| XLII. L'évolution des mammifères et l'émergence de la pensée au Cénozoïque | Page de titre | XLIV. L'évolution des éléphants |
[ p. 624 ]
¶ Les chevaux
Pour démontrer la véracité de la théorie de l’évolution, les chevaux, plus que tout autre organisme, illustrent parfaitement la mise en œuvre de cette doctrine par le biais de la sélection naturelle et de l’adaptation à l’environnement. Ils sont les « animaux emblématiques » de l’évolution, leur histoire, remontant à des millions d’années, étant désormais bien connue. Ainsi, la célèbre collection de l’université Yale, rassemblée par le professeur Marsh, a largement contribué à établir la véracité du darwinisme (voir figure, p. 627).
Il y a de nombreuses années, Huxley affirmait que le cheval devait descendre d’un quadrupède possédant cinq doigts ou orteils complets à chaque pied et dont les os de l’avant-bras et de la jambe étaient complets et séparés. De plus, il soutenait que si le cheval avait ainsi évolué et que les vestiges des différentes étapes de son évolution avaient été conservés, ils devraient nous présenter une série de formes où le nombre de doigts diminue progressivement, les os de l’avant-bras et de la jambe acquièrent une conformation équine, et la forme et la disposition des dents se rapprochent peu à peu de celles des chevaux actuels. Depuis l’époque d’Huxley, presque tous les chaînons manquants de l’évolution du cheval ont été découverts, et nulle part ailleurs cette histoire n’est aussi complète que dans les formations cénozoïques des Grandes Plaines des États-Unis.
Le cheval est le plus utile et le plus beau des animaux domestiques et a joué un rôle majeur dans le développement de la civilisation humaine. Dès les origines de l’humanité, l’homme se nourrissait de chevaux. À Solutre, en Saxe-et-Loire, un amas d’ossements équins recèle des restes estimés à 80 000 individus. Plus tard, le cheval devint le principal moyen de transport et la bête de somme indispensable à l’agriculture et à la guerre. Il compte également parmi les machines vivantes les plus performantes et les plus rapides, comme l’homme aime à le démontrer avec les courses de chevaux. Migrants sur tous les continents et capables de s’adapter à des environnements [ p. 625 ] variés, des climats les plus chauds aux plus arctiques, les chevaux n’ont eu que deux égaux : les éléphants et l’homme. À l’état sauvage, les chevaux sont aujourd’hui confinés aux vastes plaines arides d’Asie centrale et d’Afrique, car les mustangs et les broncos d’Amérique du Nord et du Sud descendent de chevaux domestiques retournés à l’état sauvage depuis l’époque des explorateurs espagnols.
Caractéristiques distinctives. — La famille des équidés (Equidse, du grec Equus caballus, le cheval vivant) comprend les chevaux, les zèbres et les ânes. Ils appartiennent aux ongulés à doigts impairs (Perissodactyle), dont l’axe du pied se situe au niveau du troisième doigt. Les chevaux se caractérisent par leurs pieds très longs et fins, chacun composé d’un seul doigt fonctionnel, le troisième orteil. Le sabot est l’équivalent de l’ongle ou de la griffe du troisième doigt ou orteil chez les autres animaux. Les chevaux marchent donc sur l’extrémité du doigt, en fait sur l’ongle du troisième doigt, le poignet étant ce que les cavaliers appellent le « genou » et le talon le « jarret ». Comme le troisième orteil de chaque membre supporte tout le poids du cheval, il est nécessairement beaucoup plus gros que chez les animaux où le poids est réparti sur plusieurs doigts. De chaque côté du doigt fonctionnel, c’est-à-dire le « canon », se trouve un os fin appelé « os spléniforme ». Il s’agit des vestiges du deuxième et du quatrième orteil, parmi les cinq orteils d’origine chez les ancêtres des chevaux (Fig., p. 627). Aussi curieux que cela puisse paraître, certains chevaux domestiques naissent avec deux ou trois orteils surnuméraires à un ou plusieurs pieds, et César aurait monté un tel cheval au combat. Toute la structure du cheval est parfaitement adaptée à la course rapide, et chaque partie de son squelette a été modifiée et spécialisée à cette fin.
Les dents des chevaux sont aussi particulières que leurs pieds monodentés (Fig. ci-dessus). Les molaires sont de longs prismes [ p. 626 ] carrés qui poussent à partir de la gencive aussi vite qu’elles s’usent au niveau de la couronne. Cette usure est accélérée par la silice contenue dans l’herbe consommée et par le sable ingéré lors de l’alimentation. La surface de mastication présente une structure complexe et particulière : des bords d’émail dur, entre lesquels se trouvent des espaces plus tendres composés de dentine et d’une substance appelée cément, semblable à la dentine par sa composition, mais plus tendre et formée différemment. La dentine se forme sur les faces internes de l’émail lorsque la dent est encore incluse dans la mâchoire ; le cément se dépose sur les faces externes de l’émail après l’éruption de la dent et avant son apparition au-dessus de la gencive.

Évolution. — On retrouve la famille des chevaux jusqu’au début du Cénozoïque, sans rupture majeure. Lorsque les petits chevaux à quatre orteils (Eohippus), pas plus grands qu’un petit chien, apparurent dans l’ouest de l’Amérique du Nord au début de l’Éocène (Wasatch), le terrain était bien plus proche du niveau de la mer qu’aujourd’hui et le climat, d’abord marqué par des hivers rigoureux, devint rapidement chaud et tempéré tout au long de l’année. Pendant longtemps, [ p. 628 ] les saisons [ p. 627 ] furent très similaires et le climat tropical et suffisamment humide pour favoriser le développement de vastes forêts, du moins au-dessus des Cordillères (monts Laramiens). Sur les Grandes Plaines, en revanche, le climat était plus sec et de vastes plaines herbeuses s’étendaient. Au Miocène, tout l’ouest de l’Amérique du Nord commença à s’élever, atteignant son altitude maximale au Pléistocène. Parallèlement à ces changements topographiques, le climat devint plus froid, plus sec, puis glacial. Face à tous ces changements environnementaux, les chevaux se sont adaptés ou ont migré vers des habitats plus favorables, passant ainsi des formes plus petites à nombreux doigts aux formes plus grandes, à moins de doigts, plus rapides et plus intelligentes.
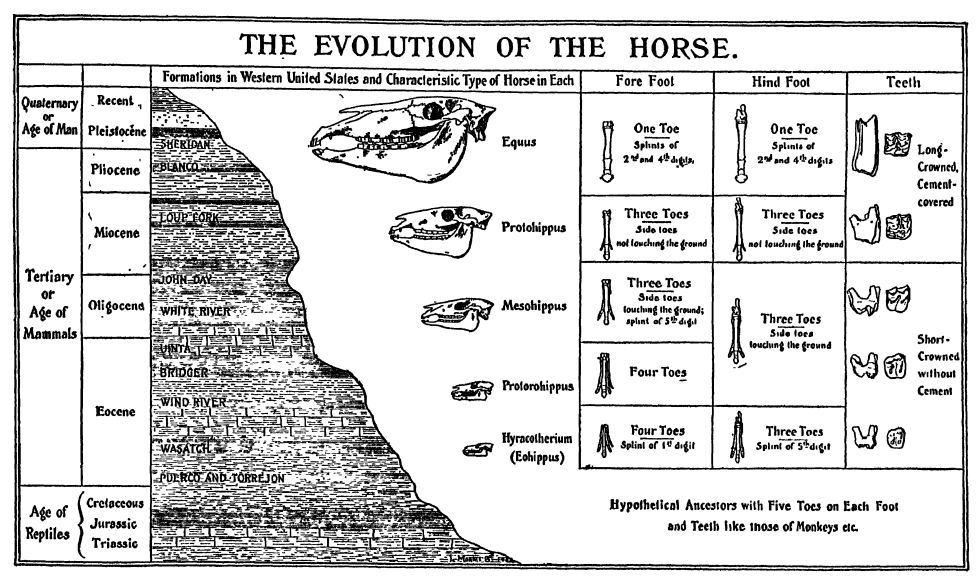
Au départ, les chevaux polydactyles broutaient dans les forêts où ils étaient des proies faciles pour les carnivores de l’époque. Mais avec la disparition progressive des forêts et l’apparition des plaines herbeuses plus sèches, ils se sont dispersés dans ces plaines pour s’abriter, et c’est là qu’ils ont développé une vitesse croissante. L’allongement de la partie inférieure des membres et le développement de la capacité à se cabrer rapidement ont entraîné la perte progressive, par désuétude, des orteils supplémentaires, ainsi qu’une transformation tout aussi remarquable de la dentition, passant d’une dentition à couronne courte (adaptée au broutage) à une dentition à couronne longue (adaptée au pâturage). De cette manière, les chevaux sont devenus l’un des animaux les plus spécialisés et adaptés à un environnement particulier.
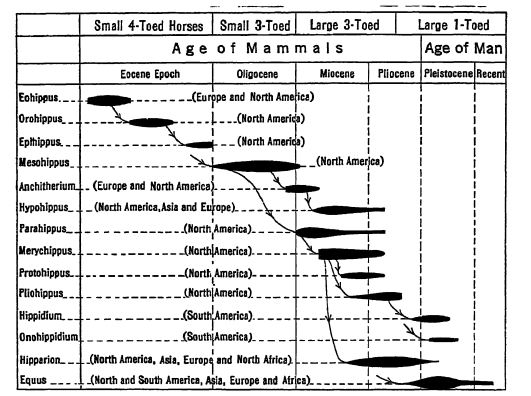
L’origine de la famille des chevaux demeure inconnue. Les premiers chevaux apparaissent à peu près simultanément, et au même stade d’évolution, en Europe occidentale et en Amérique du Nord. En Europe, ils disparaissent rapidement (Éocène), tandis qu’en Amérique du Nord, ils constituent leur principal centre de développement durant tout le Cénozoïque. Curieusement, bien que les chevaux aient été présents tout au long du Pléistocène en Amérique du Nord et du Sud, ils avaient tous disparu avant l’arrivée des premiers humains. Nos chevaux sauvages actuels sont retournés à l’état sauvage, c’est-à-dire qu’ils descendent d’ancêtres domestiqués, tandis que ceux d’Asie, d’Afrique et d’Europe sont les descendants de chevaux du Miocène inférieur qui se sont répandus d’Amérique du Nord en Sibérie en passant par l’Alaska. À la fin du Miocène, les chevaux nord-américains ont gagné l’Amérique du Sud.
Matthew distingue douze étapes dans l’évolution de la famille des chevaux, présentes dans autant de formations géologiques différentes et successives. Outre la lignée principale qui a donné naissance aux chevaux, ânes et zèbres modernes, il existe plusieurs branches collatérales qui n’ont pas laissé de descendance.
Scott résume ainsi le long et remarquable développement de la famille des chevaux : (1) Leur taille a augmenté de façon quelque peu fluctuante, passant progressivement des petits chevaux de l’Éocène inférieur, de la taille d’un fox-terrier, [ p. 629 ] aux grands animaux du Pléistocène, mesurant environ 1,40 m au garrot. (2) Les molaires, initialement basses, cuspidées et munies de racines, sont progressivement passées d’une forme adaptée au broutage à une forme très longue, prismatique et complexe, adaptée au pâturage, et les mâchoires inférieures se sont développées en profondeur pour permettre cet allongement. (3) Le museau s’est allongé et les mâchoires se sont progressivement reculées. (4) Le cou, initialement court, s’est considérablement allongé et les vertèbres se sont modifiées afin de gagner en flexibilité sans perte de force. (5) Les membres s’allongèrent considérablement, les os de l’avant-bras et de la jambe fusionnèrent, ceux du côté interne (radius et tibia) s’élargissant pour supporter tout le poids, tandis que ceux du côté externe (cubitus et péroné) s’atrophièrent plus ou moins. (6) Les pieds s’allongèrent fortement et le troisième orteil, ou médian, de chacun s’élargissait progressivement jusqu’à supporter tout le poids, modifiant simultanément la forme du sabot pour qu’il devienne le seul support du corps. Les autres orteils s’atrophièrent progressivement et devinrent non fonctionnels, bien que souvent conservés sous forme d’attelles. Le premier orteil disparut en premier, puis le cinquième, puis le deuxième et le quatrième furent réduits à des ergots et enfin à des attelles. Ainsi, les chevaux pentadactyles de l’Éocène inférieur se transformèrent en espèces monodactyles du Pliocène et du Pléistocène.
Cerveau et mentalité. — Le cerveau du cheval est volumineux et complexe, ce qui suggère une grande intelligence, mais il n’égale pas celui de l’éléphant. La docilité du cheval et sa capacité d’apprentissage sont remarquables. Par ailleurs, il est sensible, et sa psychologie est étroitement liée à son mode de défense naturel — la fuite — puisque le premier réflexe d’un cheval domestique face à un phénomène inconnu est de s’enfuir. À l’état sauvage, ce même réflexe constitue un atout majeur pour sa survie. (Rires.)
Ancêtre des chevaux. — Lorsque le grand paléontologue anglais Richard Owen décrivit le petit « cheval de l’aube » européen à cinq orteils (Hyracotherium), une espèce proche du « cheval de l’aube » américain (Eohippus), il ignorait totalement qu’il s’agissait de l’ancêtre commun des chevaux, tant il était différent de la forme moderne. La découverte ultérieure en Amérique du Nord de stades de développement intermédiaires entre cet ancêtre et d’autres chevaux a clairement démontré que l’Hyracotherium était une forme généralisée issue d’une souche ayant donné naissance non seulement aux chevaux, mais aussi aux tapirs et aux rhinocéros. On constate ainsi comment une souche ancestrale se déployant dans différents environnements conduit finalement à des animaux d’apparence très différente, tout en [ p. 630 ] possédant des structures corporelles révélant leurs liens de parenté. « La conclusion est inévitable : le cheval, le rhinocéros et le tapir, trois espèces aujourd’hui très différentes, descendent par évolution progressive d’un ancêtre commun » (Matthew).
Succession géologique. — Le plus ancien membre connu de la famille des Équidés était l’« Eohippus », qui peuplait les forêts et les clairières de l’Éocène inférieur (Wasatch). C’étaient de gracieuses petites créatures, pas plus grandes qu’un chien, mesurant environ 28 cm au garrot. Leurs pattes antérieures possédaient quatre doigts fonctionnels et le vestige d’un cinquième, tandis que leurs pattes postérieures comptaient trois orteils et deux minuscules rémiges, vestiges du premier et du cinquième doigt. Ces éléments indiquent clairement qu’Eohippus descend d’ancêtres à pattes pentadactyles. Les molaires étaient courtes, pointues et de type brouteur ; le cou était très court, le corps long, avec un dos arqué, les membres et les pieds courts, et les membres postérieurs beaucoup plus longs que les antérieurs (Fig., p. 626).
Les chevaux de l’Oligocène présentaient un développement intermédiaire entre ceux de l’Éocène et du Miocène. À l’Oligocène inférieur (White River), les plus grands atteignaient la taille de moutons (Mesokippus). Leurs dents, initialement basses et de type brouteur, évoluaient vers celles des herbivores. Les pattes antérieures possédaient trois orteils fonctionnels, et bien que l’orteil médian fût le plus long, les deux orteils latéraux touchaient le sol. Les pattes postérieures étaient également à trois orteils, mais sans attelles (Fig., p. 627).
Au Miocène moyen, les chevaux à trois orteils étaient encore principalement des brouteurs, bien que certains des types les plus évolués se soient rapidement transformés en herbivores. C’est à cette époque qu’apparaissent les [ p. 631 ] formes intermédiaires (Merychippus) et que l’on observe la division de la famille des chevaux en plusieurs embranchements ayant évolué indépendamment. Le Pliocène inférieur présentait à la fois des types évolués et des types conservateurs de ces animaux. Au Pliocène moyen, parmi les troupeaux de chevaux parcourant les plaines, on trouvait trois embranchements de chevaux herbivores à trois orteils, mais les doigts latéraux ne touchaient plus le sol ; bien qu’il s’agisse d’ergots, ceux-ci conservaient parfois une certaine utilité. Au Pliocène supérieur apparurent les derniers chevaux brouteurs et herbivores à trois orteils, et c’est également à cette époque que vécut le premier membre à un seul doigt du genre Equus. Dans la première moitié du Pléistocène, on comptait au moins dix espèces du même genre, parmi lesquelles des chevaux plus grands que tous ceux vivant actuellement. (Voir figure, p. 630.)
[ p. 632 ]
¶ Les chameaux
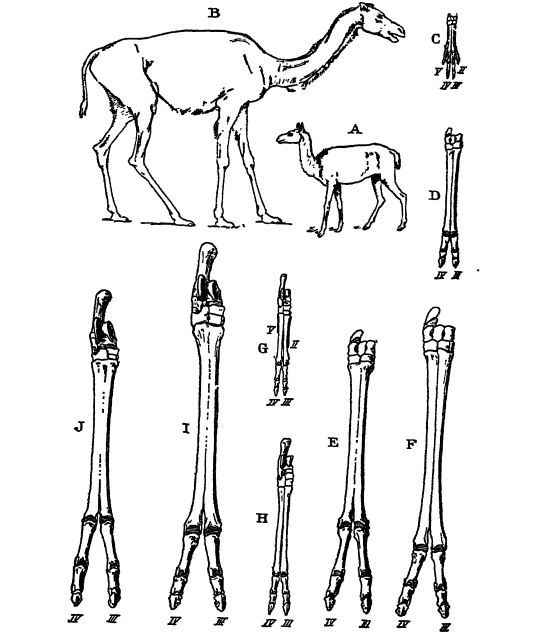
La famille des camélidés comprend aujourd’hui les deux espèces de chameaux des zones désertiques d’Asie centrale, ainsi que les ouamas et les guanacos des régions plus élevées et plus froides d’Amérique du Sud. Ces animaux étranges ont leur origine et leur évolution principale dans l’ouest de l’Amérique du Nord, et leur histoire est parallèle à celle des chevaux. Ils représentent des adaptations aux climats et aux plaines sablonneuses ouvertes. Les chameaux appartiennent à l’embranchement des ongulés, les Artiodactyles. Chez eux, l’axe du pied se situe entre le troisième et le quatrième orteil, et non, comme chez les chevaux, au niveau du troisième orteil.
Les premiers chameaux sont apparus à l’Éocène supérieur, sous une forme de la taille d’un chat domestique, et leur population s’est ensuite étendue tout au long du Cénozoïque. Au Miocène inférieur, les chameaux, à l’instar des chevaux, ont commencé à se diversifier. De grands chameaux étaient encore abondants en Amérique du Nord durant la première moitié du Pléistocène, avant de disparaître peu après. Les lamas se sont répandus vers le sud à travers les tropiques jusqu’en Amérique du Sud au Pliocène, tandis que, simultanément, les populations de chameaux se sont dispersées vers le nord, traversant finalement le pont terrestre Alaska-Sibérie pour gagner l’Asie, puis l’Afrique.
Scott affirme que le mode d’évolution des chameaux ne diffère en rien de manière significative de celui des chevaux. On observe la même augmentation de la taille et de la longueur relative des membres et des pieds, une diminution du nombre de doigts, passant de cinq à deux chez les chameaux (Fig., p. 631) et à un seul chez les chevaux (Fig., p. 627), ainsi qu’une évolution similaire des dents broyeuses à couronne haute à partir des dents de broutage à couronne basse. Les deux familles, chameaux et chevaux, sont apparues chacune au sein d’une même lignée.
¶ Les amblypodes à pieds courts
Parmi les mammifères archaïques de l’Éocène figurait un groupe d’herbivores, lents et à la mécanique squelettique maladroite, mais abondamment représentés par diverses espèces et genres. On les appelle Amblypodes, un nom qui fait référence à leurs pieds courts et trapus. Leurs pattes étaient robustes et massives, et leurs pieds courts possédaient cinq orteils munis de coussinets élastiques, comme chez les éléphants. À l’instar de tous les mammifères archaïques, leur cerveau était petit par rapport à la taille de leur corps.
Les amblypodes sont probablement originaires d’Amérique du Nord et datent de la fin du Mésozoïque, puisque les formes les plus anciennes connues remontent au Paléocène. Communs dans la région de Wasatch, ils ont atteint [ p. 633 ] une taille de plus en plus importante et se sont diversifiés au cours de l’Éocène supérieur, avant de disparaître dans la région de Bridger. Ces mammifères, très caractéristiques de l’Éocène nord-américain, sont remarquables, et seuls quelques individus (comme le Coryphodon) ont migré vers l’Europe occidentale.
Parmi les ambhypodes, les plus remarquables étaient les uintathères, ainsi nommés d’après le genre Uintatherium (voir fig. ci-dessous), découverts dans les monts Uinta, en Utah. Nombre d’entre eux atteignaient la taille de petits éléphants, mesurant jusqu’à 2,10 mètres au garrot. La forme générale de leurs larges pattes et de leurs pieds, mais pas celle de leur tête, rappelait celle des proboscidiens. Selon Scott, les uintathères étaient de véritables géants et les animaux les plus fantastiques de leur époque. Autrefois considérés comme des proboscidiens, il est aujourd’hui établi que les deux groupes ne sont aucunement apparentés. Ils étaient néanmoins les animaux ressemblant à des éléphants de l’Éocène nord-américain.
La tête des uintathères était volumineuse et composée de plusieurs os. Curieusement, le sommet du crâne était creux ou en forme de bassin, une caractéristique qui leur est propre. Ils possédaient de véritables cornes recouvertes de corne, comme les rhinocéros, une paire au-dessus du nez, et au-dessus des yeux et des oreilles se trouvaient d’autres paires de protubérances osseuses plus ou moins hautes, probablement recouvertes uniquement d’une épaisse peau. Autre caractéristique frappante, propre aux mâles : le développement des canines supérieures en deux redoutables défenses incurvées en forme de sabre. Afin d’éviter que ces défenses ne se cassent lors de leur utilisation, la mâchoire inférieure avait développé de larges replis osseux pour les protéger. Leur fonction demeure inconnue. Chez les deux sexes, les incisives supérieures étaient supprimées, comme chez de nombreux ruminants actuels. Le cerveau des uintathères était « ridiculement petit » et leur faible niveau mental parmi les mammifères est comparable à celui des dinosaures parmi les reptiles. Il s’agit peut-être de la principale cause de leur extinction, car ils devaient rivaliser avec les hordes toujours croissantes de mammifères modernisés, dotés d’un cerveau plus gros et donc plus alertes.
¶ Les bêtes géantes ou Titanothères
Au cours de l’Éocène inférieur, parmi les mammifères modernisés immigrants apparut un imgulé à doigts impairs (Eotitanops), plus petit qu’un mouton et d’apparence similaire à celle d’un tapir. Cette forme ancestrale, selon Osborn, [ p. 634 ] a évolué en onze branches principales, leur expansion débutant dans la formation de Bridger, se poursuivant dans la formation supérieure d’Uinta, et disparaissant à l’apogée de son développement au début de l’Oligocène (formation de White River). L’évolution la plus significative de ces animaux se déroule donc à moins de 60 mètres des strates oligocènes. À la fin de l’Éocène, ils se sont répandus en Mongolie (Protitanotherium), mais n’y ont apparemment pas vécu longtemps.
Ce sont les titanothères, ces « bêtes géantes », mammifères caractéristiques du Cénozoïque nord-américain. Seules les formes les plus récentes atteignent la taille de petits éléphants, mais le Brovtotherium, mis à part les éléphants, est « le produit le plus imposant de l’évolution des titanothères ». On ignore pourquoi les titanothères ont disparu alors qu’ils étaient à leur apogée, mais il est possible que l’aridification du climat au Miocène et la transformation de leurs habitats forestiers en prairies ouvertes aient déjà commencé et, le cas échéant, qu’elles soient à l’origine de leur extinction (voir Fig. ci-dessus).
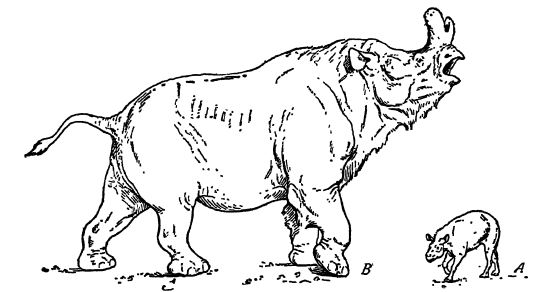
Les titanothères étaient des animaux massifs, aux pattes colonnaires et aux pieds courts, ces derniers reposant sur d’épais coussinets comme chez les éléphants. Chez tous, les pattes antérieures comptaient quatre orteils et les postérieures trois, et chez les formes plus anciennes et plus petites, les orteils et les sabots étaient plus saillants. Leur caractéristique la plus marquante réside dans l’évolution de leur tête. Chez les individus les plus anciens, le crâne était petit, long et étroit, dépourvu de protubérances. Les canines, en revanche, étaient proéminentes et ressemblaient [ p. 635 ] à des défenses, tandis que les incisives servaient à brouter la végétation. De toute évidence, les défenses des formes primitives étaient utilisées comme armes défensives. Dans la région d’Uinta (Scott, 1977), les titanothères étaient plus grands et présentaient de petites protubérances au-dessus des yeux qui, avec le temps, se sont progressivement développées et déplacées vers l’avant, jusqu’à ce que, dans la région de White River, ces cornes osseuses atteignent une taille considérable et soient situées sur le nez. Parallèlement à la croissance des cornes, le crâne se modifiait pour supporter leur poids et mieux résister aux chocs lors de leur utilisation. Lorsque les cornes devinrent des armes, les défenses devinrent insignifiantes et les incisives ne servaient plus à couper les dents, cette tâche étant désormais assurée par la langue et la lèvre supérieure. Au stade final du développement des titanothères, la tête était longue, très large, imposante et massive ; le profil, profondément concave, évoquait celui d’un rhinocéros fantastique doté de cornes nasales. Le corps et les membres de ces formes dérivées ressemblaient également à ceux de grands rhinocéros.
Le cerveau de la plus grande espèce était très petit, pas plus gros que le poing d’un homme, indiquant que ces grandes bêtes « devaient être incroyablement stupides et ennuyeuses, surpassant même les rhinocéros modernes à cet égard » (Scott).
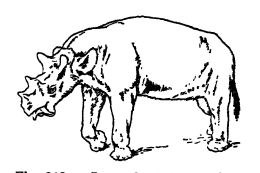
¶ Les rhinocéros
Les rhinocéros appartiennent à la famille des ongulés à doigts impairs ; ils possèdent généralement trois doigts et une peau épaisse. En règle générale, ils sont peu poilus, bien qu’une espèce laineuse ait existé à la même époque que l’homme au Pléistocène, dans le climat froid de Sibérie et d’Europe du Nord (Ceratotherium antiquitatis). Ce sont des brouteurs qui vivent dans les forêts, les steppes et les marais. Ils ont une grosse tête, un cou court et un corps massif et allongé, et leurs membres sont courts, robustes et colonnaires, comme ceux des éléphants. Les individus actuels mesurent de 1,20 m à 1,95 m au garrot. Les espèces à une seule corne se rencontrent en Inde et à Java, tandis que celles qui possèdent deux cornes en tandem, l’une sur le nez et l’autre sur le front, vivent en Afrique et à Sumatra. Aucun des fossiles américains n’était aussi grand ni aussi lourd que le plus grand spécimen actuel. Les cornes des rhinocéros sont particulières : contrairement à celles des bovins, elles ne sont ni creuses, ni osseuses, mais constituent des excroissances dermiques solides formées de poils agglutinés. C’est pourquoi on ne les retrouve jamais à l’état fossile. Leur présence chez les spécimens fossiles est cependant toujours indiquée par des os nasaux épaissis et rugueux.
Depuis le Pliocène moyen, aucun rhinocéros n’a été présent en Amérique du Nord. Pourtant, ce continent pourrait bien avoir été non seulement leur berceau, mais aussi le théâtre de leur évolution la plus significative. L’origine et le développement des formes ancestrales en Amérique du Nord, puis des rhinocéros géants de l’Ancien Monde, constituent une histoire très complexe, bien plus complexe que celle des chevaux et des titanothères. [ p. 636 ] Des formes ressemblant à des rhinocéros sont apparues en Amérique au début de l’Éocène, sous forme de petits animaux actifs et généralisés (Hydradiyvs, Fig., p. 638), qui, au cours du Cénozoïque, se sont diversifiées en au moins huit branches. Celles-ci sont regroupées en quatre lignées évolutives principales : (1) le groupe ancestral de petites formes de rhinocéros, sans défense, sans corne et coureuses ; (2) un type aquatique spécialisé, de grande taille et à courte durée de vie, vivant dans les rivières et les lacs ; (3) les gigantesques baluchithères d’Asie ; et (4) la souche principale des vrais rhinocéros, des animaux lents, de grande taille et généralement sédentaires. Au Miocène et au Pliocène, ces espèces étaient présentes sur tous les continents, à l’exception de l’Australie et de l’Amérique du Sud. Les populations ancestrales et aquatiques ont disparu d’Amérique du Nord durant l’Oligocène, et toutes au début du Pliocène.

Les formes sans cornes sont apparues en premier, puis, à différentes époques du Cénozoïque, des formes à cornes. Ces cornes peuvent être simples, doubles et disposées transversalement (espèces à cornes jumelées), ou encore l’une derrière l’autre le long de la ligne médiane du crâne (espèces à cornes en tandem) ; elles sont généralement placées au-dessus du nez, mais chez certaines espèces, la corne se situe sur le front. La présence de cornes, comme chez les titanothères, a entraîné des modifications de la forme du crâne (voir figures ci-dessus et p. 637).
Le plus grand mammifère terrestre découvert à ce jour est une forme de rhinocéros sans corne, connue sous le nom de Bdiichitherium. [ p. 637 ] Il vivait à la fin de l’Oligocène ou au début du Miocène et a été découvert pour la première fois au Baloutchistan, puis retrouvé au Turkestan et en Mongolie. Selon Osborn, il mesurait environ 4 mètres au garrot et environ 7,5 mètres de long ; en comparaison, tous les autres rhinocéros sont petits. Son cou était long et équin, sa tête longue et étroite, d’environ 1,5 mètre de long, et dotée de deux puissantes défenses. Ses membres, très longs et échassiers, lui permettaient de brouter le feuillage des arbres.
Scott distingue sept branches de rhinocéros véritables, dont trois sont encore représentées aujourd’hui. Il semble que ces animaux remarquables descendent du genre nord-américain Trigonias, vivant dans le cours inférieur de la White River. Cette forme sans cornes possédait quatre orteils aux pattes avant, contrairement aux trois autres rhinocéros. Un autre rhinocéros sans cornes de la même époque, Cenopus, tridactyle, a également existé jusqu’au Pliocène. Parmi les rhinocéros à habitat fixe, on remarque les petits dicératères (rhinocéros à deux cornes), apparus à l’Oligocène et ayant persisté jusqu’au Miocène (Fig., p. 636). Ils ont également migré vers l’Eurasie et se caractérisent par une paire de cornes transversales sur le nez. De l’Ancien Monde, le genre Teleoceras, une espèce aquatique dotée d’une petite corne sur le nez et de pattes anormalement courtes, arrivant en Amérique du Nord au Miocène moyen, s’est développé (Fig. ci-dessus). Cette espèce a persisté en grand nombre jusqu’au Pliocène inférieur.

Au cours du Miocène supérieur et du Pliocène inférieur de ce continent, au moins quatre espèces distinctes de rhinocéros [ p. 638 ] vivaient en très grand nombre, mais aucune n’atteignait la taille ni ne portait les grandes cornes des espèces actuelles. On ignore pourquoi elles n’ont pas survécu et pourquoi le rhinocéros laineux d’Eurasie n’a pas migré vers l’Amérique du Nord en même temps que le mammouth.
Les hyracodontes sont les ancêtres sans cornes de tous les rhinocéros. Le plus ancien spécimen connu date de l’Éocène (Bridger) et tous ont disparu à l’Oligocène (White River). Hyrackyus (Fig. ci-dessous), le genre le plus ancien, est très généralisé et pourrait avoir donné naissance aux chevaux et aux titanothères. Il était de la taille d’un mouton, mais plus lourd. Les hyracodontes étaient de constitution légère, avec une tête massive et maladroite, mais un cou et des membres longs et fins ; leur apparence évoque davantage celle des chevaux que celle des rhinocéros. De fait, les pieds des spécimens de White River ressemblent à ceux des chevaux de la même époque (Mesohippus). Leur sécurité dépendait de leur vitesse, d’où leur appellation de rhinocéros coureurs. Scott souligne qu’il est intéressant de constater que si les hyracodontes de White River et les tapirs ancestraux avaient vécu jusqu’à nos jours, ils auraient, au cours de leur évolution, développé des pieds à un seul doigt, comme les chevaux. Chez les hyracodontes, comme chez les chevaux, l’évolution s’est principalement traduite par un allongement du cou, des membres et des pieds, ainsi que par un élargissement des doigts médians. Avec l’augmentation de leur taille corporelle au cours de leur existence, leurs membres se sont renforcés, mais tous étaient de type coureur. Seul un genre a développé une paire de petites cornes nasales (ColonocercLs).
Les amynodontes étaient des rhinocéros aquatiques spécialisés, dépourvus de cornes. Ils sont probablement apparus sous le nom d’Hyrachyus à la fin de l’ère Bridger et ont disparu à l’Oligocène. L’Amynodon était plus petit et plus léger que le Metamynodon, plus tardif et plus imposant, de la rivière White. Selon Scott, ce dernier était [ p. 639 ] la créature la plus lourde et la plus massive de son époque. Sa tête était grande, aplatie et large, les narines étant situées haut, conformément à ses mœurs aquatiques ; son cou était court, son corps long et massif, et ses membres courts et robustes, avec quatre orteils aux pattes avant et trois aux pattes arrière.
¶ Lecture collatérale
R. S. Lull, L’évolution de la famille des chevaux. American Journal of Science, 4e série, vol. 23, 1907, pp. 161-182.
W. D. Matthew et S. H. Chubb, Évolution du cheval. American Museum of Natural History, Guide Leaflet Series, n° 36, 1913.
H. F. Osborn, L’Âge des mammifères. New York (Macmillan’), 1910.
H. F. Osborn, Le rhinocéros géant éteint Baluchithère d’Asie occidentale et centrale. Natural Historj", Vol. 23, 1923, pp. 209-22S.
W. B. Scott, Histoire des mammifères terrestres de l’hémisphère occidental. New York (Macmillan), 1913.
| XLII. L'évolution des mammifères et l'émergence de la pensée au Cénozoïque | Page de titre | XLIV. L'évolution des éléphants |