunque está muy lejos de estar demostrado, que cuando los cromañones entraron en Europa occidental, en los albores del Paleolítico Superior, estuvieran armados con armas que, dada su inteligencia y físico superiores, les habrían dado una gran ventaja en las contiendas con los neandertales.
[ pág. 259 ]
¶ Bibliografía
(1) Commont, 1912.1, pág. 294.
(2) Smith, W., 1894.1, cap. XV.,
(3) Dietrich, 1910.1, págs. 329, 330.
(4) Penck, 1909.1.
(5) Leverett, 1910.1, págs. 306-314.
(6) Geikie, 1914.1.
(7) Op. cit., pág. 2 j 2,
(8) Op. cit., págs. 265-266.
(9) Keith, 1911.1, pág. 23, Figura 5.
(10) Munro, 19 12.1, págs. 46, 47.
(11) Lartet, 1861.1; 1875.1,
(12) De Vibraye, 1864.1.
(13) Massenat, 1868.1.
(14) Smith, W., 1894.1, cap. XIV. (is) Geikie, 1914.1, pág. 119,
(16) Smith, Wy op. cit. y págs. 196, 197.
(17) Op. cit., pág. 224.
(18) Geikie, 1914. i, pág. 118.
(19) Bachler, 19 1 2.1.
(20) Schmidt, 1912. 1, págs. 18-32, 165 171.
(21) Op. cit., Tabla opuesta a la pág. 270.
(22) Osborn, 1910.1, págs. 419, 420.
(23) Niezabitowski, 1911.1.
(24) Harlé, 1908.1, pág. 302.
(25) Obermaier, 1912. i, pág. 135.
(26) Keith, 1911.2.
(27) Boule, 1913. 1, págs.220, 221.
(28) Op. cit., pág. 64.
(29) Fischer, 1913.1, págs. 33^, 337
(30) Schaaffhausen, 1875.1; 1858.1.
(31) Lyell, 1863,1, págs. 80-92.
(32) Schwalbe, 1897.1; 1901.1; 1901.2; 1904.1.
(33) Rey, 1864.1.
(34) Cope, 1893.1.
(35) Wilser, 1898.1.
(36) Fraipont, 1887.1.
(37) Schwalbe, 1 914. 2.
(38) Dupont, 1866.1.
(39) Máscara, 1886.1.
(40) Rzehak, 1906.1.
(41) Fischer, 1913.1.
(42) Kiaatsch, 1909.1.
(43) Bouyssonie, 1.909. X.
(44) Boule, 1908.1; 1908.2; 1909.1; 1911, 1; 1912.1. :
(45) Bola, 1913.1.
(46) Martín, H., 1911.1.
(47) Nicolle, 1910.1.
(48) Keith, 19 II. I.
(49) Fischer, 1913. i, pág. 352.
(50) Schwalbe, 1914. i, pág. 544, Figs. 4 y 5.
(51) Fischer, op. cU.
(52) Boule, 1913. 1, pág. 85.
(53) Gorjanovic-Kramberger, 1909.1.
(54) Boule, 1913. 1, pág. 104.
(55) Tomos, 1914.1, págs. 588-598.
(56) Schwalbe, 1901.2; 1914.x, págs. 534, 535.
(57) Schwalbe, 1901.1.
(58) Bola, 1913. 1.
(59) op. cit., págs. 66, 67, 72, 75*
(60) Berry, 1914.1.
(61) Johnson, 1913.1.
(62) Cuartetos, 1884.1, pág. 394.
(63) Martín, R., 1914. i, pág. 645.
(64) Bola, 1910.1; 1911.1.
(65) Antonio, 1912.1.
(66) Boule, 1913. 1, pág. 119.
(67) Op. cit., pág. 120.
(68) Geikie, 1 914. 1, pág. 130; Godwin Austen, 1840,1. .
(69) Schmidt, 1912.1, págs. 23, 32, 66,
75, 76, ley, 169.
(fo) Op. cit., pág. 12S.
(71) Schuchhardt, 19x3.1, pág. 144.
(72) Dechelette, 1908.1, vol. I, págs. 98-101.
(73) Obermaier, 19 12.1, pág. 130.
(74) Commont, 1909.1.
(75) Schmidt, 1912. 1, págs. 126-128.
(76) Dechelette, 1908.1, vol. I, págs. 104, 105.
(77) Hrdlicka, 1914.1
¶ Notas al pie
Geikie denominó la cuarta glaciación completa Mecklemburgo;6 la recesión podría corresponder a su cuarta etapa interglacial, el Forestiano Inferior. Se trata del Winn de Penck en la región alpina, con un primer y un segundo máximo separados por la recesión conocida como Laufenschwankung. En América, corresponde al Wisconsin temprano con el intervalo de recesión Peoriano, seguido del Wisconsin tardío, que constituye la última gran glaciación de América. ↩︎
Obermaier, Breuil y Schmidt asignan La Micoque a la transición entre el Achelense tardío y el Musteriense temprano. ↩︎
El clima de las tundras es extremo, la temperatura en invierno cae en promedio a -2° C bajo cero, mientras que en verano la temperatura es de alrededor de 50° F. En las estepas subárticas la temperatura media de enero apenas supera los -0° C, mientras que la de julio es de 70° F. ↩︎
Los últimos ejemplares de esta raza muy primitiva de la gran isla de Tasmania se extinguieron en 1877.62 ↩︎
Esta caverna, como muchas de las descubiertas en los primeros días de la investigación antropológica, no fue explorada cuidadosamente en referencia a la importantísima estratificación horizontal de las capas de lascas de pedernal y de restos animales. ↩︎