| XXI. Le Silurien et les premiers animaux respirant de l'air | Page de titre | XXIII. L'essor des poissons et la prophétie de la domination des vertébrés |
[ p. 282 ]
Description générale. — Dans les mers et océans peu profonds actuels, et plus particulièrement dans les eaux chaudes, on trouve une grande variété d’animaux à symétrie radiale, avec ou sans squelette externe de matériaux calcaires ou calcaires. Ces animaux à l’allure florale sont les coraux et les anémones, techniquement appelés Caelenterata. Ce nom suggère une structure en forme de sac, avec une unique cavité interne servant à la fois de cavité corporelle (cœlome, caractéristique de tous les animaux supérieurs) et de sac digestif (entéron). Cette structure simple, comparable à un sac à double paroi, présente une extrémité buccale prolongée en tentacules, tandis que l’autre extrémité est fixée à des objets étrangers sur le fond marin.
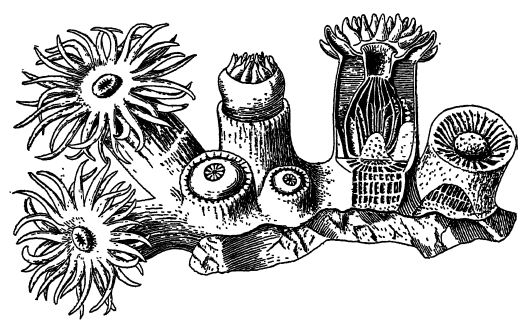
Les cœlentérés sont, en règle générale, des animaux très simples chez lesquels apparaissent les prémices d’organes définis. Dans [ p. 283 ] leur diversité, ils semblent presque épuiser les possibilités de la symétrie radiale, et beaucoup ressemblent à des fleurs. Ils ont donc également été appelés zoophytes (animaux-plantes), et leurs colonies bourgeonnantes offrent des illustrations intéressantes de coopération et de division du travail.
Certaines espèces, comme les anémones, ne sécrètent pas de squelette, mais beaucoup d’autres en possèdent un, et toutes ces espèces sont communément appelées coraux pierreux. Cependant, tous les squelettes calcaires ressemblant à des coraux ne sont en aucun cas l’œuvre de véritables coraux.

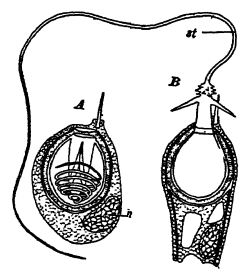
Les cœlentérés tuent par piqûre les animaux plus gros dont ils se nourrissent. Cette piqûre est effectuée par une multitude de filaments très fins, chacun étant projeté hors d’une cellule enfouie dans la peau et pénétrant dans tout corps mou entrant en contact avec les animaux (Fig. ci-contre).
La nourriture est introduite dans la bouche, puis digérée dans une cavité relativement simple du sac. Les résidus indigestes sont ensuite expulsés par la même ouverture. Ces organismes se nourrissent généralement de petits animaux, mais lorsque les individus, ou polypes, sont minuscules, leur alimentation se compose d’animaux ou de plantes microscopiques.
¶ Anthozoaires
Il existe de nombreux types de cœlentérés, mais ce chapitre ne s’intéresse qu’aux plus complexes : les anthozoaires constructeurs de roches (qui signifie « animal-fleur »). Il s’agit des coraux durs, dont beaucoup sont à l’origine de récifs et de roches. Les animaux qui construisent les récifs durs sont généralement très petits (moins de 6 mm), bien que certaines espèces atteignent un diamètre de 2,5 cm ou plus. Les squelettes coloniaux peuvent atteindre jusqu’à 4,5 m de diamètre et de hauteur.
La cavité interne contient un œsophage digestif tubulaire rudimentaire, qui s’ouvre vers le haut dans une bouche allongée et vers le bas dans la véritable cavité digestive ou codentéron [ p. 284 ] (voir Fig., p. 282). La paroi de cette dernière présente de nombreux replis longitudinaux ou invaginations de la paroi corporelle qui s’enfoncent plus ou moins profondément dans la cavité. Lorsqu’un de ces polypes est coupé transversalement, les replis ou cloisons apparaissent comme des rayons, d’où leur autre nom d’animaux rayonnants (Fig., p. 282). Lorsqu’un squelette de soutien rigide ou une partie pierreuse, appelée corallum, est présent, comme c’est le plus souvent le cas, il est généralement composé de carbonate de calcium (voir Pt. I, p. 182), est presque toujours sécrété par la couche externe du corps mou et possède sa forme précise. Par conséquent, les squelettes des coraux ont une structure rayonnée ou cloisonnée, et seul ce squelette calcaire est conservé dans les fossiles (voir Fig., ci-dessous). Les polypes vivent soit isolément et sont alors de forme cylindrique ou conique (coraux en coupe), soit, plus fréquemment, forment des colonies par bourgeonnement d’individus très proches.

Le squelette corallien. — Les coraux en coupe, qui sont les bases calcaires de polypes isolés, sont très fréquents dans les roches du Paléozoïque. Ils se multiplient par reproduction sexuée et non par bourgeonnement, comme c’est le cas pour les formes coloniales. Les coraux en forme de coupe sont généralement coniques et plus ou moins incurvés, avec des coupes de profondeur variable divisées en de nombreux compartiments peu profonds par des cloisons radiales appelées septa (voir Fig. ci-dessus). Ces septa prennent naissance dans la paroi externe et sont de longueur variable, les différentes longueurs étant disposées [ p. 285 ] de manière régulière ; seule une partie d’entre eux, généralement les septa primaires, se rejoignent au centre, où ils forment souvent une colonne simple ou torsadée, la columelle. Les coraux coloniaux ont une structure générale similaire à celle des polypes isolés, mais lorsqu’ils sont très serrés, les corallites prennent une forme prismatique (voir Fig. p. 284). Ils peuvent également avoir une forme circulaire, les espaces entre les individus étant alors remplis d’une structure lâche.
Les types de structures coralliennes les plus fréquemment observés à l’état fossile sont les suivants :
Les coraux tabulés (du grec « tabula », signifiant « table », en référence aux nombreuses cloisons transversales présentes dans les tubes) sont des coraux paléozoïques primitifs mais spécialisés. Ils se développent toujours en colonies et jamais individuellement, ce qui explique l’absence de coraux en forme de coupe parmi les Tabulata. En règle générale, les polypes individuels sont petits, voire minuscules, transversalement, mais ils construisent longitudinalement des tubes plus ou moins longs, abondamment cloisonnés par des tabules et perforés de pores à mesure que les animaux grandissent (voir Fig. ci-dessus). La croissance des polypes peut être lâche lorsque les corallites individuels sont ronds ou ovales, mais le plus souvent, les tubes sont étroitement accolés et les corallites uniformément prismatiques. On n’observe pas de septa radiaux, et lorsqu’ils semblent présents, il s’agit d’une série de courtes épines disposées en rangées longitudinales (des septa naissants, en quelque sorte), ou bien les épines peuvent être disposées de manière irrégulière. Les Tabulata sont donc des coraux paléozoïques dépourvus de véritables cloisons, et se distinguent ainsi aisément de leurs proches parents, les Tetracoralla. Les parois des corallites sont minces et perforées de larges pores, les pores muraux ou pariétaux (voir Fig. B ci-dessus), qui correspondent à des échecs de bourgeonnement, l’espace étant quasiment insuffisant pour permettre à la grande majorité des bourgeons potentiels de se développer en individus.
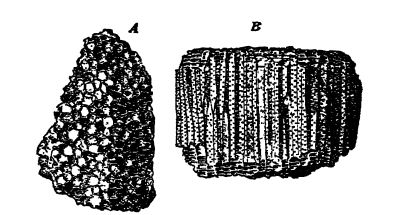
Les coraux tabulés sont apparus au Champlainien moyen et ont connu leur plus grand développement au Silurien et au [ p. 286 ] Dévonien. Le corail en chaîne (Halysites, Pl., p. 270, Fig. 1) était caractéristique du Silurien et du Champlainien, tandis que les coraux alvéolés (Favosites, Fig., p. 285) étaient d’importants constructeurs de récifs au Silurien et au Dévonien, certaines colonies atteignant un diamètre de 1,2 mètre. Le corail en forme d’orgue (Syringopora, Pl., p. 320, Fig. 8) était commun tout au long du Silurien et pendant la majeure partie du Paléozoïque ultérieur.
Les tétracoraux (coraux à septa disposés en quatre quadrants ou bimdles) sont des fossiles courants tout au long du Paléozoïque, depuis le Champlainien moyen, et plus particulièrement à partir du Silurien (voir fig., p. 284, et 4-7, p. 320). Ils se présentent sous forme de corallites isolés, de coraux en forme de coupe, ou en colonies composées. Les coraux en forme de coupe étaient très communs au Paléozoïque moyen et étaient plus nombreux, plus variés et de plus grande taille (jusqu’à 60 cm de long) que ceux des périodes ultérieures. Les tétracoraux composés étaient également abondants et, avec les coraux en forme de coupe, largement répandus, notamment comme constructeurs de récifs au Silmien et au Dévonien. Ils étaient rares au Pennsylvanien et ont disparu avec le Permien.
Chez les tétracoraux en forme de coupe, le regroupement des septa est généralement bien visible, et leur revêtement extérieur lisse ou légèrement ridé (épithèque) s’étendant de la base jusqu’au bord de la coupe permet de les distinguer aisément des autres types. Les tétracoraux composés sont généralement plus difficiles à différencier des hexacoraux (coraux à six rayons), car le tétramérisme, ou division en quatre parties, est fortement ou totalement masqué. Cependant, on observe généralement que les polypes sont plus grands tandis que les septa radiaux sont plus nombreux, ou du moins plus fins, plus droits et plus simples. Certains de ces tétracoraux composés atteignent la taille des colonies modernes et ont été observés jusqu’à 3,6 mètres de diamètre.
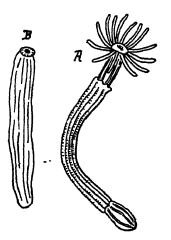
Le zoanthaire le plus primitif des mers actuelles est Edwardsia, et l’on sait que les hexacoraux actuels passent par un stade de développement similaire à celui d’Edwardsia. Edwardsia est un animal à corps mou, c’est-à-dire dépourvu de squelette, et vit dans les eaux froides. Au Cambrien moyen, près de Field, en Colombie-Britannique, Walcott a découvert un fossile très semblable à Edwardsia, également sans squelette calcaire. Il l’a identifié comme Mackenzia costalis, le considérant toutefois comme un holothurie. Raymond suggère [ p. 287 ] que Mackenzia pourrait être étroitement apparenté à Edwardsia, et que des formes similaires pourraient avoir donné naissance, au Champlainien, aux Tetracoralla sécrétant du calcaire, puis, plus tard au Trias, aux coraux pierreux, ou Hexacoralla. (Voir Fig., p. 256.)
Les hexacoraux étaient les plus communs après le Paléozoïque et constituent les principaux constructeurs de récifs des océans actuels. Leur nom fait référence au fait que les jeunes polypes possèdent six septa primaires et que tous les cycles suivants de cloisons s’insèrent régulièrement entre ces septa primaires. Il existe donc une disposition radiale régulière des septa, qui ne sont jamais regroupés en quadrants comme chez les tétracoraux, et les polypes isolés ne présentent pas non plus une forme aussi nettement en coupe ou en entonnoir. Chez les formes composées, les animaux sont généralement petits, souvent très petits, comme chez les coraux corne de cerf, tandis que les corallites ne sont généralement pas aussi nettement séparés les uns des autres que chez les coraux plus âgés. Les parois apparaissent souvent épaisses, et chez de nombreuses formes, les individus sont comprimés et semblent fusionner irrégulièrement les uns avec les autres, évoquant les circonvolutions d’un cerveau de mammifère, d’où le nom de coraux cérébraux, qui leur est parfois attribué. Voir Pt. I, p. 183.
Habitats des coraux vivants. — Les hexacoraux vivent dans tous les océans, de la surface jusqu’à 3 350 mètres de profondeur, mais sont particulièrement communs dans les eaux tropicales claires et à des profondeurs inférieures à 73 mètres. Dans les eaux plus profondes, plus sombres et plus froides, les polypes simples prédominent, tandis que sur les récifs où la lumière du soleil est intense, les coraux composés sont les plus prolifiques. On trouve cinq fois plus d’espèces d’hexacoraux dans les eaux de moins de 45 mètres de profondeur qu’entre cette profondeur et 73 mètres, la profondeur maximale à laquelle toute espèce récifale peut se développer. Entre 180 et 730 mètres, on trouve une autre zone corallienne, généralement composée de petits polypes simples ou de formes composées délicates et ramifiées. Au-delà de cette profondeur, les coraux sont peu nombreux et peu variés, et ceux qui existent sont petits et extrêmement fragiles. Au large de la côte est de l’Amérique du Nord, entre le cap Hatteras et Terre-Neuve, vivent quatorze espèces de coraux, dont plusieurs descendent à plus de 1 800 mètres de profondeur (Verrill). Les coraux hémisphériques peuvent atteindre 4,5 mètres de diamètre, tandis que les colonies de coraux corne de cerf peuvent atteindre 4,5 mètres de hauteur.
Concernant la température, les coraux strictement constructeurs de récifs ne prospèrent qu’entre 20 et 26 °C, mais peuvent survivre jusqu’à 29 °C. La zone corallienne plus profonde présente une température bien plus basse, entre 4 et 10 °C, et les espèces abyssales peuvent vivre dans une eau aussi froide que 0 °C. En d’autres termes, les coraux peuvent vivre à n’importe quelle profondeur, mais les coraux constructeurs de récifs ne vivent que dans des eaux chaudes et peu profondes
[ p. 288 ]
Il y a probablement aujourd’hui plus de récifs coralliens qu’à la plupart des époques de l’histoire de la Terre, même s’ils sont désormais tous confinés aux océans plus chauds. On estime qu’environ un vingtième des mers continentales (500 000 milles carrés) sont actuellement recouvertes de récifs coralliens. Cet excédent semble être dû à la récente élévation du niveau océanique, conséquence de l’augmentation du volume d’eau résultant de la fonte des calottes glaciaires polaires. Avec l’élévation du niveau océanique, les zones inférieures des récifs coralliens deviennent des habitats plus favorables et plus vastes. Concernant la nature des récifs coralliens et leurs lieux de croissance, voir les pages 182 à 190 de la partie I. Pour les récifs coralliens fossiles, voir les chapitres sur le Silurien et le Dévonien.
¶ Lectures complémentaires
J. D. Dana, Corals and Coral Islands, 1872 ;
W. Saville-Kent, The Great Barrier Reef of Australia. 1893.
T. W. Vaughan, « Quelques coraux d’eaux peu profondes de l’île Murray (Australie), des îles Coeos-Keeling et de l’île Fanning ». Publications du Département de biologie marine, Carnegie Institution of Washington, vol. 9, 1918, p. 51-210. (Voir pour de superbes photographies de coraux vivants et pour une liste d’autres articles sur les coraux du même auteur.)
Coraux du Paléozoïque :
W. J. Davis, « Coraux fossiles du Kentucky ». Kentucky Geological Survey, 1876. James Hall, « Fossiles de coraux du Dévonien ». New York Geological Survey, 1876. Également dans Paléontologie de New York, vol. 6, 1887.
P. E. Raymond, « Histoire des coraux et des océans sans calcaire ». American Journal of Science, 5e série, vol. 2, 1921, p. 343-347.
C. A. Romingeb, « Coraux fossiles ». Étude géologique du Michigan, vol. 3, 1876.
| XXI. Le Silurien et les premiers animaux respirant de l'air | Page de titre | XXIII. L'essor des poissons et la prophétie de la domination des vertébrés |