[ p. 207 ]
(See Pls. 4, 7, and 12)
History. — Trilobites were the fet fossils to attract the attention of naturalists and have long been of popular interest. It was Lhywd, the curator of the Ashmolean Museum at Oxford, England, who, in 1698, first called attention to an entire specimen. He did not know how to classify it but thought it was the skeleton of an unknown fish, and for a long time the animals were regarded as beetle winp, caterpillars, insects without wings, fo ssil butterflies, and even molluscs. The great Swedish naturalist, Linnaeus, first correctly recognized their relationship with the Crustacea, animals such as shrimps, crabs, and lobsters. Professor Waleh of Jena, Germany, in 1771, gave them the generic name Trilobites, a term that has since been raised to the rank of a subclass Trilobita of the class Crustacea of the phylum Arthropoda, which contains an exceedingly varied array of animals with articulate or segmented bodies and limbs, among them the crustaceans, insects, scorpions, and spiders. There are now more than two thousand kinds of trilobites known.
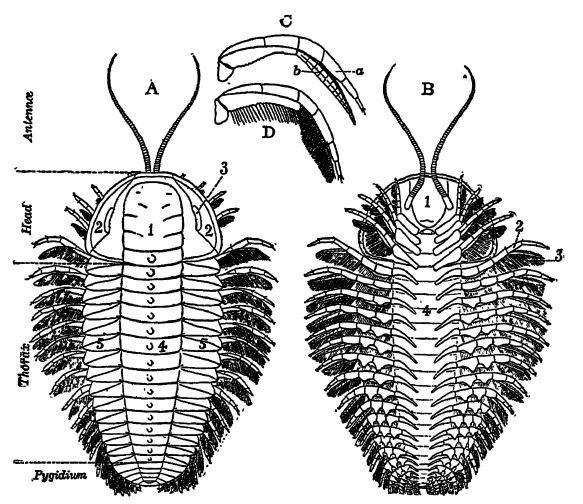
Definition. — The word trilobite means three-lobe-like, and has reference to the three longitudinal lobes (the central axis and lateral pleura) seen on the dorsal or upper side of most trilobites (see Fig., p. 208) . They were sexed animals. Their bodies were made up of segments, many of which articulated upon one another; and these segments were gathered together into three dhisions, as may be seen in any entire specimen of the upper shell or carapace, the part usually preserved. The imder, or ventral side, with the limbs, had a very thin outer shell and was preserved only in exceptional conditions, usually in black, more or less sulphurous shales. The carapace was thicker, and made up of chitin (very much as in horn, hair, etc.), a nitrogenous substance to which is added more or less of lime salts. Chitin greatly resists chemical alteration, and this is the reason why trilobites are so often preserved as fossils.
Habits and Habitat. — Trilobites inhabited only marine, and in the main, shallow waters, and their tests or outer shells are found [ p. 208 ] in all kinds of marine sediments, but chiefly in shales and limestones. In general they were rather sluggish animals, floating readily, but swimming probably in a jerking manner, and particularly backward, either with the ventral or dorsal side up. Over the sea bottoms they crawled slowly with the aid of numerous stout legs. Some of them dug into the sand for living food, going in head first, while others entered tail first and thus concealed awaited their prey, darting after it suddenly in the manner of fishes. Still others groveled around eating the mud. The small species of highly spinous forms may have spent their lives floating and swimming in the plankton, while those with large eyes may have dwelt in the dark deeper parts of the seas, rising at night to the surface in search of food. Many trilobites, and particularly those of the Cambrian [ p. 209 ] of the Atlantic province, are said to have been blind, having no e 3 'es, or certainly none on the dorsal side. Because of this blindness, they are thought to have lived generally in very deep or even abyssal waters, but the sediments in which they occur and the fossils associated with them indicate that they also lived in shallow seas.
Most trilobites could roll their bodies up like the sow-bugs or plll-bugs of our cellars, animals that are not insects but Crustacea (isopods) and distant relatives of the trilobites. This rolling up was for the protection of the softer and more delicate parts of the ventral side, thus presenting to the enemy the hard, thick carapace, an effectual armor against other trilobites but ineffective against the cephalopods and fishes. The power of coiling up first appeared in the Upper Cambrian, and with the Champlainian all forms had adopted such a method of protection.
As a rule, trilobites were camiyorpus, and as scavengers kept the sea bottoms cleaned of their dead animals; some were omnivorous; others probably wholly vegetarians; and a few were “mud eaters,” the digestive tract assimilating the organic matter in the muds for bodily sustenance.
In the early Cambrian, trilobites with large pygidia, or tail-pieces, began to be common, and this greater size, attaioed through the consolidation of a larger number of segments, is thought by some to have resulted from the use of the enlarged tail as a swimming organ by striking it rapidly downward and forward, causing the to dart backward as do the living crayfish and lobsters. The maximum of pygidial increase and greatest abundance of forms having it were attained in the Champlainian. This tendency toward an enlarging tail appears to have developed a little in advance of the first abundance of cephalopods, and as these animals probably fed on the trilobites, it is thought that the latter developed in defense not only a more rapid mode of swimming, but a backward one as well, a new locomotor addition to the abundant legs.
Size. — In size, the trilobites varied greatly at maturity, ranging in length from 0.38 of an inch up to 27.5 inches, but an average size was about 1.5 inches. Many species attained a length of from 3 to 4 and even 6 inches, but these were large individuals, and those above a foot in length were giants. Those represented as from 18 to 27.5 inches are, in nearly all cases, based on restorations pieced together from large fragments.
Geologic Duration. — Trilobites were characteristic of the Paleozoic era, beginning in considerable variety in the Lower Cambrian (about 37 genera and 125 special and dominating the seas of the [ p. 210 ] Cambrian (about 95 + 625) and the Champlainian (more than 120 + 1050). In the Silurian, though they were still common, the trilobites were nevertheless on the decline (35 + 500), and this ebbing of their vital force is seemingly shown in many picturesque forms replete with protuberances, spines, and exaggeration of parts. As a rule, in evolution, one finds that when an organic stock is losing its vitality there arises in it an exaggeration of parts, as if heroic efforts were being made to maintain the race. Spinosity in animals is often the prophecy of tribal death. In the Devonian, the variety and number of the trilobites were greatly reduced (30 + 120), at a time when the ancient t3q)es of fishes, which undoubtedly fed on these crustaceans, began to be common in the seas. In the later Paleozoic seas, the trilobites were relics, or animals surviving from a time better suited to their needs, and one by one they vanished, tmtil a little before the close of the Paleozoic era none were left. While they were in their prime, they dominated the life of the seas, since they were the chief carnivores and scavengers of Cambrian and Champlainian times, but with the rise of the cephalopods and the fiish-like animals in the Silurian and the true fishes in the Devonian their death knell was sounded by the incoming greater mentality and alertness of these higher types.
Definition of Parts (study with Fig., p. 208). — The carapace of trilobites is divided transversely into (1) a head portion, called the cephalon; (2) a series of segments, forming the thorax; and (3) a tail-piece or pygidium, forming the abdomen. The axis is the area of the vital organs, while the pleural regions are protective coverings over the ventral limbs.
The cephalon or head is the most important part of the animal, and in the classification of the group has the most reliable features for ordinal and generic distinction. It is composed of a medial glabella and outer free cheeks bearing the compound eyes. In many forms the free cheeks are separated from the glabella by a facial suture which generally shows as a faint raised line, and in all such forms the three parts are apt to separate after death and thus cause imperfect fossils. In some trilobites compound eyes are not known, such forms being called blind; usually, however, two more or less large compound eyes are present. In the majority of trilobites each compound eye is covered over by a single cornea and the individual lenses beneath may or may not show through. In others each lens has its own cornea. The number of lenses in a compound eye may vary from fourteen to the astonishing number of fifteen thousand. Imagine an animal with thirty thousand eyes!
[ p. 211 ]
The thorax consists of a variable number of articulating segments (in the most primitive of all trilobites, Naraoia, there are none), the number being constant in the adults of each species, but variable in the young of each form and in the genera. With one exception there are never less than two, as in Agnostus (Pl., p. 201, Fig. 4), or more than forty-four. In a general way, it may be said that where there are many thoracic segments the pygidium is small, and where there are few the abdomen is large. It should be stated here that so long as these animals are growing, the new thoracic segments added during the moulting stages are derived through the breaking down of the pygidium.
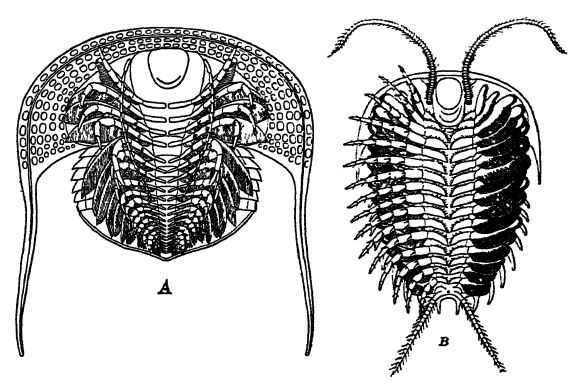
The pygidium or abdomen of trilobites may be small or large, and consists of a single piece of united segments varying between two and twenty-eight in number. Generally, the axial and pleural regions are plainly demarcated and some forms clearly show the number of coalesced segments.
Extending back from the pygidium in Neolenus are two cercopods that probably were used as sensory organs in the same way as the antennae (Fig. B, above). These cercopods may have had their origin in the crawling legs. While the cercopods are known only [ p. 212 ] in this genus, it may well be that other small-tailed trilobites had them, but in large-tailed forms they were lost when the pygidium became a swimming organ.
Ventral Side. — Beneath the glabella and hinged to its bent-over and thickened posterior margin is the hypostoma or upper lip covering the mouth (Fig. 5, p. 208). On each side of this plate there is a pair of short or long antennse, many-jointed sensory or feeling organs. All the limbs are in pairs, and each limb is double, or biramous, with each of the. two branches, in turn, made up of many joints.
The head has four pairs of such biramous limbs which serve also for chewing the food and passing it into the mouth. Back of the head each thoracic and abdominal segment has its similar pair of biramous limbs. Both limb branches arise from a single basal joint fastened to appendifers which are infoldings of the dorsal shell of the rachis. The stouter lower ramus or leg branch, with fewer joints, serves for crawling, while the other, which is more delicate and much more jointed, is used for swimming and is also replete with long hairlike appendages (setae) that are delicate tubes in which the circulating blood extracts from the water the necessary oxygen for life sustenance (Fig. D, p. 208). Lim bs are now known in twelve species and nine genera of trilobites, representing three orders and seven families.
Trilobite Moulting. — As trilobites grew, they periodically shed their chitinous shell, as do the crabs, and this happened many times before they attained full growth and probably occasionally during maturity. During the time of moulting, the animals grew and the detailed characteristics were changed, along with the introduction of new segments. These new segments were added just in front of the anal one, and at the time of moulting one or more of the forward segments of the pygidium may have been detached and added to the thorax. At first, the trilobite was a very minute, apparently unsegmented animal, all head (Pl., p. 242, Figs. 14, 15); then it consisted of head and pygidium, the latter of which rapidly added more segments; these stages were followed by other moults that long before maturity was attained rapidly introduced the full number of thoracic and pygidial segments according to the characteristics of each species. Later moultings simply enlarged the individuals.
Evolution. — Trilobites are the most primitive of Crustacea, and are generally assumed to have developed out of a many-segmented annelid in Proterozoic time. Recently, however, Raymond has postulated the progenitor of trilobites as having been a soft-bodied, depressed, floating-swimming, “worm”-like animal, composed of a few segments, probably with simple marginal eyes or even blind, with a mouth beneath the anterior margin, tactile organs at one or both ends, an oval shape, and a straight narrow gut running from the anterior mouth to the terminal anus.
It was probably the above type of animal that gave rise at some time during the Proterozoic to the trilobites, out of which then came some of the higher Crustacea. We may therefore say, with Raymond, that the trilobites gave rise, directly or indirectly, to all other arthropods.
For a complete discussion of the evolutionary trend among the Arthropoda the student is referred to Raymond’s book.
[ p. 213 ]
¶ Collateral Reading
P. E. Raymond, The Appendages, Anatomy, and Relationships of Trilo Memoirs of the Connecticut Academy of Arts and Sciences, Vol. 7, 19
C. D. Walcott, Appendages of Trilobites. Smithsonian Miscellaneous lections, Vol. 67, 1918, pp. 115-217.
C. D. Walcott, Notes on the Structure of Neolenus. Ibid., Vol. 67, pp. 365-457.