| XXVII. La période pennsylvanienne, l'époque de la plus grande production de charbon. | Page de titre | XXIX. Le charbon et sa présence dans la nature |
[ p. 373 ]
On trouve des plantes de toutes sortes sur tous les terrains et sous tous les climats. Elles ne se limitent pas aux sols humides des vallées, mais poussent aussi sur les plus hautes montagnes et même, plus rarement, dans les déserts les plus arides. Dans les eaux douces peu profondes, on rencontre de nombreuses espèces végétales, et sur les fonds marins, jusqu’à environ 180 mètres de profondeur, poussent des algues fixées. Les lichens bruns et gris, une fine pellicule de matière végétale, s’accrochent aux rochers, aux arbres et à d’autres objets. Dans les forêts humides et les prairies, on trouve des fimgi, notamment les champignons, des plantes dépourvues de chlorophylle qui se nourrissent de la matière humique (moisissure végétale) d’autres végétaux morts ou mourants. Une grande variété de plantes microscopiques, les bactéries, vivent partout : dans le sol, dans presque toutes les eaux, dans les océans, sur et dans d’autres plantes et animaux. Elles et les champignons sont les principaux agents naturels de la décomposition des organismes morts, consommant de l’oxygène et rejetant du dioxyde de carbone. Les levures transforment le sucre en alcool par fermentation et sont utilisées dans la fabrication des boissons alcoolisées ; d’autres bactéries sont utiles dans la fabrication du fromage ; d’autres encore sont pathogènes et se retrouvent même chez tous les organismes sains.
Flores du Paléozoïque. — Les archives géologiques nous apprennent que la première flore connue est apparue dans la seconde moitié du Dévonien, bien que quelques plantes terrestres soient attestées dès le Silurien. Il est probable, cependant, que les basses terres étaient couvertes d’une végétation luxuriante au moins depuis le début du Champlainien, mais que les premières plantes n’avaient pas encore acquis suffisamment de tissu ligneux pour se conserver dans les sédiments et, de ce fait, n’ont aucune importance en géologie historique. Depuis le Dévonien moyen, les plantes terrestres à tissu ligneux sont devenues de plus en plus prolifiques et ont atteint des dimensions et une stature plus importantes, ce qui signifie qu’elles ont également progressivement acquis une plus grande taille.
La vie des marais du Pennsylvanien. — La flore des charbons de la fin du Paléozoïque est aujourd’hui bien connue, et on estime que plus de trois mille espèces ont été décrites (voir fig., p. 360 et 374). Elle se distingue par sa répartition [ p. 375 ] [ p. 374 ] quasi mondiale et sa luxuriance, ainsi que par un sous-bois dense et varié. Ses représentants les plus remarquables, tant par leur nombre que par leur taille, sont les arbres à écailles, une sorte de conifères aux feuilles relativement petites et aciculaires ; certains de ces arbres atteignaient plus de 30 mètres de hauteur et un diamètre allant jusqu’à 1,8 mètre (voir pl., p. 377). On observe chez eux une absence de cernes de croissance, indiquant également une absence de saisons jusqu’à la fin du Pennsylvanien. Un autre groupe remarquable, les calamites ou joncs géants (pl., p. 381), atteignaient au moins 18 mètres de long et 38 centimètres d’épaisseur. Elles ressemblaient aux fourrés de cannes et aux bambouseraies d’Uving. Cette flore comprenait également de nombreuses formes ressemblant à des fougères, à la fois délicates et robustes, dont certaines étaient grimpantes, tandis que d’autres se développaient en arbres majestueux (Fig., p. 374) ; la plupart portaient des graines, mais certaines étaient sporifères et donc de véritables fougères.

En général, ces forêts devaient atteindre une hauteur de plus de 12 mètres. Elles étaient caractérisées par une croissance rapide et un bois tendre, voire spongieux, comme en témoignent l’écorce lisse, dure et persistante, les cellules larges et à parois fines du bois, la quantité relativement importante de tissu médullaire et cortical et les grands espaces intercellulaires, ainsi que les nombreux pores aquifères des feuilles.
Les nuances de vert étaient dominantes, et la monotonie de la verdure n’était nulle part égayée par des fleurs éclatantes. Des fleurs étaient toutefois présentes, mais de faible qualité, insignifiantes par leur taille et sans doute peu attrayantes. Probablement plus de la moitié de la flore était hétérosporée, et l’on peut considérer sans risque que la plupart des plantes les plus communes des formations houillères étaient dépourvues de graines. La fécondation ne s’effectuait pas encore par l’intermédiaire d’insectes mellifères et pollinisateurs, comme c’est le cas si souvent chez les plantes à fleurs actuelles, mais était assurée, dans la plupart des formations houillères, par les pluies et les vents. Au moment où les arbres et les fougères sporifères libéraient leurs spores, toute la forêt était recouverte d’une poussière jaune verdâtre ou brune, et certains charbons sont en grande partie constitués de ces spores (White et Jeffrey).
Les spores (Pl., p. 377, Fig. 4-6) diffèrent des graines en ce que ces dernières donnent directement naissance à des plantes sexuées, tandis que les premières se développent également en plantes sexuées, mais celles-ci ont une durée de vie courte et donnent à leur tour naissance aux plantes asexuées et vivaces qui portent les spores. Autrement dit, les plantes à spores présentent une « alternance de générations », tandis que les plantes à graines ont un développement direct. Ce point sera abordé plus en détail ultérieurement dans ce chapitre.
L’air n’était pas parfumé de douces odeurs, car il n’y avait pas de miel, mais il est probable que des odeurs résineuses, telles qu’on les trouve dans les forêts de conifères vivantes, étaient présentes.

Fig. 1, Alethopteris grandifolia ; 2, Neuropteris fasciculata ; 3, N. capitata ; 4, Pseudopecopteris mazonana, avec ses capsules à spores ; 5, probablement un arbre à graines ressemblant à une fougère, reconstitué par Kidston ; 6, Sphenopteris mixta ; 7, Archaeopteris stricta ; 8, Aneimites (x2) ; 9, même espèce avec une jeune graine (x4) ; 10, même espèce, une des graines (x3). Fig. 1 à 4, d'après les inventaires des États de l'Illinois et de l'Ohio ; Fig. 8 à 10 d'après David mite.
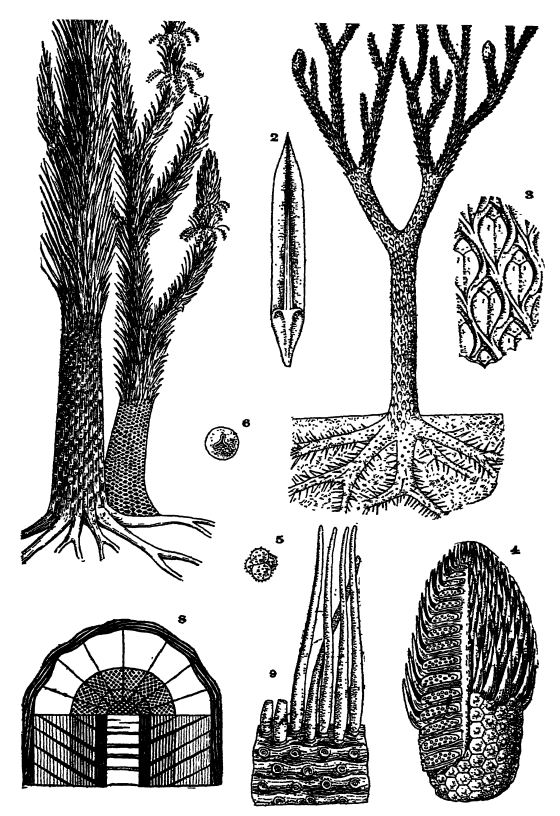
Fig. 1, restauration de l'arbre Lepidodendron, environ x 1/100 ; 2, feuille du cône sporifère, environ x ⅓, celles des branches étant très similaires mais plus longues ; 3, surface externe de l'arbre, montrant la nature des bases des feuilles, environ x ⅂ (L. cbovatum) ; 4, cône sporifère (Lepidostrobus), avec et sans feuilles, sectionné pour montrer les spores et les sporanges ; 5, groupe de quatre microspores ; 6, macrospore ; 7, deux espèces de l'arbre Sigtllaria, restaurées, environ x A ; 8, section du tronc, montrant les épaisseurs relatives de la couche externe foliaire, deux zones d'écorce liégeuse, un cylindre interne de bois relativement mince et la moelle médiane ; 9, racines de ces arbres (Stigmaria ficoides), avec radicelles et cicatrices de leur insertion, fig. 1-6 d'après Steinmann ; Fig. 7 et S d'après Zittel. (677)
[ p. 378 ]
Dans ces forêts du Pennsylvanien, on aurait pu observer les plus grands insectes ayant jamais existé, de gigantesques libellules atteignant une envergure de 74 cm. D’énormes blattes pullulaient partout, et il n’était pas rare d’en trouver des géantes de 10 cm de long (voir Pl., p. 363, Fig. 4). Ces insectes étaient généralement carnivores et ne transportaient pas le pollen d’une fleur à l’autre, évitant ainsi la pollinisation croisée si fréquente chez les insectes actuels. Les plus petits insectes étaient la proie des scorpions et des araignées, ces dernières ne tissant pas de toiles mais vivant au sol ou dans des troncs pourris, en compagnie de nombreux myriapodes. À notre connaissance, aucun insecte de cette époque n’émettait de chants ni d’autres sons, et le bruissement du vent dans les arbres n’était interrompu que par le coassement des amphibiens dans les marais. Les amphibiens étaient communs dans les marais, et il est probable que de nombreux petits reptiles couraient sur le sol et dans les arbres. On n’y voyait aucun animal terrestre de grande taille, tel que nous le connaissons, ni aucun oiseau.
Classification. — Pour nos besoins, les plantes sont plus commodément divisées en (1) celles qui n’ont pas de fleurs mais se reproduisent par spores et passent par une alternance de générations (Cryptogames), comme expliqué ci-dessus ; et (2) celles qui ont des fleurs et se reproduisent directement à partir de graines (Phanérogames).
Dans ce chapitre, nous allons étudier les premières plantes terrestres, et comme celles-ci ne peuvent être conservées sous forme de fossiles que si elles possèdent une matière fibreuse ou ligneuse, le paléontologue ne s’intéresse pas directement aux Cryptogames inférieurs.
Toutes les plantes supérieures aux mousses, du point de vue de leur organisation, se caractérisent par la présence de racines et, dans leurs tissus mous, de faisceaux vasculaires. Ces faisceaux sont composés de fibres creuses, résistantes et entrelacées, dont la fonction principale est de servir de canaux conducteurs (d’où le terme « vasculaire », dérivé de « vascidum », petit vaisseau) par lesquels transitent les différentes substances nutritives : l’eau et les sels minéraux montent, tandis que les sucres et les protéines descendent. Secondairement, ils constituent le squelette de soutien de la plante. Toute la matière ligneuse est composée de ces fibres, et c’est cette substance résistante qui permet aux feuilles et autres parties des plantes d’imprimer sur les sédiments la trace de leur existence passée.
La classification complémentaire peut être exprimée comme suit :
Cryptogames inférieures, plantes à spores
- Thallophytes, bactéries, champignons, la plupart des plantes marines (algues)
- Bryophytes, mousses
Cryptogames supérieurs, également porteurs de spores
- Ptéridophytes, fougères (Pl., p. 376, Fig. 1-5)
- Arthrophytes, joncs vivants et calamites paléozoïques (Pl., p. 381, Fig. 1, 2)
- Lépidophytes, arbres à écailles ou lycopodes (Pl., p 377)
[ p. 379 ]
Phanérogames, plantes à graines
- Gymnospenns, plantes à fleurs discrètes et imparfaites
- Ptéridospermophytes (ou Ptéridospermes, plantes ressemblant à des fougères (Pl., p. 376, Fig. 6-10)
- Cycadophytes, cycadées (Fig., pp. 27 et 3S6)
- Conifères
- Cordaites, arbres anciens à feuilles persistantes (Pl., p. 381, Fig. 3-6)
- Conifères, arbres à feuilles persistantes modernes
- Ginkgos, arbres aux quarante écus (Pl., p. 381, Fig. 7)
- Les angiospermes, les vraies plantes à fleurs
¶ Plantes sans fleurs à spores
¶ Fougères ou ptéridophytes
Presque tout le monde sait à quoi ressemble une fougère vivante, mais lorsqu’on applique ces connaissances aux plantes du Paléozoïque d’apparence similaire, même les botanistes les plus éminents ne peuvent, à l’heure actuelle, être certains, dans bien des cas, que la forme étudiée soit une véritable fougère. Au siècle dernier, on croyait encore que la moitié des plantes du Carbonifère étaient de véritables fougères et que ces magnifiques plantes dominaient la flore de cette époque. Cependant, grâce à l’accumulation des connaissances botaniques, et plus particulièrement à la découverte de spécimens portant les organes reproducteurs, on sait aujourd’hui que nombre de ces fougères dites paléozoïques portaient des graines et étaient donc d’une organisation plus complexe ; elles doivent être classées à part des véritables fougères. Il s’agissait des fougères à graines, à bien des égards une forme de transition entre les fougères à spores et les plantes à graines (voir Pl., p. 376, Fig. 6-10). Il est probable que plus de la moitié des plantes du Paléozoïque autrefois considérées comme des fougères (environ un millier d’espèces) se révéleront finalement être des plantes à graines.
Fougères vivantes. — On compte aujourd’hui plus de six mille espèces de fougères différentes, allant de formes minuscules et délicates aux magnifiques fougères arborescentes des régions tropicales, qui peuvent atteindre 15 mètres de hauteur. La plus grande diversité se rencontre sous les tropiques, mais les fougères herbacées sont présentes sous tous les climats, même au Groenland, pays glacé. En règle générale, elles préfèrent les régions boisées et humides. La fougère est une plante asexuée. Ses spores se développent généralement sur la face inférieure des feuilles, dans des capsules à spores, toujours regroupées en plusieurs amas. Chaque capsule contient une multitude de spores microscopiques identiques, souvent produites en très grand nombre (voir Pl., p. 376, Fig. 4).
Quelques-unes de ces spores peuvent germer dans le sol, donnant naissance à une plante sexuée, le prothalle, généralement une pousse primitive verte en forme de cœur, rampante et mesurant [ p. 380 ] souvent moins de 1,25 cm de long. Sur sa face inférieure se trouvent des organes mâles développés qui produisent des cellules mâles ou du pollen, et des organes femelles qui, une fois pollinisés, transforment certaines cellules en plantes asexuées. On observe donc ici une alternance de générations : des plantes asexuées donnent naissance à des plantes sexuées, qui à leur tour produisent les fougères asexuées.
Chez les petites fougères herbacées, une tige courte et robuste, recouverte d’un sous-bois dense, donne naissance à de nombreux pétioles. Chacun d’eux se développe enroulé en spirale et le reste pendant un an, voire plus. Par la suite, les pétioles se déroulent et croissent rapidement pour former des feuilles pleinement développées, généralement appelées frondes. Celles-ci peuvent atteindre une grande longueur, comme chez les fougères arborescentes, et sont nommées frondes car elles sont composées de nombreuses folioles.
Chez les fougères arborescentes, il n’y a qu’une seule tige non ramifiée, semblable à un tronc de palmier, et les frondes, composées d’une ou plusieurs feuilles à nombreuses folioles, se développent au sommet en une couronne, chaque fronde se déroulant à mesure qu’elle atteint sa pleine croissance. Le tronc porte les cicatrices des frondes tombées et, sous cette couche externe, se trouve souvent une zone épaisse, l’écorce, qui entoure le cylindre ligneux central, plus ou moins grand (Pl., p. 376, Fig. 5).
Fougères fossiles. — Les fougères ancestrales (Marattiales, ou fougères issues d’une seule cellule) étaient rares au Dévonien et ne sont pas fréquentes à l’état fossile avant le Pennsylvanien, où de nombreuses espèces herbacées de petite taille sont associées aux fougères arborescentes (voir Pl., p. 376, Fig. 1, 4). Ces dernières deviennent visibles pour la première fois au Pennsylvanien et sont abondantes dans la seconde moitié de cette période et au début du Permien. La tige pouvait atteindre 15 mètres de hauteur et, chez certaines formes, les frondes se développaient en deux ou quatre colonnes parallèles, mais le plus souvent, elles étaient disposées en spirale autour du tronc (voir Pl., p. 376, Fig. 5).
¶ Arthrophytes
Parmi les plantes vivantes, on trouve un petit groupe de formes très répandues, communément appelées joncs (semblables à Pl., p. 381, Fig. 1, 2). En règle générale, elles sont petites, mesurant moins de 45 cm de haut, et apparaissent dans les lieux humides en début d’année. En Amérique centrale et du Sud, ainsi qu’à Cuba, une forme géante se rencontre en bosquets, atteignant 12 m de hauteur, mais avec une tige ne dépassant pas 2,5 cm de diamètre. Cette forme, ainsi que toutes les autres formes vivantes, environ vingt-cinq, appartiennent au genre Equisetum (voir aussi Fig., p. 468).
Ce genre vivant de joncs possède dans le sol un rhizome horizontal asexué, vivace, rampant et ramifié, qui donne naissance, [ p. 382 ] à intervalles réguliers, à des tiges aériennes dressées et à des racines descendantes. Les tiges sont constituées d’une fine zone ligneuse, striée extérieurement, avec une moelle centrale importante. À intervalles réguliers, la tige est divisée en entre-nœuds par des cloisons transversales appelées nœuds, d’où émergent les verticilles de petites feuilles ou de feuilles réduites, munies d’une ou plusieurs nervures longitudinales, ainsi que les branches lorsqu’elles sont présentes. Les branches reprennent, à plus petite échelle, les caractéristiques générales des tiges. Les cônes sporifères se trouvent au sommet de la tige ou sur les branches.

Fig. 1, calaminés reconstitués, celui de gauche portant une capsule à spores, environ x 1/100 ; 2, extrémité basale d’un calaminé ; 3, cordaïte reconstitué (Dorvcordaita), environ x 1/100, montrant également la nature des fleurs en forme de chaton ; 4, différents types de cordaïtes environ x 1/40 ; 5, Trigonocarpon ornatum, un fruit ou une graine de cordaïte ; 6. Rhabdocarpus apiculatus, graine de cordaïte ; 7, rameau de ginkgo biloba vivant, fleurs mâles, et en haut la graine mûre.
Calamites fossiles. — À l’ère paléozoïque, du Dévonien au Permien, et plus particulièrement au Pennsylvanien, vivait une grande variété de joncs anciens, dont les plus grands sont connus sous le nom de calamites (Pl., p. 381, Fig. 1, 2). Ces plantes étaient prolifiques et, selon White, elles atteignaient en Amérique un diamètre de 30 cm et une longueur de plus de 9 m. Leur tronc était beaucoup plus épais que celui des joncs actuels, et certains spécimens présentent une écorce de 5 cm d’épaisseur, en plus d’un bois de même épaisseur. Des couches de bois secondaire se formaient sous l’écorce, un type de croissance que l’on retrouve chez les arbres modernes. Les nœuds n’étaient pas équidistants comme chez les formes actuelles, mais s’écartaient progressivement au cours de la croissance initiale, et les tiges s’épaississaient rapidement. Les calamites étaient également beaucoup plus ramifiées que les joncs actuels, et leurs feuilles étaient proportionnellement plus grandes. Le sommet du tronc était constitué d’une touffe dense de cercles de feuilles étroitement imbriqués, et les cônes contenant les spores reproductrices étaient plus ou moins nombreux sur les rameaux feuillus ou au sommet de la tige principale. En règle générale, les spores étaient toutes identiques chez les arthrophytes du Paléozoïque, mais dans certains cas, elles se différenciaient en petites et grandes spores (hétérospores, voir Pl., p. 377, Fig. 5, 6).
¶ Arbres à écailles ou lépidophytes
Lycopodes vivants. — Les lycopodes désignent un groupe de plantes herbacées primitives, persistantes et très répandues, connues sous le nom de lycopodes rampants, ces derniers étant fréquemment utilisés en art floral. Le genre Lycopodium compte une centaine d’espèces vivantes. Son nom, dérivé du grec « pied de loup », fait référence à l’aspect de ses racines. Dans les flores actuelles, les lycopodes ne forment jamais de tapis végétaux denses et sont donc peu visibles. Leur aspect général évoque les grandes mousses ou les plus petites branches de pins, car leurs feuilles sont toujours remarquablement petites.
Lépidophytes fossiles. — Au Paléozoïque, du Dévonien au Permien moyen, et plus particulièrement au Pennsylvanien, les lépidophytes [ p. 383 ] étaient les plantes dominantes et atteignaient des proportions gigantesques comparées aux lycopodes vivants, considérablement réduits, car les premières étaient alors à l’apogée de leur évolution (Fig., p. 360 et 374). Ces arbres, au Pennsylvanien, libéraient leurs germes reproducteurs hétérosporés en si grande quantité qu’ils constituaient un facteur important dans la formation du charbon (voir Pl., p. 377, Fig. 4-6).
À la fin du Paléozoïque, on distinguait deux principaux types de lépidophytes : les Lepidodendron (ou arbres à écailles) et les Sigillaria (ou arbres à sceaux), termes qui font référence à l’aspect écailleux de la base des feuilles sur les troncs et les branches de ces arbres. Plus d’une centaine d’espèces de Lepidodendron ont été décrites, datant du Dévonien au Permien (Pl., p. 377, fig. 1-3). Le tronc s’effilait progressivement et, chez certaines espèces, atteignait plus de 30 mètres de hauteur et 90 cm de diamètre. Ces arbres se distinguaient de tous les arbres modernes par le fait que leur tronc et nombre de leurs branches se divisaient régulièrement en deux branches. Les branches les plus fines se terminaient généralement par des cônes sporifères linéaires ou ovales de 2,5 à 30 cm de long (Pl., p. 377, fig. 4).
Les feuilles de Lepidodendron étaient aciculaires, toujours relativement petites, mais mesurant 1,25 cm de large à la base et de 15 à 18 cm de long (Pl., p. 377, Fig. 1, 2). Sur leur face supérieure se trouvaient deux sillons latéraux, dans lesquels étaient situés les minuscules mais nombreux organes respiratoires permettant l’extraction du dioxyde de carbone de l’atmosphère. Les feuilles tombaient rapidement et ne se trouvaient généralement attachées qu’aux jeunes rameaux en croissance. La base des feuilles était en forme de losange et disposée en spirale sur les branches et le tronc, formant ainsi les marques caractéristiques de l’arbre. La feuille était insérée dans la partie supérieure du losange (Pl., p. 377, Fig. 3).
Environ une centaine d’espèces de Sigillaria (Pl., p. 377, Fig. 7) ont été décrites. Elles se distinguent des Lepidodendron par leur faible ramification. On a trouvé des troncs d’un diamètre de 1,80 mètre au-dessus des racines, et l’un d’eux atteignait près de 30 mètres de hauteur sans être ramifié. À leur extrémité, sur environ 3 mètres, ces arbres étaient couverts de feuilles dressées, rigides, semblables à des graminées, généralement semblables à celles des Lepidodendron, mais parfois beaucoup plus grandes et plus larges.
Chez Sigillaria, les cicatrices foliaires étaient disposées en rangées verticales, les cicatrices de séries adjacentes alternant entre elles. Chez de nombreuses formes, la surface du tronc était nervurée longitudinalement, chaque nervure portant une seule rangée de cicatrices foliaires (voir Pl., p. 377, Fig. 7).
[ p. 384 ]
Les Sigillaria sont inconnues au Dévonien et ont disparu avant la fin du Pennsylvanien.
La structure du tronc et des branches des Lepidophyies fossiles était particulière en ce que la plus grande épaisseur sous la couche externe avec les cicatrices foliaires était composée de deux types d’écorce liégeuse entourant un cylindre ligneux interne relativement mince qui pouvait être entièrement rempli de bois à croissance interne — le type de croissance endogène — ou avec plus ou moins de moelle (Pl., p. 377, Fig. 8). Du bois secondaire jusqu’à 2 pouces d’épaisseur était ajouté au liège externe sous la couche foliaire, et comme ce liège était également dur, les arbres avaient une force de résistance considérable contre les vents et les tempêtes de cette époque, mais étaient moins rigides que les arbres forestiers modernes.
À leur base, les genres Lepidodendron et Sigillaria se terminaient par quatre à sept racines principales qui s’étendaient probablement de manière quasi horizontale dans le sol. Contrairement aux arbres modernes, il n’y avait pas de racine pivotante ou verticale prolongeant le tronc. Ces racines principales se divisaient une ou deux fois et s’effilaient en pointe, s’enfonçant dans toutes les directions à travers la masse de végétation en décomposition du marais. Leur surface présentait des cicatrices circulaires disposées par groupes de cinq, auxquelles étaient attachés des appendices rubanés et arrondis rayonnant dans toutes les directions. Ces derniers atteignaient une longueur de 38 cm et servaient à extraire la nourriture et l’eau du sol (Pl., p. 377, Fig. 9). Ces racines sont connues sous le nom de Stigmaria (du latin stigmate, marque) et sont très fréquemment observées dans les couches d’argile sous les gisements de charbon.
Les cônes des lépidophytes, bien que ressemblant beaucoup à ceux des conifères ou des arbres à feuilles persistantes modernes, ne contenaient pas de graines mais étaient remplis de spores (Pl., p. 377, Fig. 1, 4-6). Ces dernières se développaient dans de grandes capsules contenant soit de minuscules spores, les microspores ou organes reproducteurs mâles (Pl., p. 377, Fig. 5), soit de grandes spores, les macrospores femelles (Pl., p. 377, Fig. 6), souvent quarante fois plus grandes que les microspores. Ces deux types de spores se développaient soit dans des cônes différents, soit dans différentes parties d’un même cône, et les minuscules spores étaient beaucoup plus nombreuses que les grandes.
¶ Plantes à fleurs à graines ou phanérogames
Les plantes à graines, extrêmement variées et nombreuses, se divisent en deux grands groupes. Le groupe géologiquement le plus ancien et le plus primitif est celui des gymnospermes (ou plantes à graines nues), chez lesquelles les organes mâles et femelles peuvent se trouver dans des fleurs séparées ou réunis en une seule, mais sont toujours discrets et souvent [ p. 385 ] de couleur terne. Les graines se développent dans les ovaires ouverts, dits nus car le pollen se dépose directement sur les ovules (œufs), tandis que chez les plantes à fleurs supérieures ou véritables (angiospermes), l’ovaire est recouvert d’un stigmate qui reçoit le pollen.
¶ Fougères à graines ou ptéridospermes
Lors de la discussion sur les fougères à la page précédente, il a été mentionné que de nombreuses plantes du Paléozoïque présentaient toutes les caractéristiques des fougères, mais qu’elles produisaient des graines au lieu de spores. Autrement dit, ces plantes ne connaissaient pas d’alternance de générations pour achever leur cycle de développement ; il s’agissait de plantes sexuées, c’est-à-dire qu’elles se développaient directement à partir de la graine, donnant soit des individus mâles produisant du pollen, soit des individus femelles produisant des graines qui fructifiaient et se développaient en un embryon au sein de la plante. Elles avaient l’apparence générale des fougères et leurs graines ressemblaient beaucoup à celles des cycadées. La découverte de ces Ptéridospermophytes, ou Ptéridospermes, est l’une des plus grandes réussites des paléobotanistes du début du XXe siècle. Dans l’apparition des graines, les Ptéridospermes ont franchi une étape importante, et cette caractéristique est devenue dominante dans les flores ultérieures (voir Pl., p. 376, Fig. 6-10).
Les graines et le pollen étaient portés par des plantes à sexes indépendants. Les graines pouvaient être petites et se trouver à l’extrémité des feuilles ou pendre du côté droit, à l’extrémité des feuilles lobées, ou encore, si elles étaient grandes, être fixées à la nervure médiane épaisse de la fronde ; elles n’ont jamais été présentes dans des cônes. Leur taille variait de 6 mm à au moins 5 cm de longueur, et elles étaient souvent allongées à leur extrémité libre, où se situait l’entrée de la chambre pollinique. L’embryon, ou graine, était entouré d’une enveloppe qui, chez certaines espèces, présentait une couche extérieure épaisse et charnue et une couche intérieure dure et pierreuse. Des graines similaires ont également été observées chez les cordaïdes, les ginkgos (Pl., p. 381, fig. 5-7) et les cycadées.
Certaines espèces à graines femelles possédaient de longues tiges fines et grimpaient autour d’autres plantes ; beaucoup étaient herbacées, et d’autres (Psaronius) étaient aussi hautes et robustes que les fougères arborescentes. Elles descendaient de fougères primitives antérieures au Dévonien moyen, et leur développement atteignit son apogée au Pennsylvanien et au début du Permien, mais aucune n’est connue à ce jour au-delà du Paléozoïque.
¶ Cycadées ou Cycadophytes
Dans les flores modernes des régions chaudes, les cycadées actuelles, qui comptent environ 110 espèces, sont les vestiges d’un groupe autrefois plus diversifié, peu commun dans les flores paléozoïques, mais dominant dans celles de la première moitié du Mésozoïque. [ p. 386 ] Un exemple vivant frappant est le sagoutier de Ceylan, source de notre sjigo comestible. L’importance de ces plantes à graines anciennes par rapport aux autres plantes à fleurs nous a été apprise principalement grâce aux travaux de G.R. Wieland ; elles semblent être issues des ptéridospermes, peut-être dès le Dévonien. Les cycadées sont apparues avec le Pennsylvanien moyen, mais ne se sont répandues qu’au Trias.
La caractéristique principale des cycadées est que leurs tiges ligneuses courtes et colonnaires, à moelle très développée, sont enveloppées d’une épaisse carapace de bases foliaires persistantes, avec une masse intermédiaire feutrée. La taille des troncs varie de très petite taille à près de 18 mètres de long chez les cycadées vivantes. Les cycadées ont une croissance lente et n’atteignent leur maturité que très tard ; un tronc de 1,80 mètre peut avoir mille ans. Elles sont parfois remarquablement bien conservées dans les strates du Mésozoïque. Leurs feuilles sont de diverses sortes, mais généralement pennées, et sont très utilisées en ornementation florale (voir fig., p. 27 et suivantes).

¶ Conifères ou coniférophytes
Cordaites, ou conifères à grandes feuilles. — Au Paléozoïque, dès le Dévonien supérieur, on trouve des troncs dont [ p. 387 ] la structure est assez semblable à celle des pins ou conifères modernes. Au Pennsylvanien, des moulages de ces troncs sont fréquemment observés dans les grès, et dans les schistes bitumineux des gisements de charbon, les longues feuilles rubanées de ces arbres abondent ; elles constituent d’ailleurs souvent une bonne partie du charbon. Ces plantes, connues sous le nom de cordaïtes (d’après le paléobotaniste Corda), présentaient une grande variété de formes et étaient les Génotospermes dominantes du Paléozoïque. C’étaient des arbres à feuilles persistantes, à bois tendre, hauts et élancés, atteignant parfois 37 mètres de haut et 90 centimètres de diamètre. Chez ces grands arbres, les deux tiers du tronc étaient dépourvus de branches, le tiers ou le quart supérieur étant constitué d’une cime dense et riche en grandes feuilles simples (voir Pl., p. 381, Fig. 3-6).
Les feuilles des cordaïtes étaient toujours grandes, certaines mesurant jusqu’à 1,80 mètre de long et 15 centimètres de large. Certaines étaient en forme de lanière, avec des extrémités libres pointues ou arrondies ; d’autres ressemblaient à de l’herbe, mesurant 50 centimètres de long et 1,25 centimètre de large. Le tissu foliaire était épais et présentait des nervures parallèles, comme chez le yucca actuel (voir pl., p. 381, fig. 3 et 4).
Bien que le tronc ligneux des cordaïtes ressemble beaucoup à celui des pins modernes, il s’en distingue par la présence systématique d’une moelle centrale pouvant atteindre un diamètre de 12,5 cm. Le bois neuf se formait sous l’écorce épaisse, autrement dit, il s’ajoutait aux couches précédentes par croissance exogène.
Les cordaïdes étaient apparentées aux conifères, mais, comme le montre la description ci-dessus, les deux groupes différaient considérablement par leur composition. Les cordaïdes étaient très ramifiées uniquement à leur sommet, ne portaient pas de cônes à graines mais développaient celles-ci en chatons, leurs feuilles étaient rubanées et non aciculaires, et enfin leur tronc possédait une moelle. Les cordaïdes étaient plus étroitement apparentées aux ginkgos.
Les ginkgos se distinguent des cordaïtes d’emblée par leurs feuilles beaucoup plus petites et lobées, tandis que leurs troncs sont ramifiés sur presque toute leur longueur (voir Pl., p. 381, Fig. 7).
Conifères ou arbres à feuilles persistantes. — Les arbres à feuilles persistantes, ou véritables conifères de la flore actuelle, dont les graines se développent dans des cônes femelles proéminents et le pollen dans des cônes mâles, ne sont pas connus pour être d’une origine antérieure au Permien, mais constituent aujourd’hui le groupe de gymnospermes le plus répandu. Cet ordre comprend les araucarias, les séquoias ou « grands arbres » de Californie (des arbres apparentés, mais plus petits, remontent au Permien), les pins, les épicéas, les sapins, les genévriers, les mélèzes, les cyprès et les ifs.
[ p. 388 ]
¶ Angiospermes ou plantes à fleurs véritables
Comme les véritables plantes à fleurs n’apparaissent dans la géologie historique qu’au Crétacé, leur description est reportée au chapitre consacré à cette période.
¶ Lecture collatérale
E. W. Berry, Paléobotanique : un aperçu de l’origine et de l’évolution des flores. Rapport annuel de la Smithsonian Institution pour 1918, 1920, pp. 289-407.
C. J. Chamberlaix, Les Cycadées vivantes. Chicago (University of Chicago Press), 1919.
J. M. Coulter et C. J. Chamberlain, Morphologie des Gjunnospermes. Chicago (University of Chicago Press), 1910.
D. H. Scott, Introduction à la botanique structurale, Partie II, Plantes sans fleurs. Londres (Black), 1904.
A. C. Seward, Liens avec le passé dans le monde végétal. Cambridge (University Press), 1911.
D. White, Flore fossile des couches de charbon inférieures du Missouri. U. S. Geological Survey, Monographie 37, 1899.
D. White, La succession stratigraphique des flores fossiles de la formation de Pottsville dans le bassin houiller anthracite du sud de la Pennsylvanie. U. S. Geological Survey, vingtième rapport annuel, partie II, 1900, pp. 749-918.
G. R. Wieland, Araucariales, Cycadales et Cordaitales. Encyclopédie Americana, 1918, pages 135-138, 351-360, 683-686.
| XXVII. La période pennsylvanienne, l'époque de la plus grande production de charbon. | Page de titre | XXIX. Le charbon et sa présence dans la nature |