[ p. 289 ]
Dans ce chapitre, nous étudierons les poissons, leurs espèces, leur ascendance et comment leurs descendants, plus adaptables, ont quitté leurs habitats aquatiques ancestraux pour la terre ferme. C’est une étude des plus fascinantes, car l’humanité entière s’intéresse soit à la grande valeur économique des poissons, soit aux agréables moments que leur pêche procure au pêcheur ; mais elle revêt un intérêt particulier pour le philosophe qui voit dans les poissons une étape sur le chemin de l’évolution, menant des invertébrés vermiformes au sommet de l’autonomie organique chez l’homme raisonnant.
Les poissons furent la première classe de vertébrés à connaître un succès notable. Dotés d’une colonne vertébrale et d’un système nerveux central, et grâce à leur vigilance, leur mobilité et leur fécondité supérieures, ils surpassent aisément les invertébrés. Ils sont aussi parfaitement adaptés à l’eau que les oiseaux le sont à l’air. En règle générale, ils vivent soit exclusivement dans les mers et les océans, soit exclusivement dans les eaux douces terrestres. Cependant, de nombreux poissons marins passent une partie de leur vie en eau douce ; certains se nourrissent et se reproduisent aussi bien en eau douce qu’en eau de mer ; d’autres se nourrissent en mer et se reproduisent en rivière, ou, plus rarement, comme les anguilles, migrent des rivières vers [ p. 290 ] la mer pour se reproduire. Parmi les poissons marins remontant les rivières pour se reproduire, certains y sont restés volontairement, d’autres se sont retrouvés confinés à terre suite à la formation de barrières terrestres, et ont évolué en races ou espèces distinctes. Grâce à la capture des poissons dans les cours d’eau et à la dispersion de leurs œufs par les oiseaux échassiers, les poissons d’eau douce ont atteint leur large répartition actuelle sur les continents.
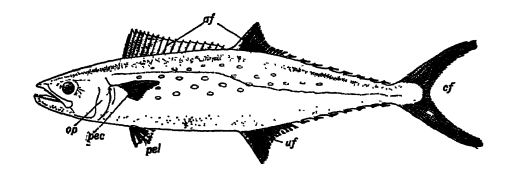
Caractéristiques distinctives. — Les poissons sont des vertébrés dépourvus de cou. Ils n’ont pas d’oreilles et donc pas de tympan. Leur température corporelle est sensiblement la même que celle du milieu dans lequel ils vivent ; on dit donc qu’ils sont à sang froid. Du fait de leur habitat aquatique, tous les poissons possèdent des branchies et des nageoires (voir Fig., p. 289). Les branchies sont situées de chaque côté de la tête et sont constituées de filaments creux et délicats dans lesquels circule le sang. L’eau est absorbée par la bouche et rejetée sur ces organes respiratoires, où le sang contenu dans les branchies extrait l’oxygène libre dissous. La bouche est généralement pourvue de dents. Une vessie natatoire est généralement présente et peut servir d’organe hydrostatique ou de flotteur ; chez certaines espèces, elle est transformée en un sac cellulaire qui fait office de poumon et assiste les poissons dans leur respiration.
La peau est soit douce et nue, soit, plus fréquemment, protégée par le développement d’épines, de mosaïques de denticules, d’écailles imbriquées ou de plaques osseuses appelées scutelles. Le principal organe de locomotion est la puissante queue. Celle-ci est assistée par les nageoires paires, dont la paire antérieure, appelée membres pectoraux, se situe juste derrière les nageoires dorsales, et la paire postérieure, ou membres pelviens, est placée tout en arrière (voir Fig., p. 289). Les nageoires paires des poissons correspondent aux membres antérieurs et postérieurs des vertébrés supérieurs. Leur squelette, cependant, ne peut être facilement comparé à la structure des membres des autres vertébrés. La façon dont ces nageoires sont devenues des membres est abordée plus loin dans ce chapitre. Les poissons possèdent également généralement un nombre variable de nageoires médianes impaires qui servent principalement d’organes d’équilibre. Les nageoires dorsales sont appelées nageoires dorsales, celles situées sur la face ventrale entre l’anus et la queue sont appelées nageoires anales, et la queue constitue la nageoire caudale (voir Fig., p. 289). Toutes les nageoires sont soutenues par des barres ou rayons osseux, appelés rayons de nageoire.
Le squelette peut être entièrement cartilagineux, bien qu’il soit généralement plus ou moins osseux. Le squelette principal est constitué de la colonne vertébrale. Les os de la tête sont de deux types : les os cutanés, comme les écailles, et les os de la boîte crânienne.
Types de queues de poissons. — La queue présente trois types généraux. (1) La colonne vertébrale se prolonge par des articulations de taille décroissante presque jusqu’à l’extrémité de l’animal et est entourée d’une nageoire caudale verticale monolobée. C’est le type le plus primitif, très répandu chez les poissons du Dévonien et présent [ p. 292 ] chez les juvéniles [ p. 291 ] de la plupart des poissons actuels ; il est connu sous le nom de queue diphyrique ou symétrique (voir Pl., p. 295, Fig. 4). (2) Chez d’autres types, la colonne vertébrale se courbe vers le haut et se termine par des articulations de taille décroissante dans le lobe supérieur, plus grand. Ce type de queue est donc rarement symétrique et est appelé hétérocerque en raison de l’inégalité des lobes (voir Pl., p. 291, Fig. 2, 4 ; Pl., p. 295, Fig. 1, 2). Elle est fréquente chez les requins actuels et chez la plupart des poissons du Paléozoïque. (3) Les poissons osseux modernes possèdent également une queue symétrique, mais bien que la colonne vertébrale se courbe ici vers le haut, elle se termine abruptement, et à partir des articulations terminales, les rayons des nageoires se développent de manière à former une queue symétrique. Ce type d’appendice caudal est appelé homocéréal (voir Fig. ci-dessous).
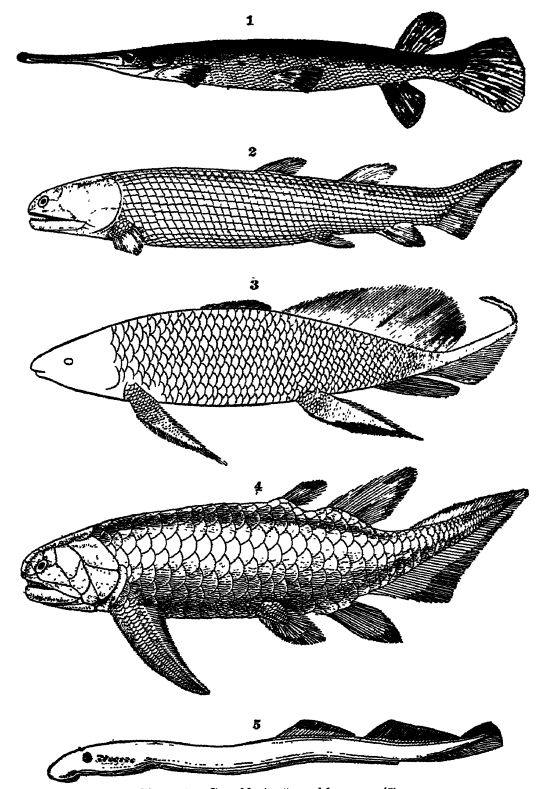
Fig. 1, brochet américain vivant (Lepidosietis osseii), x 2, ganoïde dévonien d'Écosse (Osteolepis macrolepidotui), x 1 ; 3, ganoïde du Dévonien supérieur de la baie de Scaumenac, Canada (Scaumenacia curia), x 4, ganoïde dévonien à écailles δεδ, d'Écosse (Floopichius flemingi), x 1 ; 5, lamproie marine (Pefromyzon marinus), x 1. Remarquez les sept ouvertures à l'arrière de l'œil, qui sortent des branchies ; ainsi que le bord de la bouche circulaire et l'absence de nageoires paires.
Fig. 2 et 4 d'après Woodward, British Museum Guide Book ; Fig. 3 d'après Hussakof.
Alimentation des poissons. — Parmi les poissons, certains se nourrissent exclusivement de plantes, d’autres de plantes et d’animaux, d’autres encore exclusivement d’animaux, et d’autres enfin de vase où vivent de minuscules plantes et animaux. La majorité des poissons, cependant, se nourrissent d’autres poissons, sans distinction d’espèce, et dévorent leurs propres petits aussi facilement que ceux de n’importe quelle autre espèce. Les dents à pointe émoussée servent généralement à broyer les coquilles ; les dents à bords dentelés indiquent souvent un régime herbivore ; tandis que de fortes incisives peuvent indiquer un régime alimentaire composé principalement de nageoires et de crustacés. En général, plus les dents sont pointues et la bouche grande, plus l’espèce est clairement piscivore. Les poissons à petite bouche sont souvent herbivores ou vaseux.
Croissance. — Les poissons grandissent tout au long de leur vie et leur taille dépend de leurs ancêtres ainsi que de la quantité de nourriture qu’ils peuvent se procurer : la durée de vie est variable ; certains vivent de nombreuses années, d’autres ont une courte durée de vie et certains semblent être annuels.
Origine des poissons. — Bien que la plupart des poissons possèdent un squelette osseux et que des animaux ressemblant à des poissons soient connus depuis le Champlainien moyen, leurs ancêtres fossiles les plus primitifs restent encore inconnus des paléontologues. Les zoologistes sont également incapables de déterminer leur ascendance parmi la vaste diversité des formes vivantes et leurs embryons. Tout ce que les paléontologues peuvent affirmer, [ p. 293 ] c’est que les plus anciens vertébrés fossiles sont sans aucun doute des poissons, vraisemblablement des élasmobranches et des ostracodermes. Dans ces conditions, nous laisserons cette question aux découvertes futures et passerons sans plus tarder à la description des différentes espèces de poissons.
Classification des poissons. — Pour les classifications actuelles, les poissons peuvent être divisés comme suit :
- Classe Poissons ou vrais poissons.
- Sous-classe des Élasmobranches ou poissons-cartilages
- Ordre des Acanthodei ou requins épineux
- Ordre des Selachii, ou vrais requins et raies
- Sous-classe des Ostracodermes ou requins aberrants éteints
- Sous-classe Ganoidei ou poissons à opercule émaillé
- Sous-classe des Téléostéens ou poissons osseux
- Sous-classe des dipneustes ou vrais dipneustes
- Sous-classe Arthrodira ou poissons cuirassés
- Sous-classe des Élasmobranches ou poissons-cartilages
¶ Sous-classe des Élasmobranches ou Poissons Gristle
Le terme Elasmobranchii signifie « possédant des branchies en forme de plaques », et ce groupe comprend les requins anciens et modernes, les poissons-scies, les chimères, les raies et les torpilles. À quelques exceptions près, les espèces actuelles vivent en milieu marin (voir Pl., p. 295, Fig. 3). Ce sont les poissons les plus primitifs, et ils sont à l’origine des espèces supérieures dès les premiers stades du Paléozoïque.
Chez la plupart des élasmobranches, la bouche se situe sur la face ventrale de la tête, mais chez certains, elle se trouve à l’extrémité antérieure. Le squelette interne est composé de cartilage ou de tissu granuleux, plus ou moins calcifié. La queue est généralement hétérocerque. La peau paraît souvent nue, mais elle présente toujours des denticules dermiques (petites écailles) à l’extrémité d’un émail dur et brillant, à cœur de dentine (phosphate et carbonate de calcium fluoré) et à base osseuse (phosphate de calcium) enfoncée dans les couches profondes de la peau. Ce type d’écaille est dit placoïde et possède la même structure que les dents, qui se sont également développées à partir de la peau. Lorsque les denticules sont petits et serrés, ils forment ce que l’on appelle le « shagreen » (signifiant mosaïque rugueuse).
Chez les poissons cartilagineux actuels, les cinq fentes branchiales situées de chaque côté du corps, à l’arrière de la tête, sont soit ouvertes, soit recouvertes d’un repli cutané ; elles ne sont jamais fermées par un opercule osseux comme chez la plupart des autres espèces. Les nageoires médianes sont souvent pourvues d’épines longues, pointues, humifères ou osseuses, notamment les dorsales. Ce sont ces épines et les dents des requins qui sont généralement conservées à l’état fossile ; malheureusement, elles ne fournissent que peu d’informations sur les caractéristiques générales de ces animaux.
Le cerveau est enfermé dans une boîte cartilagineuse solide, ou crâne, sans sutures ni articulations ; en cela, les élasmobranches [ p. 294 ] se distinguent de presque tous les autres poissons (à de rares exceptions près chez les ganoïdes), chez lesquels les os du crâne sont facilement identifiables. Ils ne possèdent pas de vessie natatoire.
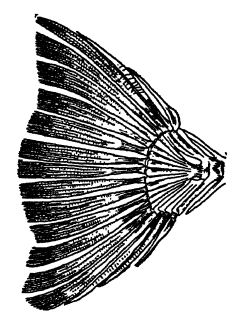
Les requins acanthodiens, ou requins épineux, sont petits et les plus primitifs des vrais requins. Ils apparaissent à la fin du Silurien, sont particulièrement communs dans le grès rouge d’Écosse et dépassent rarement 30 cm de longueur (voir Pl., p. 295, Fig. 1). Ils se distinguent des vrais requins et de la plupart des autres requins du Paléozoïque par leur peau écailleuse, dans laquelle sont enchâssées de petites plaques osseuses aplaties et étroitement imbriquées, composées de dentine et d’émail, et par le fait que leurs robustes épines ne se limitent pas aux nageoires médianes mais se développent également au niveau des nageoires paires. Leurs dents sont petites et plutôt pointues, comme celles des requins modernes. Les requins acanthodiens sont, directement ou indirectement, les ancêtres des requins plus récents et des ganoïdes supérieurs.
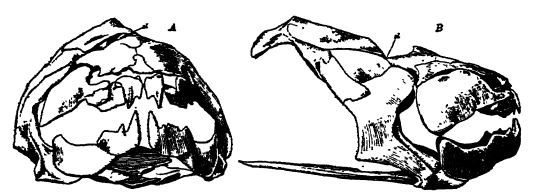
Les requins conchylivores étaient particulièrement bien représentés au Paléozoïque supérieur, par deux familles connues sous le nom de cestraciontes et de cochliodontes. Le premier groupe est aujourd’hui le mieux représenté par le requin dormeur de Port Jackson, qui vit au large de l’Australie-Méridionale (voir Pl., p. 295, Fig. 3). Il se caractérise par deux épines dorsales robustes et, dans la partie profonde de la bouche, par de nombreuses dents émoussées et papilleuses, adaptées au broyage des coquillages et des crustacés (d’où son surnom de « broyeur d’huîtres »). Chez les requins cochliodontes, les dents individuelles sont fusionnées en deux grandes plaques dont la surface supérieure présente des crêtes spiralées évoquant les spirales des coquilles d’escargots, caractéristique qui a donné naissance au nom de cette famille (voir Fig. A, p. 342). Ce sont ces parties dures qui sont si fréquentes au Paléozoïque supérieur et beaucoup plus rares au Mésozoïque. Les cochliodontes, cependant, ont disparu avec le Carbonifère.
Les requins modernes sont les véritables Selachii (terme grec signifiant requins). Cet ordre comprend les grands requins actifs, plus ou moins fusiformes, les poissons-scies, ainsi que les raies et les torpilles aplaties et lentes, vivant sur le fond marin, de l’époque actuelle et des ères Cénozoïque et Mésozoïque. Ce type de requin, doté de dents acérées, a été abondant. Dans le Miocène de Caroline du Sud, on a retrouvé des dents de ce type, dont les plus grandes mesuraient près de 15 cm (Carcharodon), ce qui indique l’existence de requins atteignant une longueur de 18 mètres. Le grand requin bleu actuel atteint une longueur de 12 mètres.
Occupance géologique. — En Amérique, les plus anciens restes de requins sont de petites épines de nageoires, apparemment d’espèces acanthodiennes (Pl., p. 295, Fig. 1), découvertes dans le Silurien supérieur (Cayugan et non Clinton comme indiqué initialement) de Pennsylvanie. Au Dévonien moyen, les épines, les dents et les denticules cutanés de petits et grands requins deviennent courants. Au Mississippien, les formes conchylivores étaient très diversifiées (voir p. 341), mais perdent leur prédominance au Pennsylvanien et disparaissent pratiquement avec le Paléozoïque. Le type de dent coupante, si répandu chez les requins actuels, est rare au Paléozoïque, plus fréquent au début du Mésozoïque, mais ne devient courant qu’au Crétacé.
¶ Ostracodermes de sous-classe ou requins aberrants
Les ostracodermes sont des poissons fortement cuirassés, probablement un groupe de requins cartilagineux très aberrants. Cette sous-classe comprend les plus anciens poissons connus, apparus au Champlamien moyen et disparus au Dévonien [ p. 296 ] supérieur. Ils représentent une lignée de l’évolution des poissons dans la mauvaise direction, car ils n’ont laissé aucune descendance. Ils sont, en fait, si différents des autres vrais poissons qu’il est impossible, à ce jour, de déterminer leur lien de parenté avec les autres vertébrés. Les ostracodermes étaient petits, ne dépassant pas 18 cm, probablement benthiques et d’un tempérament lent, vivant apparemment principalement dans les lagunes marines et les rivières. La tête et la partie antérieure du corps étaient grandes, larges et aplaties, et recouvertes de petites plaques ou écailles placoïdes, parfois fusionnées en plaques osseuses plus ou moins épaisses, tandis que la région postérieure était soit nue, soit plus ou moins recouverte de plaques ou d’écailles similaires. La queue était hétérocerque et il n’y avait pas de squelette interne (voir Figs., ci-dessus, et p. 297).

Fig. 1, Lower Devonian acanthodian shark, from Scotland (Climatius X i: 2. Upper Devonian shark, from Ohio (Cladoselache fyleri), x J; 3, livin Port Jackson cestraciont shark, from Austraha (Cestracion philippi) x 1/10; livin lung-fish of Australia (Neocerotodus fosters) X A; 5, Devonian lung-fish,

La structure et la forme des poissons à peau osseuse présentaient une grande variété. Les espèces les plus anciennes, de construction très diverse, étaient dépourvues de nageoires latérales ou d’appendices natatoires (Pteraspis, Cephalaspis, Drepanaspis_), et leurs yeux étaient soit très écartés, soit très rapprochés (voir Fig., p. 297). Les formes à nageoires antérieures blindées, à armure marquée sur la région antérieure et aux yeux rapprochés (Bothriolepis, Pterichthys, autrefois considérés comme des coléoptères géants, voir Fig. ci-dessus) étaient [ p. 297 ] spécifiques au Dévonien supérieur. Il est possible, cependant, que ces derniers genres — les « poissons ailés » de Hugh Miller, qui remarqua que sa première impression fut d’avoir trouvé en eux « le lien entre les tortues et les poissons » — n’appartiennent pas à cette sous-classe. Les oétracodermes sont largement répandus en Europe et en Amérique du Nord.
¶ Sous-classe Ganoidei ou Poissons à écailles émaillées
Le terme Ganoidei, qui signifie « aspect brillant », fait référence à la surface luisante de la plupart de ces poissons, comme par exemple chez les lépisostés actuels (voir Pl., p. 291, Fig. 1). Cette brillance est due aux écailles rhombiques plus ou moins épaisses des lépisostés ou aux écailles cycloïdes des formes dévoniennes, recouvertes d’un émail brillant semblable à celui des dents. Les écailles des ganoïdes diffèrent de celles des placoïdes des élasmobranches par le fait qu’elles ne possèdent que deux couches : un émail externe et une couche osseuse sous-jacente plus épaisse. Certains ganoïdes présentent cependant de véritables écailles cycloïdes élastiques, semblables à celles que l’on trouve chez les poissons consommés, c’est-à-dire de fines plaques circulaires de phosphate [ p. 298 ] de calcium, mais sans émail (voir Pl., p. 291, Fig. 4). Les ganoïdes sont essentiellement des poissons d’eau douce, bien que certains soient marins et ne remontent les rivières que pour frayer. Dans le passé géologique, les espèces marines semblent avoir été considérablement plus répandues qu’aujourd’hui. Les exemples vivants les plus connus sont les esturgeons et les lépisostés, mais l’origine de cette sous-classe remonte au moins au Dévonien inférieur, car elles constituent la moitié de la faune piscicole connue du Vieux Grès Rouge (voir Pl., p. 291, Fig. 2-4) et restent abondantes jusqu’au début du Crétacé. Par leur organisation, elles forment un lien entre les requins acanthodiens et les poissons osseux.

Le crâne est bien protégé par des os dermiques ou est complètement ossifié, et bien que le squelette interne soit plus ou moins osseux, il existe chez certaines formes un squelette entièrement cartilagineux. C’est dans cette sous-classe qu’apparaît pour la première fois la bouche de poisson parfaite, où la mâchoire inférieure s’articule avec les dents de la mâchoire supérieure, elle aussi parfaite, comme chez les vertébrés supérieurs. Une vessie natatoire est toujours présente et reliée à l’œsophage ; chez les lépisostés, elle participe à la respiration mais n’est jamais cellulaire comme chez les dipneustes. L’intestin possède une valvule spirale, et la queue est hétérocerque ou diphycerque.
Ganoïdes à nageoires frangées. — Selon la morphologie de leurs nageoires, les ganoïdes se divisent en deux ordres : (1) les ganoïdes à nageoires frangées ou lobées (Crossopterygii) et (2) les ganoïdes à nageoires en piliers (Stylopterygii). Chez ces derniers, les membres ne présentent pas les lobes épais, musclés et recouverts d’écailles des Crossopterygii, mais ressemblent davantage à ceux des poissons consommés. Le premier ordre ne compte plus que deux genres, tous deux vivant en eau douce en Afrique. Bien que leur vessie natatoire leur serve également de poumon, ces poissons ne peuvent survivre hors de l’eau plus de trois heures. Ils étaient communs au Dévonien inférieur, les espèces les plus connues étant les genres Holoptychius, Eusthenopteron et Osieolepis (Pl., p. 291, Fig. 2-4). Chez ces ganoïdes, les couches osseuses des dents coniques sont souvent profondément et complexement plissées ou labyrinthiques, ce qui est particulièrement intéressant car le même type de dents se retrouve chez tous les amphibiens du Paléozoïque. Cette similarité de structure dentaire est un autre indice suggérant l’origine probable des Amphibiens (ordre des Stégocéphales) chez ces ganoïdes d’eau douce à nageoires digitiformes du Dévonien.
¶ Sous-classe Dipnoi ou vrais dipneustes
Les dipneustes (du grec me aning, « double respiration ») sont ainsi nommés car les trois genres actuels possèdent une vessie natatoire qui s’ouvre dans la bouche et qui peut être simple ou double, composée de nombreux espaces cellulaires. Cette vessie natatoire est une excroissance de la région antérieure du tube digestif et sert de poumon, complétant, voire remplaçant en période de sécheresse, les fonctions des branchies comme organes respiratoires. D’un point de vue évolutif, les dipneustes sont très importants car leur vessie natatoire est comparable [ p. 299 ] au poumon des vertébrés supérieurs : elle ramène le sang oxygéné directement au cœur, alors que chez la plupart des autres poissons, le sang transite par la circulation générale depuis la vessie natatoire avant d’atteindre le cœur. Les sacs nasaux sont situés à l’extérieur du museau et s’ouvrent dans la bouche, une caractéristique que l’on retrouve également chez les amphibiens et les vertébrés supérieurs, mais très rarement chez les poissons. C’est donc chez les dipneustes que l’on observe les possibilités de développement des vertébrés supérieurs, sans pour autant qu’ils en soient nécessairement les ancêtres. À bien des égards, leur structure est intermédiaire entre celle des poissons inférieurs et celle des salamandres amphibies, mais ils ne sont pas considérés comme des chaînons directs entre les poissons et les amphibiens. Il est probable que les ganoïdes à nageoires frangées (crossoptérygiens) aient été les ancêtres des amphibiens.
L’étude des dipneustes vivants nous apprend que certaines espèces (Lepidosiren) vivent dans les ruisseaux et les marais boueux, où elles se nourrissent de plantes. D’autres se nourrissent de vers, d’insectes, de coquillages, de crustacés et de grenouilles. Ce sont des animaux lents qui remontent occasionnellement à la surface pour respirer. En période de sécheresse, le Lepidosiren s’enferme dans un cocon de vase et d’argile et respire par une ouverture pratiquée dans la boue. Pendant la saison des pluies, les dipneustes vivants mangent voracement, accumulant d’importantes quantités de graisse entre leurs muscles. Cette graisse leur permet de survivre pendant la saison sèche, parfois jusqu’à huit mois sans nourriture, jusqu’au retour des eaux.
Chez les formes actuelles, on observe deux ou quatre paires de branchies recouvertes d’un opercule osseux mobile. Leur apparence est généralement anguilliforme, les nageoires médianes dorsale et ventrale se prolongeant habituellement par la nageoire caudale. Chez les formes dévoniennes, la queue est généralement de type hétérocerque (Pl., p. 291, Fig. 2), tandis que chez les formes récentes, elle est diphycerque (Pl., p. 295, Fig. 4). Les nageoires paires sont longues, largement lobées, musclées, écailleuses et frangées chez les espèces dévoniennes et chez les Ceratodus actuels, mais chez les autres formes modernes, elles sont très fines, pointues et dépourvues de franges. Le corps est recouvert d’écailles cycloïdes imbriquées.
Les dipneustes sont des poissons cartilagineux présentant une certaine ossification, mais dépourvus de vertèbres à proprement parler. Leur crâne est constitué de cartilage recouvert d’un nombre variable d’os membraneux, plus épais chez les genres du Paléozoïque. Ces os superficiels se développent dans la membrane recouvrant le cartilage et ne se forment pas dans le cartilage lui-même. Leurs dents sont peu nombreuses, généralement disposées en trois paires, dont deux groupes sont de grandes lames broyeuses.
[ p. 300 ]
Les dipneustes sont étroitement apparentés aux ganoïdes à nageoires frangées, ces deux types de poissons étant les lignées les plus importantes des requins primitifs et ayant évolué vers les vertébrés supérieurs. Les dipneustes les plus anciens, une quinzaine d’espèces environ, datent du Dévonien d’Europe et d’Amérique.
¶ Sous-classe Arthrodira ou Poissons cuirassés
Les poissons les plus remarquables des mers du Dévonien étaient les Arthrodères, des poissons fortement cuirassés qui représentaient près de 40 % de tous les poissons du Dévonien. Le terme Arthrodère signifie « cou articulé », car la cuirasse de la tête glisse légèrement sur celle du corps, permettant ainsi à la tête de se mouvoir de haut en bas ; une caractéristique rare chez les poissons et particulièrement développée chez cette sous-classe. Les Arthrodères sont plus ou moins lourdement cuirassés et, à cet égard, rappellent les dipneustes et les premiers amphibiens. Leur relation systématique demeure inconnue, mais il semble qu’ils soient plus proches des dipneustes et des ganoïdes à nageoires frangées. C’est en raison de cette parenté supposée qu’A. Smith Woodward les appelle « dipneustes à grosse tête ». Bien entendu, on ne sait rien des poumons ni des habitudes respiratoires des Arthrodères.
Les plus anciens arthrodires ont été découverts dans les dépôts d’eau douce du Dévonien inférieur d’Allemagne, tandis que ceux du Dévonien moyen et supérieur d’Amérique du Nord étaient marins. Ils étaient les poissons les plus grands et les plus féroces de cette période, mais ont disparu au début du Mississippien, époque où les requins conchylivores ont pris le dessus sur les mers.
Les arthrodires ne possédaient pas de véritables dents ; les structures remplissant cette fonction étaient de grandes lames tranchantes ou des plaques broyeuses. La queue [ p. 301 ] était hétérocerque et il semble qu’ils aient eu une paire de doigts pelviens. Le squelette interne était rudimentaire, car l’axe vertébral n’était pas osseux, bien que les arcs neural et ventral fussent quelque peu ossifiés.
Les Arthrodira dominaient aisément la vie aquatique des mers du Dévonien. Une espèce américaine, Dinickthys, le « poisson terrible », atteignait une longueur de plus de 6 mètres (Fig., p. 300). On connaît plus de quarante espèces en Amérique du Nord, et un seul site en Allemagne (Wildungen) en a recensé plus de cinquante.
¶ Sous-classe Téléostéens ou Poissons osseux
Les téléostéens (signifiant « vrais os ») sont les poissons modernes et présentent l’organisation la plus poussée parmi ces animaux, leur évolution les ayant conduits à une adaptation complète au milieu aquatique (voir Fig., p. 289). Ils constituent environ 99 % de tous les poissons actuels. La plupart des poissons d’intérêt commercial et sportif appartiennent à cette sous-classe et sont abondants en eaux douces, en mers et en océans. Originaires des Ganoidei, leur origine remonte vraisemblablement à la fin du Paléozoïque, mais ils ne se sont répandus qu’au Crétacé inférieur, leur développement maximal ayant eu lieu au Cénozoïque et se poursuivant jusqu’à nos jours.
Les poissons osseux sont presque toujours recouverts d’écailles cycloïdes fines et élastiques, jamais d’écailles placoïdes, et très rarement d’écailles ganoïdes. Chez certains, la peau est nue et visqueuse. La nageoire caudale est généralement homocerque et les membres pairs ne sont jamais lobés. Les branchies sont toujours recouvertes d’un opercule mobile. La vessie natatoire, allongée, lorsqu’elle est présente, est remplie de gaz et se situe dorsalement à la cavité corporelle ; il ne s’agit pas d’un poumon mais d’un organe hydrostatique. Le squelette interne est entièrement ossifié, les vertèbres sont creuses à leurs deux extrémités (amphicoeles), un arc neural osseux entoure la moelle épinière, et chaque vertèbre possède également une paire de côtes renfermant les viscères. Nombre de ces caractères se retrouvent aussi, mais de façon moins marquée, chez les Ganoidei, et c’est pourquoi certains naturalistes considèrent les deux sous-classes comme ne représentant qu’un seul et même groupe.
¶ Origine des nageoires et des membres
Développement des nageoires. — Chez les animaux marins supérieurs, la locomotion volontaire débute par l’immobilité latérale d’un corps flexible, auquel se sont ajoutées les nageoires pour faciliter la progression dans l’eau. Il existe deux types de nageoires : les nageoires impaires et les nageoires paires. Les nageoires impaires, initialement flexibles, se sont développées à partir de la peau et se sont rigidifiées par la suite, [ p. 302 ] offrant au poisson une surface plus résistante. Elles peuvent être plus ou moins continues, partant de la tête, longeant la ligne médiane du dos (nageoire dorsale), contournant la queue (nageoire caudale) et s’étendant vers l’avant le long de la face ventrale jusqu’au cloaque (nageoire anale). Chez la plupart des poissons, ces nageoires impaires sont généralement discontinues, c’est-à-dire localisées.
Les nageoires paires, comme les impaires, proviennent très probablement, selon Lull, de replis cutanés se développant de part et d’autre du corps. Elles servaient initialement de carènes, assurant un meilleur équilibre au corps étroit du poisson ou freinant sa propulsion. On les observe clairement chez les requins acanthodiens du Silurien et du Dévonien (Pl., p. 295, Fig. 1), où elles étaient maintenues par de robustes épines osseuses, à l’instar des nageoires dorsales impaires, verticales, chez les requins modernes. Chez d’autres espèces, ces épines osseuses latérales ont été remplacées par des tiges de rigidification internes. Osborn explique qu’au cours de l’évolution, ces tiges se concentrent pour former l’axe central d’une nageoire entièrement articulée, puis se transforment, avec la suite du développement, en cartilages et en os des ceintures scapulaire et pelvienne (voir Fig. ci-dessus).
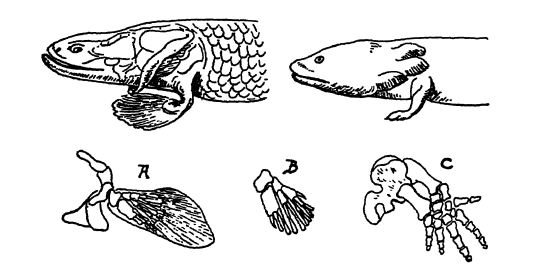
Chez les poissons, les nageoires paires et leurs ceintures constituent les rudiments à partir desquels les pattes et les pieds se sont développés grâce aux déplacements forcés des ganoïdes à nageoires frangées à la recherche de points d’eau dans les régions désertiques. On pense qu’une structure précurseure des pattes et des pieds s’est développée chez eux, bien que le membre transitionnel actxial, que ce soit sous forme fossile ou dans les embryons de vertébrés à membres actuels, soit totalement inconnu.
La plus ancienne empreinte connue, celle de Thinopus (Fig., p. 331), datant du Dévonien supérieur, pourrait nous éclairer sur cette évolution. On n’y distingue que deux doigts complètement formés, probablement le premier et le deuxième, séparés [ p. 303 ] par une articulation qui s’enfonçait profondément dans la plante du pied ; le fossile indique par ailleurs la présence d’un troisième orteil nidifuge, voire d’un quatrième. Si l’on se fie à la structure de cette empreinte, alors le pied terrestre a débuté avec deux orteils au lieu de cinq, comme on le pensait généralement.
¶ Origine de la double respiration chez les poissons
Organes respiratoires des animaux marins. — L’étude des animaux marins les plus communs révèle de nombreuses et diverses méthodes d’extraction de l’oxygène libre de l’eau. Cependant, quel que soit le type d’organe respiratoire, il repose toujours sur la même structure fondamentale : un ou plusieurs organes localisés, dotés de canaux délicats aux membranes très fines où circule le sang, mettant ainsi les globules rouges en contact quasi direct avec l’eau environnante. L’hémoglobine de ces globules rouges a une affinité pour l’oxygène, qu’elle extrait de l’eau et que les cellules transportent par la circulation sanguine vers toutes les parties du corps de l’animal. Aucun animal marin ne possède d’organe supplémentaire permettant l’extraction directe de l’oxygène de l’air, à l’exception des espèces connues pour avoir des ancêtres terrestres.
Évolution des poumons. — Chez la plupart des poissons, à l’exception des requins, se trouve un organe appelé vessie natatoire, qui est une excroissance en forme de sac, unique ou paire, du tube digestif. La fonction principale de cette structure dégénérée et secondairement modifiée chez la plupart des poissons actuels est celle d’un organe hydrostatique, assurant la flottabilité. Elle est contrôlée par des muscles permettant à son possesseur de se maintenir à la profondeur souhaitée dans l’eau. La vessie natatoire est l’homologue des poumons des vertébrés terrestres et est utilisée par les dipneustes, et dans une moindre mesure par d’autres poissons reliques (Amia, Lepidostevs, Pdypterus), comme organe respiratoire complémentaire aux branchies.
La grande variété de modifications de cette structure témoigne de son origine extrêmement ancienne, et son état initial est peut-être illustré par la présence, chez les requins, de deux excroissances en forme de poches au niveau du pharynx, ou cavité gulaire. La stagnation de l’eau et la raréfaction de l’oxygène libre contraindraient les poissons à remonter à la surface pour respirer, et ces poches, si elles étaient vascularisées, contribueraient de façon rudimentaire à l’aération du sang. On pense que le développement de ces structures aurait favorisé leur évolution jusqu’à l’état observé chez les dipneustes modernes.
Origine des poumons chez les animaux vivant dans des étendues d’eau temporaires. — Les animaux originaires et vivant [ p. 304 ] dans des étendues d’eau permanentes, et notamment dans l’océan, n’ont pas besoin de respirer l’air. On suppose donc que le stimulus à l’origine d’une telle modification pourrait être apparu là où l’eau venait périodiquement à manquer à ces animaux. Aucun poisson respirant l’air en permanence n’est connu pour habiter la zone située entre les marées basses et hautes, bien que plusieurs espèces y vivent temporairement à la recherche de nourriture. Par ailleurs, on ne trouve dans aucun des deltas marins actuels les conditions climatiques nécessaires pour contraindre les poissons à se développer en poissons à poumons, ni dans les zones de rivage ou le long des cours d’eau permanents. Il n’existe nulle part dans ces lieux une zone de transition forçant les poissons aquatiques à s’adapter à la terre ferme.
Tout comme nous connaissons aujourd’hui des régions arides, il semble que des climats terrestres similaires aient existé pendant une grande partie des temps géologiques. Sous ces climats, les étendues d’eau fluctuent au gré des saisons, et les animaux ont donc recours à diverses stratégies pour assurer leur survie pendant les périodes de sécheresse. Durant la saison sèche, la lutte pour la survie est intense, non seulement en raison de la promiscuité extrême des individus dans des espaces de plus en plus restreints et de la raréfaction de la nourriture, mais surtout à cause de la salinité et de l’amertume croissantes de l’eau. On pense que, sous l’effet de ces changements, les poissons à respiration branchiale se sont d’abord adaptés à s’enfouir dans le sable. Ainsi protégés dans l’eau et les trous de boue, l’humidité pouvait un temps atteindre leurs branchies, mais dans ces environnements, la vie était très précaire et la plupart des individus périssaient dans la lutte pour la survie. Après d’innombrables échecs dans leurs efforts pour avaler l’air dans le pharynx, efforts qui ont duré pendant de longues périodes géologiques, les ganoïdes et les dipneustes se sont progressivement développés et perfectionnés, leur première apparition remontant au Dévonien le plus ancien.
Mental suprême chez les vertébrés terrestres. — La respiration aérienne, une fois établie chez les poissons, a permis aux vertébrés terrestres d’atteindre une perfection mécanique et mentale suprême, une évolution rendue nécessaire par l’adaptation à une grande variété de conditions environnementales, contrairement à l’uniformité relative du milieu marin. Le mental le plus développé en mer provient souvent des continents, car de nombreuses espèces de vertébrés terrestres se sont adaptées à la mer en raison de la facilité avec laquelle elles peuvent y chasser les êtres moins vigilants et intelligents. Cependant, de ces espèces ne découle pas un mental supérieur. Elles représentent une adaptation dans la mauvaise direction, [ p. 305 ] c’est-à-dire vers une vie plus facile, car le mental le plus sophistiqué ne s’est développé que sur terre, où la lutte pour la survie est la plus intense en raison de la nécessité constante de s’adapter à un environnement changeant. La suprématie organique ne s’atteint que par une vigilance constante.
¶ Lecture collatérale
Joseph Barrell, Influence des climats siluriens-dévoniens sur le Ri«e des vertébrés à respiration aérienne. Bulletin de la Société géologique d’Amérique, Vol. 17, 1916. pp. 387-436.
T. C. Chamberlin, Sur l’habitat des premiers vertébrés, Journal of Gcolo©% Vol. 8, 1900, pp. 40(M12.
Bashford Dean, Poissons, vivants et fossiles. New York et Londres (Macmillan), 1895.
C. R. Eastman, Poissons dévoniens des formations new-yorkaises. Musée de l’État de New York, Mémoire 10, 1907.
R. S. Lull, L’évolution organique. New York (Macmillan), 1917.
H. F. Osborn, L’origine et l’évolution de la vie, New York (Scribner), 1917.