| II. Les organismes, leur composition, leur structure et leur classification | Page de titre | IV. L'évolution, le changement constant des êtres vivants |
[ p. 22 ]
Les plantes et les animaux vivants constituent le monde organique actuel, tandis que les fossiles sont les restes d’organismes ayant vécu dans le passé géologique. Les strates fossilifères sont les cimetières d’un passé révolu, des civilisations disparues qui relient le passé au présent. « La poussière que nous foulons était jadis vivante » (Byron). Les géologues de l’État de New York, lorsqu’ils commencèrent leurs travaux en 1838, trouvèrent des fossiles « dans les murets de pierre et les fondations des fermes ; ils jonchaient le long des cours d’eau et sur les rives des Finger Lakes ; et ils affleuraient des rochers au bord des falaises. Ils étaient si omniprésents que les Indiens Seneca utilisaient les coquillages fossilisés en forme de coupe pour fabriquer des pipes, assemblaient les articulations des tiges de crinoïdes en colliers et enterraient des coquilles de brachiopodes avec des haches et des pointes de lance dans les tombes de leurs guerriers » (Clarke).
On peut trouver des fossiles dans n’importe quelle roche sédimentaire des plaines côtières ou des terres profondes, voire même dans les plus hautes chaînes de montagnes. Ils renseignent non seulement sur les espèces animales qui y vivaient, mais aussi sur la nature de leur environnement. Par exemple, les restes d’un animal marin, comme une huître, découverts naturellement enfouis dans des strates géologiques n’importe où sur le territoire actuel, indiquent que la mer existait à l’endroit où se trouve ce vestige à l’époque où vivait cet organisme.
Conceptions antiques sur les fossiles. — La découverte de fossiles ressemblant en tous points à des coquilles d’animaux marins loin à l’intérieur des terres a longtemps intrigué l’humanité. Certains Grecs des IIIe et IVe siècles avant J.-C. les expliquaient comme des tentatives infructueuses de création par une force plastique inhérente à la terre. Cette interprétation poétique a refait surface à maintes reprises au cours des siècles suivants, et la controverse sur les fossiles s’est intensifiée après le XVe siècle. Ces découvertes étaient alors considérées par beaucoup comme de simples concrétions minérales, qualifiées d’« Imvs naiurm », des anomalies de la nature, tandis que d’autres affirmaient que ces pierres « figurées » ou « formées » étaient le fruit du travail du sol sous [ p. 23 ] l’influence des astres. D’autres encore soutenaient, à juste titre, que les fossiles ne pouvaient provenir que de plantes et d’animaux ayant vécu autrefois, mais attribuaient leur présence dans les roches au Déluge. Ils croyaient qu’au plus 6 000 ans s’étaient écoulés depuis la Création et que le Déluge avait été universel, détruisant toute vie et dispersant ses vestiges sur toute la surface terrestre. « Ils n’avaient aucune idée de l’impossibilité physique d’accumuler toutes les formations fossilifères de la croûte terrestre en l’espace de cent cinquante jours, période durant laquelle «les eaux recouvrirent la terre et toutes les hautes collines sous le ciel furent submergées» », comme l’a dit Sir Archibald Geikie. C’est ainsi qu’est née la « théorie du Déluge », et avec elle une vive controverse qui ne prit fin qu’à la fin du XIXe siècle.
Dans les pays méditerranéens, cependant, les formations géologiques plus récentes « sous-tendent nombre de plaines et s’élèvent haut sur les flancs des collines. Dans ces dépôts, des coquillages et autres restes de créatures marines ont été conservés en si grand nombre qu’ils ne pouvaient manquer d’attirer l’attention, même aux premiers temps de l’humanité. Puisque ces organismes ressemblent manifestement à ceux qui vivent encore dans la mer voisine, on pouvait aisément en déduire que la mer recouvrait autrefois les étendues de terre où ces vestiges avaient été laissés. Cette conclusion fut tirée par certains des premiers philosophes grecs [notamment Xénophane, 576-480 av. J.-C.], et il ne fait guère de doute qu’elle a conduit à ces conceptions larges des vicissitudes de la Nature, adoptées au cours des siècles suivants par leurs successeurs. » (Geikie.) Ainsi naquit la théorie des « cataclysmes et recréations », qui ne fut abandonnée qu’au siècle dernier au profit de la théorie de l’évolution, laquelle enseigne que la vie, une fois apparue, s’est poursuivie sans interruption jusqu’à nos jours.
Transition du vivant actuel vers les formes fossiles. — La transition aisée entre les mollusques marins (coquillages) actuels et leurs ancêtres géologiques s’illustre parfaitement par les exemples suivants. La formation du Pléistocène est la plus récente des formations géologiques et, en un lieu donné, toutes les espèces peuvent être encore vivantes, tandis qu’à un autre endroit, dans des formations plus anciennes, jusqu’à 25 % d’espèces peuvent être éteintes. Dans le Pliocène, encore plus ancien, moins de la moitié des formes sont vivantes, et dans la formation immédiatement inférieure, le Miocène, seulement 20 à 40 % sont des espèces actuelles. Enfin, dans la plus ancienne des formations du Cénozoïque, l’Éocène, on trouve rarement plus de 5 % d’animaux à coquille vivants. Autrement dit, la vie actuelle se fond progressivement dans celle du passé géologique, cette transition étant la plus douce chez les invertébrés marins, tandis que peu d’espèces vivantes [ p. 24 ] de vertébrés terrestres sont connues dans les formations géologiques. Cela s’explique par le fait que l’évolution a été plus lente chez les invertébrés marins et les plantes terrestres, et plus rapide chez les mammifères terrestres.
¶ Ce que les fossiles nous apprennent
Les fossiles ne sont ni des anomalies de la nature, ni de simples vestiges fortuits d’êtres autrefois vivants ; ils constituent des archives géologiques essentielles qui ont permis de reconstituer une grande partie de l’histoire de la Terre. Ces archives révèlent : (1) le cours de l’évolution organique, ainsi que la répartition géographique des plantes et des animaux ; (2) la séquence des temps géologiques, ou chronologie ; et (3) la nature de l’environnement des fossiles, qu’ils aient vécu en milieu marin, en eau douce ou sur la terre ferme, et nous renseignent sur la profondeur et la température des mers et les climats terrestres.
La première valeur mentionnée étant d’une importance capitale en paléontologie pure et en biologie générale, il n’est pas nécessaire de la traiter en détail dans cet ouvrage. La valeur héréditaire des fossiles est cependant d’une grande importance en géologie historique et mérite, par conséquent, d’être étudiée avec attention. Toute vie a constamment évolué, non seulement dans sa forme spécifique, mais aussi, en règle générale, dans des directions bien définies, des structures les plus simples aux plus complexes, et souvent dans le sens inverse. C’est pourquoi les fossiles sont d’une valeur inestimable pour reconstituer l’histoire passée de la Terre.
La valeur temporelle des fossiles a été découverte par William Smith et publiée entre 1816 et 1820. Il affirmait que « chaque strate contenait des fossiles organisés qui lui étaient propres », mais cette théorie ne considère que la signification des fossiles comme « témoins de la création » et comme « signes classifiés permettant de reconnaître les formations géologiques ». C’est le domaine de la paléontologie ancienne. La paléontologie supérieure ou comparative, telle que développée par Cuvier (1800-1832), Deshayes, Lyell et Lonsdale, étudie les relations entre les fossiles, avec ceux qui les ont précédés et avec leurs successeurs. Elle traite de l’histoire des organismes et peut ainsi trouver dans les fossiles eux-mêmes des preuves de l’ordre de succession des roches qui les contiennent (H.S. Williams).
Habitats. — Chaque espèce des règnes végétal et animal possède un milieu spécifique, appelé habitat, qui peut être la terre ferme, les rivières et les lacs, ou les mers et les océans. De plus, la température varie entre les pôles et l’équateur, et les organismes sont donc adaptés aux climats froids, tempérés ou tropicaux. Toutes ces différences d’habitat se reflètent dans les fossiles, qui sont ainsi de précieux témoins des climats et [ p. 25 ] des types d’environnements passés. Bien sûr, les organismes terrestres peuvent être emportés vers des milieux aquatiques, mais dans ce cas, on trouvera des traces de ce mélange accidentel, ainsi que la proximité de la terre ferme avec le lieu d’enfouissement.
Fossiles indicateurs. — Comme tous les organismes vivants, à l’instar des individus, ont une durée de vie, généralement courte à l’échelle géologique, il s’ensuit que, puisque les espèces et les genres évoluent constamment, ils constituent en quelque sorte des indices ou des indicateurs de leur période d’existence. Certains fossiles sont de meilleurs indicateurs que d’autres et peuvent être utilisés avec précision pour corréler les strates d’un âge donné d’un lieu à l’autre, voire d’un continent à l’autre.
Faciès fossiles. — Il a été démontré que, puisque toute vie, présente et passée, est adaptée à un environnement particulier, il s’ensuit que les espèces habituées aux marais ou aux eaux douces, aux terres émergées ou aux climats froids, tempérés ou chauds, porteront, même parmi les fossiles, les traces de leurs anciens habitats. De plus, les organismes marins vivant sur des fonds vaseux seront généralement différents de ceux habitués aux fonds sableux ou calcaires. Les formes nageant ou flottant librement, en revanche, peuvent être trouvées dans tous types de dépôts sédimentaires, puisqu’à leur mort, elles se déposent sur n’importe quel substrat. On dit que ces différents habitats impriment leur faciès (environnement sableux, vaseux ou calcaire) aux organismes d’un lieu et d’une époque donnés. Il est vrai que certaines espèces peuvent vivre sur n’importe quel type de fond, mais en règle générale, chaque espèce est dépendante d’un type de fond marin spécifique et y passe sa vie. Les bivalves et les limules préfèrent la vase ou le sable, tandis que les crabes vivent sur tous types de fonds marins et, la nuit, viennent même à terre à la recherche de nourriture.
Évolution et superposition. — Pour déterminer le cours de l’évolution, il est nécessaire d’étudier les fossiles dans l’ordre de leur apparition au sein d’une série de roches donnée, en observant leur présence dans les strates. Ceux des roches inférieures sont naturellement plus anciens que ceux des strates supérieures ou superposées. L’ordre de superposition est généralement déterminé dans les régions où la croûte terrestre n’a pas été perturbée, car les couches de roches s’y succèdent, déposées par l’eau (voir frontispice). En revanche, lorsque les strates sont faillées ou déformées en montagnes, l’ordre naturel de la séquence stratigraphique est parfois très difficile à établir. Cependant, après un siècle de recherches, de nombreuses connaissances ont été acquises sur la séquence évolutive des organismes, et ces connaissances, établies dans de nombreux pays, permettent désormais de déterminer la séquence stratigraphique.
[ p. 26 ]
¶ Comment les fossiles sont préservés
Les fossiles se trouvent toujours dans les roches sédimentaires ou stratifiées déposées dans les étendues d’eau ou sur les terres par l’action des vents. Un autre type de strates sédimentaires susceptibles de contenir des fossiles est constitué de cendres volcaniques qui, lors d’éruptions, sont projetées haut dans l’atmosphère puis transportées par les vents sur des distances plus ou moins longues au-dessus des terres ou des mers, ensevelissant toute forme de vie. C’est ainsi que la station balnéaire romaine de Pompéi fut ensevelie sous les cendres du Vésuve en 79 après J.-C. Aujourd’hui, lors des fouilles de la ville, on découvre des empreintes de victimes. En remplissant ces cavités dans les fines cendres avec du plâtre, on obtient des moulages précis des personnes et des animaux tels qu’ils apparaissaient de leur vivant ; on peut en voir au Musée national de Naples. On trouve aussi parfois des ossements de victimes humaines sous une fine coulée de lave, comme à Pedregal de San Angel, près de Mexico. Cependant, on ne trouve pas souvent de fossiles sous ou dans la couche basale des laves et on ne peut donc pas dire qu’en général ils ne se trouvent pas dans les roches ignées.

Tout organisme mort exposé à une température supérieure au point de congélation de l’eau est, en règle générale, immédiatement attaqué par les champignons et bactéries microscopiques omniprésents [ p. 27 ] du règne végétal, ainsi que par les animaux nécrophages, petits et grands, et disparaît rapidement sans laisser de trace de son existence passée. L’oxygène de l’atmosphère participe également à ce processus, et il en va de même pour les organismes aquatiques, bien que dans ce dernier cas la destruction complète soit plus lente. L’effet dissolvant des eaux en mouvement achève, dans la plupart des cas, la destruction même des parties squelettiques les plus dures. Autrement dit, des espèces entières de flore et de faune disparaissent actuellement sous l’influence d’autres facteurs, de l’atmosphère et de l’hydrosphère. Il est probable que plus de 99 % de toute vie ait ainsi disparu. Toute trace organique peut être oxydée et dissoute dans les éléments dont elle provient : l’air, l’eau et la poussière terrestre. Par conséquent, pour qu’un organisme se fossilise, il doit être rapidement recouvert de sédiments, et même alors, seule une empreinte de sa forme extérieure peut subsister. La destruction complète est la règle chez tous les organismes à corps mou, dépourvus de squelette rigide, et il est exceptionnel, parmi eux, de laisser ne serait-ce qu’une empreinte de leur corps. Lorsqu’il existe un squelette, qu’il soit externe, comme les coquilles, ou interne, comme les os des vertébrés, la conservation de ces structures, ou leur simple présence sous forme d’empreintes, dépend de la nature chimique de ces structures, des caractéristiques des sédiments et, enfin, de la composition chimique des eaux présentes dans les roches.

[ p. 28 ]
Fossiles déformés. — La plupart des sédiments subissent une consolidation lors de leur accumulation, sous l’effet de la superposition de charges et surtout de l’expulsion de l’eau, ce qui permet un compactage plus important des matériaux granulaires. Ce compactage est particulièrement marqué pour les mudstones, et les organismes qu’ils contiennent subissent un broyage important et une déformation plus ou moins prononcée. Lorsque les strates sont plissées en chaînes de montagnes, elles sont soumises à des pressions considérables, ce qui entraîne soit un resserrement, soit un étirement des roches. Cette condition est également particulièrement vraie pour les mudstones.
En revanche, les strates autrefois riches en fossiles peuvent perdre toute trace reconnaissable de ces derniers sous l’effet de la chaleur d’importantes intrusions ignées, ou plus fréquemment sous l’effet de fortes pressions lors de déformations crustales à l’origine de chaînes de montagnes, comme décrit au chapitre XIII de la première partie de cet ouvrage. Il s’agit des roches métamorphisées, et la présence ou non de fossiles reconnaissables dépend de la nature des sédiments qui les composent et du degré d’altération qu’ils ont subi.
Nature des parties conservables chez les organismes. — Le squelette des plantes est très rarement composé de silice, cette caractéristique ne se rencontrant que chez les diatomées unicellulaires et microscopiques. Celles-ci sont si petites qu’un pouce cube de tripoli contient 41 milliards d’individus (voir fig., p. 69). Chez les animaux, la silice est également utilisée avec parcimonie et se limite presque exclusivement aux espèces marines. Les diatomées microscopiques et les radiolaires (voir fig., p. 70) l’utilisent abondamment, et parmi les éponges, probablement un peu moins de la moitié possèdent un squelette siliceux. Chez ces éponges, la silice du squelette est sous forme colloïdale (non cristalline, mais semblable à l’opale) et se dégrade donc facilement lors de la décomposition de la matière organique ; c’est pourquoi les éponges vitreuses (voir fig., p. 199) sont rarement conservées à l’état fossile et ne sont communes que dans les strates du Mésozoïque. Chez les diatomées et les radiolaires, cependant, la silice se présente sous forme insoluble et est très rarement détruite au moment du dépôt ou par la suite.
Le carbonate de calcium est largement utilisé par de nombreux invertébrés, mais chez les plantes, il n’est employé librement que par les algues calcaires marines et d’eau douce. Ces algues transforment le sulfate de calcium présent dans l’eau en carbonate de calcium, constituant ainsi leur squelette carbonaté lié par une base organique. Cette sécrétion, appelée conchine, cristallise sous deux formes minérales : l’aragonite (plus dure et plus dense, cristallisant dans le système rhombique et facilement soluble) et la calcite (cristallisant dans le [ p. 29 ] système hexagonal). Il arrive qu’une structure soit entièrement composée d’aragonite ou de calcite, mais généralement les deux formes coexistent chez un même individu, comme par exemple chez les mollusques, où les couches internes nacrées sont d’aragonite et les couches externes porcelanées de calcite. Dans d’autres cas, la calcite prédomine, comme chez les huîtres. Parfois, les granules ou fibres de matière minérale présentes dans les coquilles sont des associations d’aragonite et de calcite. Il est crucial de le savoir, car les structures composées de calcite, ou où ce minéral est prédominant, sont très susceptibles d’être conservées telles qu’elles ont été sécrétées par les organismes. En revanche, les parties sécrétées sous forme d’aragonite se dissolvent très rapidement lors de la décomposition ou sont remplacées par d’autres minéraux sous l’effet des eaux d’infiltration dans les strates. Dans les roches paléozoïques, la disparition des structures d’aragonite est quasi systématique, notamment chez les mollusques. Par contre, dans les roches mésozoïques et cénozoïques, les couches nacrées sont beaucoup plus fréquemment conservées. Les raisons de ce phénomène restent encore inconnues. Le squelette interne des vertébrés, c’est-à-dire les os, est composé en grande partie de phosphate de calcium lié par un composé organique (le collagène).
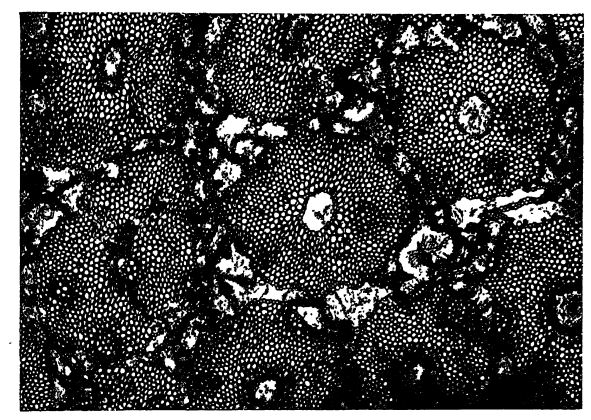
D’autres structures squelettiques, beaucoup plus fines et généralement assez flexibles, sont constituées d’un composé de substances azotées du groupe de l’ammoniac et d’un glucide, appelé [ p. 30 ] chitine. Ce matériau présente une grande résistance aux agents chimiques et est donc difficile à détruire. On le retrouve souvent, sous une forme ou une autre, dans les fossiles. La chitine constitue le squelette externe de nombreux invertébrés, par exemple les arthropodes (limules, trilobites, insectes), auxquels on ajoute souvent des sels de calcium. Une substance similaire est la spongine, qui compose les fibres des éponges de bain ; celle-ci est rarement conservée à l’état fossile. La kératine contient du soufre en plus de l’azote et est très rarement conservée dans les fossiles ; elle constitue les poils, les ongles et les cornes des mammifères, les plumes des oiseaux et les écailles des poissons.
Conservation des parties molles. — Il a été mentionné précédemment que les parties molles des animaux ne se conservent que de façon exceptionnelle. La conservation la plus remarquable d’animaux entiers est cependant celle des éléphants laineux (Elephas primigenius) enfouis dans la toundra (sol gelé, principalement de glace) du nord de la Sibérie, où ils ont été conservés intacts grâce à une technique de conservation naturelle. Il est difficile de dire depuis combien de temps ces carcasses sont emprisonnées dans la toundra, certainement des milliers d’années, et pourtant, lorsque la glace fond, la chair est exposée et avidement dévorée par les chiens des peuples sibériens. Le squelette d’un de ces mammouths, découvert à l’embouchure du fleuve Léna, et un autre provenant de Beresovka, avec la peau et le squelette montés et tous les organes internes conservés dans l’alcool, sont exposés au Musée d’histoire naturelle de Petrograd, en Russie. Un rhinocéros poilu (Rhinoceros tichorhinus) a été découvert en Sibérie, conservé de la même manière, et une conservation similaire, mais bien moins parfaite, a été mise au jour dans les sols gelés du nord-ouest de l’Alaska. Enfin, de larges fragments de peau de rhinocéros ont été retrouvés en excellent état de conservation dans les suintements de pétrole de Galice.
On trouve souvent des moisissures recouvrant l’intégralité de l’enveloppe extérieure d’organismes, même d’animaux au corps extraordinairement mou comme les méduses, dont la substance organique est composée d’au moins 95 % d’eau. Les exemples les plus remarquables proviennent des calcaires jurassiques de Solenhofen, en Allemagne.
Préservation de la couleur. Il est extrêmement rare de trouver parmi les fossiles une trace de leur couleur d’origine. Lorsqu’ils sont conservés dans des calcaires schisteux clairs, l’ancienne coloration peut apparaître sous forme de bandes sombres, comme on en trouve des exemples datant du Champlainien. La couleur nacrée, ou nacre, est cependant souvent préservée, et particulièrement bien, dans les schistes calcaires imprégnés de pétrole. Mais cette couleur est due au jeu de la lumière dans les parties prismatiques ou aragonitiques des coquilles, et non à un pigment présent dans le matériau.
¶ Comment se forment les fossiles
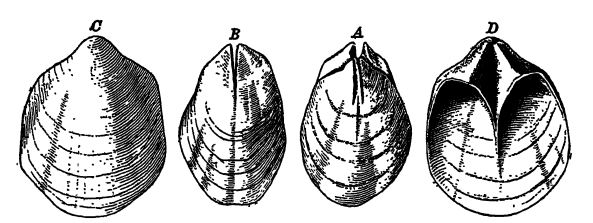
Les fossiles se présentent sous sept formes naturelles différentes, dont trois concernent la substance laissée par les organismes, trois leur forme, et une les deux. (1) La grande majorité des spécimens fossiles conservent plus ou moins la substance dure ou minérale d’origine de la plante ou de l’animal. À cette substance, lors de la minéralisation, une autre substance minérale peut s’être ajoutée dans les interstices organiques, formant ainsi des fossiles perminéralisés. (2) Lorsque la matière [ p. 31 ] minérale d’origine est remplacée par une autre, généralement différente, le substitut conservant la structure microscopique d’origine de l’organisme, ce processus de substitution est appelé histomotase (deux mots grecs signifiant « échange de tissus ») (Fig., p. 29). Dans ce cas, les parties ligneuses des plantes sont souvent conservées et, pour les études, sont aussi pertinentes que les parties similaires des plantes vivantes. (3) Les plantes peuvent être entièrement carbonisées (charbon), leur structure organique d’origine étant alors plus ou moins complètement détruite. Ces fossiles présentent peu d’importance paléontologique, mais une grande valeur économique et sont souvent utilisés comme plans de référence en stratigraphie et en géologie structurale. La forme des organismes, privée de leur substance originelle, peut apparaître dans les roches sous forme de (4) moules, (5) empreintes et (6) moulages naturels (Fig. ci-dessous). Il n’y a pas de différence notable entre moules et empreintes, si ce n’est que ce dernier terme s’applique aux impressions de substances fines, comme des feuilles, etc. Les moulages naturels sont les copies d’organismes obtenues en remplissant les moules des fossiles avec un substitut non cristallisé ; (7) lorsque le matériau de remplacement est un minéral cristallisé, comme la calcite, la pyrite et, plus fréquemment, la silice sous forme de calcédoine, le remplacement est appelé pseudomorphe (de deux mots grecs signifiant « fausse forme ») (Fig., p. 26 ; voir aussi Fig., p. 32).
Le bois fossilisé est présent en quantité presque incroyable au nord et au sud du fleuve Colorado. Ici comme ailleurs, les troncs siliceux gisent à plat dans des dépôts continentaux d’origine fluviale, ou dressés dans des cendres volcaniques. L’agent pétrifiant lors de l’histométabase semble avoir été les eaux alcalines périodiques des climats semi-arides ou arides, contenant en solution la silice qui s’est infiltrée à travers les sables ou les cendres non consolidés lors de leur accumulation. La silice remplace le tissu ligneux, [ p. 32 ] molécule par molécule, et dans la plupart des cas, préserve remarquablement la microstructure des plantes (Fig., pp. 27, 29). Les troncs enfouis dans des dépôts continentaux de climats pluviaux sont dissous et se présentent sous forme de moulages ou de charbon. Le tissu ligneux peut également être remplacé par de l’alcite, mais dans ce cas, le remplacement semble s’être produit sous une couverture d’eau permanente, et peu de temps après l’enfouissement, comme dans le cas des « boules de charbon ». La plupart de ces occurrences rares se trouvent dans des dépôts de marais d’eau douce, et on peut se demander si un tel remplacement a jamais lieu dans les strates marines.
Les traces d’animaux rampant sur les fonds vaseux sous les étendues d’eau, et notamment en mer, sont très fréquemment conservées. Les animaux terrestres laissent aussi souvent leurs traces dans les archives géologiques, mais celles des vertébrés se trouvent le plus souvent dans les argiles rouges ou les sables déposés dans les déserts ou du moins dans les régions arides. On observe fréquemment des traces de reptiles au Carbonifère (Mauch Chunk) et elles sont particulièrement abondantes au Trias.

Évolution géologique des organismes. — Dans les dépôts cénozoïques, on trouve généralement des coquilles marines presque inchangées par l’ajout de matière minérale. De plus, on peut généralement ajouter que plus les organismes sont anciens géologiquement, plus ils sont susceptibles d’être minéralisés. Environ 50 % des fossiles restent dans leur état calcaire d’origine ou ne sont que légèrement perminéralisés. Cette minéralisation a pu se produire par l’infiltration des eaux dans les roches à tout moment après l’enfouissement des fossiles ou lors de l’altération des formations, mais elle a plus vraisemblablement eu lieu au moment de l’enfouissement, lorsque les eaux souterraines du fond marin étaient chargées d’acide carbonique et d’autres acides provenant de la décomposition organique. Pour ces raisons, les squelettes fossiles d’organismes se présentent dans tous les états, de l’état inchangé à des pseudomorphoses complètes, dans la calcite, la dolomite, la silice, la pyrite de fer, voire le plomb, le zinc, etc.
[ p. 33 ]
Là encore, les eaux d’infiltration terrestres ou les eaux souterraines marines peuvent avoir un pouvoir dissolvant limité, emportant avec elles toutes les coquilles organiques. Dans ce cas, leur présence antérieure est attestée par des cavités dans les roches, les moules d’organismes, plus fréquemment observés dans les dolomies et les grès. Ces moules sont souvent d’une netteté remarquable, permettant d’en réaliser des moulages ou des empreintes d’une grande précision. En revanche, il arrive souvent que les moules, notamment ceux présents dans les dolomies, soient recouverts d’une couche de matière minérale cristallisée, auquel cas les fossiles sont presque toujours sans valeur. Bien que les moules soient volumineux à collecter, il convient néanmoins de les recueillir et de préserver également les empreintes internes, correspondant au remplissage des cavités des organismes. Les empreintes, plus fréquentes dans les schistes, sont les impressions laissées dans la boue par les parties externes des plantes et des animaux.
¶ Où trouve-t-on des fossiles
Il convient de rechercher les fossiles principalement dans les strates régulières d’origine marine, plus ou moins calcaires ou magnésiennes, et en moindre mesure dans les schistes et grès rouges. Dans presque tous les lits rouges, à l’exception des calcaires rouges, la présence de fossiles est exceptionnelle, car les plantes et les animaux ont été oxydés et dissous lors de la sédimentation subaérienne. Ce sont donc les lits verts, bleus, gris, noirs et, plus tard, jaunes (après oxydation) qui sont susceptibles de contenir des fossiles. Cependant, les schistes et grès rouges peuvent également renfermer des fossiles tels que des empreintes ou des traces d’animaux, très rarement des ossements de vertébrés, et encore plus rarement des empreintes de plantes ou des bois calcédés. Dans les dépôts d’eau douce, les fossiles sont généralement très rares, mais après le Silurien, ces lits peuvent contenir des restes de plantes et de vertébrés.
Le hasard de l’enfouissement a beaucoup à voir avec les différences marquées entre les types de traces organiques, car un animal vivant dans la mer peu profonde et surtout dans ou sur le fond marin se trouve dans l’endroit le plus favorable pour être enseveli, tandis que seuls les animaux terrestres peuvent laisser des fossiles qui vivent dans, ou dont les corps sont emportés dans, une zone où s’accumulent des sédiments ; les fossiles terrestres les plus abondants étant les restes de ceux qui vivent dans les plaines deltaïques ou en partie dans les rivières.
De toutes les roches stratifiées, les schistes argileux représentent 80 %, les grès environ 15 % et les calcaires 5 % (voir partie I, p. 281). C’est dans les calcaires, « le sel conservateur du monde géologique » (Hugh Miller), que les fossiles sont presque toujours présents en abondance. Cependant, [ p. 34 ] cela ne signifie pas que pour chaque mètre de calcaire, neuf schistes argileux et grès sont dépourvus d’organismes. Le carbonate de calcium est largement disséminé dans les strates, et tous les schistes argileux calcaires sont susceptibles de contenir de beaux fossiles qui se libèrent par érosion ; même les grès calcaires renferment généralement des organismes en quantité non négligeable. De plus, les épaisses formations de schistes argileux verts et bleus présentent souvent de minces lits de calcaire ou de grès impurs, et c’est dans ces couches qu’il faut rechercher des fossiles.
Les conglomérats peuvent être riches en fossiles, mais il convient alors d’être particulièrement vigilant pour bien séparer les spécimens de chaque galet, car leur âge peut être très différent. Il ne faut pas non plus mélanger les fossiles de la matrice avec ceux des galets, sous peine de créer une association chronogénétique impossible. En revanche, les brèches et les conglomérats intraformationnels contiennent les mêmes espèces que la matrice qui les lie, car ils se sont tous formés simultanément.
Comment les fossiles sont collectés ? — La recherche de fossiles ne doit pas être précipitée, car c’est un travail de longue haleine. On croit souvent, à tort, que les paléontologues « sentent » la présence de fossiles, car ils ne sont souvent découverts qu’après une recherche patiente.
Sur toute surface de roches stratifiées affleurant, des fossiles peuvent apparaître ; il convient donc d’examiner attentivement la surface à la recherche de spécimens détachés. Ces fossiles libres sont les plus recherchés, car leur nettoyage est plus facile et, de plus, ils présentent toutes les parties de l’individu. Il faut également fouiller toutes les argiles rouges résiduelles, car elles regorgent souvent de beaux spécimens libres, généralement conservés dans la silice sous forme de pseudomorphes.
Lorsqu’on trouve des fossiles non fixés, il faut remonter à la couche dont ils proviennent et noter la présence d’autres fossiles encore en place. Les meilleurs endroits pour trouver des fossiles sont naturellement les affleurements calcaires, qu’il convient de casser par endroits à l’aide d’un marteau. Parfois, une fine couche de roche renferme un amas de fossiles ; lorsque l’altération les a détachés au point qu’ils tombent librement sous les coups de marteau, la méthode la plus simple pour les collecter consiste à prélever un morceau de 15 à 30 cm de diamètre. À ce propos, il convient d’ajouter que les roches altérées livrent beaucoup plus facilement des fossiles que les roches non altérées.
Les fossiles de nature siliceuse et partiellement altérés dans le calcaire doivent être ramassés en grande quantité, pour être traités en laboratoire avec de l’acide chlorhydrique dilué (Fig., p. 26). On peut facilement tester sur le terrain la résistance des matériaux à ce traitement à l’aide d’un couteau de poche ; si la lame ne raye pas le fossile mais laisse une marque noire, l’expérience peut être réalisée. Bien sûr, lorsque les fossiles sont inclus dans des cherts ou plus ou moins entourés de silice amorphe, il est impossible de les extraire.
Le géologue de terrain, lorsqu’il collecte des fossiles destinés à l’étude du paléontologue, fera bien de se rappeler que la valeur du travail de ce dernier dépend largement du nombre de spécimens qu’il peut étudier ; il convient donc de recueillir [ p. 35 ] autant de matériel que les circonstances le permettent. Ce n’est cependant pas tant la quantité de spécimens que la diversité des espèces qui intéresse le paléontologue, car on sait que différentes formes présentent des valeurs chronogénétiques très différentes.
¶ Lecture collatérale
A. M. Davies, Introduction à la paléontologie. Londres (Murby), 1920.
W. Deecke, La fossilisation. Berlin (Bomtraeger), 1923.
H. L. Hawkins, Paléontologie des invertébrés, une introduction à l’étude des fossiles. Londres (Methuen), 1920.
Charles Schuchert, Instructions pour la collecte et la préparation des fossiles. États-Unis
Musée national, Bulletin 39, Partie K, 1895.
H. W. Shimer, Introduction à l’étude des fossiles. New York (Macmillan), 1914.
C. A. White, La relation de la biologie à l’investigation géologique. Rapport annuel du Musée national des États-Unis pour 1892, 1894, pp. 245-368.
| II. Les organismes, leur composition, leur structure et leur classification | Page de titre | IV. L'évolution, le changement constant des êtres vivants |