| XLIII. L'évolution des chevaux et des autres mammifères ongulés | Page de titre | XLV. Le Pléistocène et le dernier climat glaciaire |
[ p. 640 ]
Parmi les animaux du Cénozoïque, aucun groupe n’a présenté une évolution et une répartition plus spectaculaires que les éléphants, ces imposants mammifères à trompe. De nos jours, ils dominent encore les rangs des mammifères terrestres par leur taille, leur morphologie singulière et le volume de leur cerveau. Il n’existe cependant que deux espèces d’éléphants : les éléphants d’Afrique, plus grands et aux grandes oreilles, dont certains atteignent un poids d’environ 8 tonnes et une hauteur au garrot de près de 4 mètres ; et l’espèce, un peu moins lourde et aux oreilles plus petites, que l’on trouve en Inde et en Asie centrale.
Les mammifères ressemblant à des éléphants sont techniquement appelés Prohoscidea, la trompe étant leur caractéristique la plus marquante. Il s’agit en réalité d’un nez, de narines et d’une lèvre supérieure très allongés, formant un appendice musculaire très flexible et puissant à la tête, remplissant de nombreuses fonctions, mais servant principalement à collecter la nourriture et l’eau et à les transporter jusqu’à la bouche, bien au-dessus du sol. La trompe est également un organe sensoriel très développé ; tenue haute au-dessus de la tête, elle permet de détecter l’eau et les prédateurs au loin. La vue, en revanche, est beaucoup moins développée. Le cou des proboscidiens étant très court, leur tête très lourde et leurs pattes longues et robustes, la longue trompe a dû se développer au cours de l’évolution du groupe pour permettre à la lèvre supérieure d’atteindre le sol afin de se nourrir.
La tête des éléphants est non seulement volumineuse, mais aussi particulière par sa grande hauteur par rapport à sa largeur ; en d’autres termes, elle est de type bulldog. La hauteur du crâne est une adaptation qui offre une plus grande surface musculaire et, par conséquent, un meilleur appui aux muscles du cou qui soutiennent la tête et la trompe. La partie supérieure du crâne est, quant à elle, nettement cellulaire. La plus grande transformation des proboscidiens s’est opérée au niveau de la tête et de la trompe, débutant chez des formes à tête allongée et à trompe très courte et progressant graduellement vers le type actuel de crâne haut et de longue trompe.
Le cerveau des éléphants est volumineux, environ deux fois plus gros que celui de l’homme, et sa structure est primitive : le cerveau antérieur (cerveau) ne recouvre pas le cerveau postérieur (cervelet) comme chez la plupart des autres animaux.
Aujourd’hui, à l’état sauvage, les éléphants sont confinés aux forêts et jungles d’Asie du Sud-Est et d’Afrique [ p. 642 ] centrale. [ p. 641 ] Au Pléistocène, cependant, leur répartition était quasi mondiale et ils vivaient sous tous les climats, même les plus froids de Sibérie et d’Alaska, jusqu’à l’océan Arctique. On pense que les formes les plus primitives vivaient dans les cours d’eau et les lacs, c’est-à-dire qu’elles étaient amphibies, et ce n’est qu’après leur colonisation des forêts et des plaines herbeuses que leur répartition s’est étendue à une zone aussi vaste.

Les restes de proboscidiens du Pléistocène sont fréquemment découverts près de la surface, dans les tourbières et les marais. Le comté d’Orange, au sud-est de l’État de New York, a livré pas moins de trente et un squelettes. Ce sont généralement les dents ou les os des membres les plus importants qui sont mis au jour, et leur découverte au Haut Moyen Âge a alimenté les légendes de peuples anciens géants. On a aussi cru qu’il s’agissait des restes des « bêtes gigantesques qui faisaient trembler la terre » utilisées par les Romains lors de leurs invasions de l’Europe occidentale. C’est le grand naturaliste Cuvier qui, au XIXe siècle, a démontré avec succès que les ossements d’éléphants trouvés en Europe occidentale et méridionale appartenaient à des espèces totalement différentes de celles qui vivent actuellement. Il nomma l’une d’elles « mastodonte », ce qui signifie « dent-mamelon », en référence à la forme très crêtée ou conique de la surface broyeuse de ses dents, qui est la forme la plus courante.
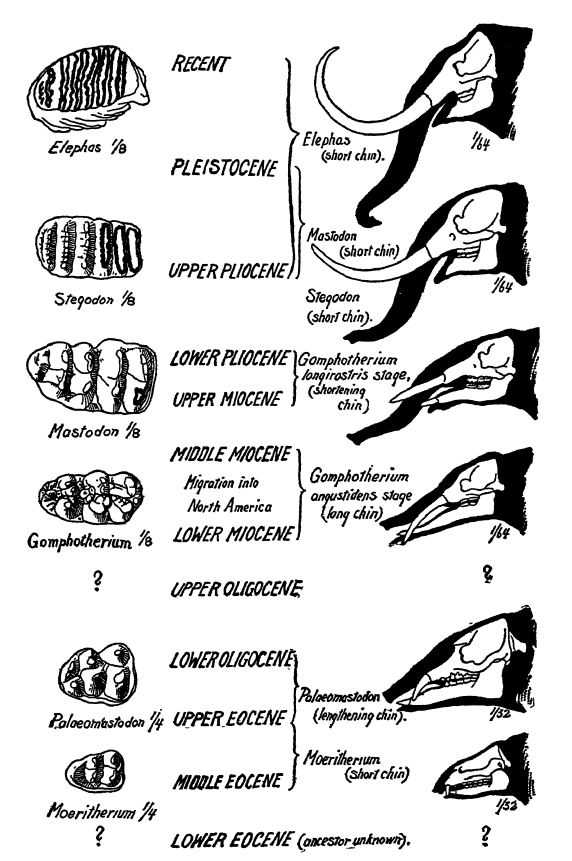
Origine. — En 1901, Beadnell et Andrews ont décrit trois espèces de proboscidiens de l’Éocène supérieur et de l’Ogocène d’Égypte (Maeritherium, Palaeomastodon et Phiomia). Les première et dernière espèces ont depuis été découvertes en Asie du Sud. L’origine de ce groupe se situe donc dans l’Éocène inférieur d’Eurasie ou d’Afrique.
Évolution. — Au lieu de n’avoir que deux souches principales d’animaux à tronc, comme on le supposait généralement, Osborn identifie désormais douze lignées de formes évolutives.
Ces animaux se regroupent en quatre branches principales : (1) Les Thémoérithères, présents dans l’Éocène supérieur et l’Oligocène inférieur d’Égypte ; il s’agissait de petits animaux dépourvus de trompe, vivant dans les marais et appartenant peut-être ou non à la famille des proboscidiens. (2) Les Dinothères spécialisés, formes amphibies, dont certains atteignaient la taille des plus grands mastodontes, dépourvus de défenses supérieures mais dotés de défenses inférieures grandes, robustes et recourbées vers le bas, contrairement à tous les [ p. 644 ] autres groupes de proboscidiens, [ p. 643 ] et possédant une trompe sans aucun doute imposante et massive ; on les trouve dans le Miocène et le Pliocène d’Europe, d’Asie et d’Afrique. (Voir Fig. ci-dessus.) (3) Les mastodontes. Un groupe très diversifié, la principale lignée de proboscidiens brouteurs, originaire de PalcBomastodon de l’Oligocène inférieur d’Égypte, à distribution mondiale au Miocène et surtout au Pléistocène, et disparu lors de la dernière période glaciaire. (4) Les éléphants, ou groupe de proboscidiens herbivores, originaires du Pliocène, ayant également atteint une dispersion mondiale au Pléistocène, et représentés dans le monde actuel par les éléphants d’Afrique et d’Inde.
Le plus ancien animal connu de type proboscidien est Maeritherium, datant de l’Éocène supérieur et de l’Oligocène inférieur d’Égypte. De la taille d’un tapir, il était dépourvu de trompe, même embryonnaire. Bien qu’il ait d’abord été considéré comme l’ancêtre commun de tous les mammifères à trompe, sa relation avec les proboscidiens reste aujourd’hui incertaine. Quoi qu’il en soit, il illustre l’évolution possible de ces animaux. Sa tête était longue et étroite, son museau plutôt court, sans aucune trace de trompe. Sa caractéristique la plus intéressante était cependant la présence de défenses issues des deuxièmes incisives, les autres dents antérieures demeurant petites. Les défenses supérieures, encore à l’état embryonnaire, étaient très proéminentes et pointaient fortement vers le bas, tandis que celles de la mâchoire inférieure étaient presque couchées, avec une légère inclinaison vers le haut (Fig., p. 641).
Le Paléomastodon, de l’Oligocène inférieur d’Égypte, le plus ancien proboscidien connu, était beaucoup plus petit et plus malléable. De la taille d’un tapir, son museau étroit était plus allongé que celui du Moeritherium, plus ancien, et il possédait un museau flexible et bien développé plutôt qu’une trompe. Les défenses du crâne étaient plus longues, comprimées et orientées vers l’extérieur ; celles des mâchoires, bien que plus grandes, pointaient droit devant. Toutes les dents broyeuses (prémolaires et molaires) étaient présentes et fonctionnelles simultanément, contrairement aux proboscidiens plus récents (Fig., p. 641). Ses membres ressemblaient à ceux des éléphants.
Les nombreuses espèces de mastodontes du Pliocène et du Miocène étaient plus petites que celles du Pléistocène. Certaines possédaient deux défenses (Dibelodon), tandis que d’autres, à museau allongé, en avaient quatre (Tetralophodon, Trilophodoriy, Gomphotherium). C’est à partir des formes à museau allongé que s’est développé pleinement le genre des proboscidiens. En Amérique du Nord, l’espèce la plus connue est le mastodonte américain (Mammut americanum), dont on trouve des squelettes de la Floride jusqu’en Alaska, du Connecticut à la Californie, et du centre de la Russie jusqu’à la Sibérie.
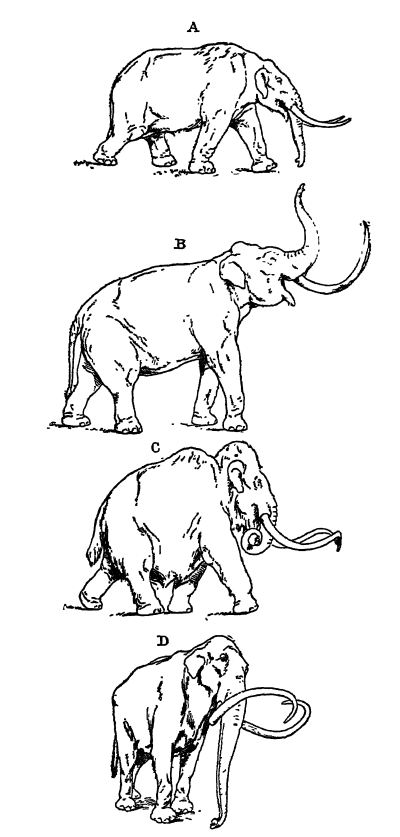
Il existe au moins une douzaine d’espèces d’éléphants éteintes. Selon Matthew, certaines sont plus proches de l’éléphant d’Asie actuel (Elephas), d’autres de l’éléphant d’Afrique (Loxodon), tandis que d’autres encore (Stegodon) présentent [ p. 645 ] des caractéristiques intermédiaires entre ces derniers et les mastodontes plus anciens. Trois espèces ont vécu au Pléistocène en Amérique du Nord : le mammouth à fourrure, l’éléphant de Colombie et l’éléphant impérial, tous de très grande taille. Parmi celles-ci, le mammouth (Elephas primigenius) est la plus connue. Son aire de répartition était très vaste, s’étendant non seulement à l’Amérique, mais aussi à l’Europe et à l’Asie. En Europe, on trouve des restes de ces animaux jusqu’en Espagne et en Italie, et en Amérique, de la Caroline du Nord et de la Californie vers le nord. Le terme « mammouth », cependant, ne signifie pas de taille gigantesque mais vient du mot tatar mama’ntee, signifiant souris de terre, un habitat attribué à ces animaux par une légende chinoise qui dit qu’ils vivaient sous terre et périssaient lorsqu’ils arrivaient à la lumière du jour (voir Fig. C, p. 643).
En résumé, l’évolution des Proboscidiens, selon Lull, se caractérise par : une augmentation de la taille et le développement de membres en forme de piliers pour supporter leur poids considérable ; une augmentation de la taille et de la complexité des dents, entraînant une diminution de leur nombre et le développement d’une méthode particulière de succession dentaire ; la perte des canines et de toutes les incisives, à l’exception de la deuxième paire, dans les mâchoires supérieure et inférieure, et leur transformation en défenses ; un allongement progressif de la symphyse ou union des mâchoires inférieures pour renforcer et soutenir les défenses inférieures lors du creusement, aboutissant au Gomphothère ; le raccourcissement apparemment soudain de cette symphyse suite à la perte des défenses inférieures et l’augmentation compensatoire de la taille et la modification de la courbure de celles de la mâchoire supérieure.
L’augmentation de la masse et de la hauteur, conjuguée au raccourcissement du cou imposé par le poids croissant de la tête et de ses imposantes défenses, a nécessité le développement d’une lèvre supérieure préhensile qui a progressivement évolué en une trompe pour la recherche de nourriture. L’allongement de la mâchoire inférieure implique un allongement similaire de la trompe afin que celle-ci puisse atteindre des objets situés au-delà des défenses. La trompe n’a cependant atteint son utilité maximale que lorsque le raccourcissement de la mâchoire, en supprimant son support inférieur, l’a laissée pendante, comme chez l’éléphant actuel.
Migration. — Le Paléomastodon, ou ses descendants, les tétrabélodons à long museau, ont traversé l’Afrique par un pont terrestre méditerranéen reliant Tunis, la Sicile et l’Italie à l’Eurasie. Ce pont existait déjà à l’Oligocène. De là, leur expansion s’est faite vers l’ouest, en Grande-Bretagne, et peut-être même en Amérique du Nord, via un pont terrestre du Miocène inférieur reliant l’Amérique et l’Europe par les îles Shetland, l’Islande et le Groenland, et vers l’est à travers l’Asie, pour finalement atteindre Nome en Alaska par la Sibérie. Au Pliocène, les éléphants se sont finalement [ p. 646 ] répandus d’Amérique du Nord en Amérique du Sud. Ainsi, les proboscidiens ont été de grands voyageurs à travers le monde, égalés seulement par les chevaux et surpassés seulement par l’homme.
Les éléphants contemporains de l’homme. — En Europe occidentale, d’excellentes preuves attestent de la connaissance qu’avait l’homme du mammouth velu, puisqu’à la fin du Pléistocène, il en gravait l’image sur des os et de l’ivoire et la peignait sur les parois des grottes. Il ne fait aucun doute que l’homme, le mammouth velu et d’autres proboscidiens ont coexisté avec lui durant tout le Pléistocène en Eurasie. En Amérique, il semble que les constructeurs de tumulus connaissaient également les éléphants ou les mastodontes, puisqu’ils ont bâti des tumulus dont la forme leur rappelait celle de ces animaux.
¶ Lecture collatérale
R. S. Lull, L’évolution de l’éléphant. American Journal of Science, 4e série, vol. 25, 1908, p. 169-212. Rapport annuel de la Smithsonian Institution pour 1908, 1909, p. 641-675.
R. S. Lull, Évolution organique, Chapitre 34. New York (Macmillan), 1917.
W. D. Matthew, Mammouths et mastodontes. American Museum of Natural History, Guide Leaflet Series, n° 43, 1915.
H. F. Osborn, Évolution, phylogénie et classification des Mastodontoidea. Bulletin de la Société géologique d’Amérique, vol. 32, 1921, pp. 327-332.
H. F. Osborn, L’évolution, la phylogénie et la classification des Proboscidea. American Museum Novitates, n° 1, 1921.
W. B. Scott, Une histoire des mammifères terrestres de l’hémisphère occidental, Chapitre 10. New York (Macmillan), 1913.
| XLIII. L'évolution des chevaux et des autres mammifères ongulés | Page de titre | XLV. Le Pléistocène et le dernier climat glaciaire |