[ p. 289 ]
In this chapter we are to study the fishes, their kinds, their ancestry, and how their more versatile descendants came to leave their ancestral water habitats for the land. It is a most fascinating study, since all humanity is interested either in the great economic value of fishes, or in the happy hours their pursuit gives the angler, but it has a special interest for the philosopher who sees in the fishes a step on the path of evolution that leads from the wormlike invertebrates to the pinnacle of organic independence in reasoning man.
Fishes were the first markedly successful class of vertebrates. With the possession of a vertebral column and a central nervous system, and with their greater alertness, mobility, and fecundity, they have an easy preeminence over their invertebrate inferiors. They are as markedly adapted to the water as the birds are to the air. As a rule they live either wholly in the seas and oceans or wholly in the fresh waters of the lands. Many of the marine fishes, however, spend part of their lives in fresh waters; some feed and breed in either fresh or marine habitats; others feed in the sea and breed in the rivers, or in rare cases, like the eels, pass from the rivers out into the sea to breed. Of the marine fishes ascending rivers to breed, some have remained there through choice, others have become “ land-locked through the rising of land barriers, and have evolved [ p. 290 ] into distinct races or species. Through stream capture and the spreading of fish eggs on the muddied feet of wading birds, the fresh-water fishes have attained their present wide radiation over the continents.
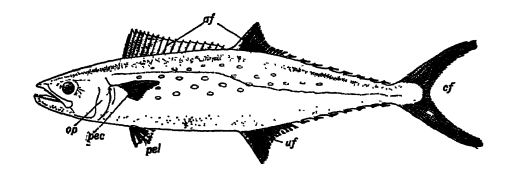
Distinctive Features. — Fishes are vertebrates without a neck. They also have no ears and therefore there is no ear drum present. Their body temperature is about that of the medium in which they live, wherefore they are said to be cold-blooded. Because of their water habitat, all fishes have gills and fins (see Fig., p. 289). The gills are on each side of the head and consist of delicate, hollow filaments in which the blood circulates. Water is taken in through the mouth and passed out over these breathing organs, where the blood in the tubes extracts the free oxygen dissolved in it. The mouth is usually provided with teeth. An air bladder is generally present and then may serve as a hydrostatic organ or float, though in certain forms it is modified into a cellular sac, which acts as a lung and assists the gQls in respiration.
The skin is either soft and naked, or, more commonly, protected by the development of spines, mosaics of denticles, overlapping scales. or bony plates known as scutes. The principal organ of locomotion is the powerful tail. This is assisted by the paired fins, of which the forward set, known as the pectoral limbs, lie just behind the gflls, and the rear pair, or pelvic limbs, are placed fartW back (see Fig., p. 289). The paired fins of fishes correspond to the fore and hind limbs of the higher vertebrates. Their skeleton, however, cannot be readily compared with the limb structure of other vertebrates. How these fins became limbs is discussed later in this chapter. Pishes also usually possess a variable number of unpaired median fins that are used mainly as balancing organs; those on the back are called dorsed fins, those on the under side between the anus and tail are known as anal fins, and the tail represents a caudal fin (see Fig., p. 289). AH the fins are supported by skeletal bars or rays, called fin rays.
The skeleton throughout may be of cartilage, though usually it is more or less bony. The main skeleton consists of the vertebral column. The bones of the head are of two kinds, dermal or skin Ixmes like the scales, and the actual bones of the brain case.
Types Fish Tails. — The tail presents three general types. (1) The vertebral column continues in constantly diminishing joints nearly to the end of the a nimal , and is there surrounded by a S3rmnretrical, one-lobed, vertical tafl fin. This is the most primitive type, very ccwninon among the Devonian fishes, and seen in the baby stages of most living fishes; it is known as the diphy cereal or symmetrical [ p. 291 ] [ p. 292 ] tail (see Pl., p. 295, Fig. 4). (2) In others, the vertebral column bends upward and terminates with diminishing joints in the upper and larger lobe. Hence this type of tail is rarely symmetric, and is called heterocercal because of the inequality of the lobes (see Pl., p. 291, Figs. 2, 4; Pl., p. 295, Figs. 1, 2). It is common in living sharks and in most Paleozoic fishes. (3) The modern bony fishes also have a symmetric tail, but while the vertebral column here again bends upward, it ends rather abruptly, and from the terminal joints the fin rays develop in such a way as to form a symmetric tail. This kind of caudal appendage is called homocereal (see Fig., below).
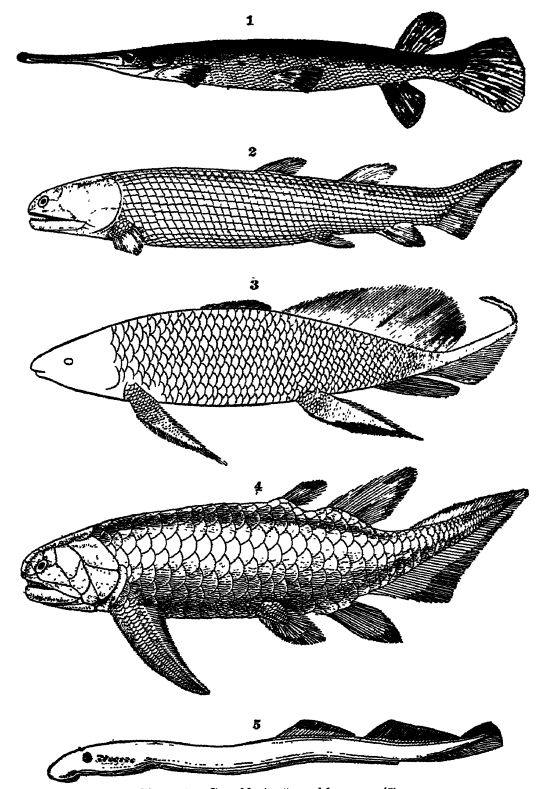
Fig. 1, living American gar-pike (Lepidosietis osseiLs), x 2, Devonian ganoid from Scotland (Osteolepis macrolepidotu8)i x J; 3, Upper Devonian ganoid from Scaumenac Bay, Canada (Scaumenacia curia), x 4, Devonian ganoid with Q’cloid scales, from Scotiand (ffolopiychius flemingi), X §; 5, Innng marine lamprey (Pefromyzon marinus), x i, note the seven openings back of the eye, exits from the gills; also edge of circular mouth, and absence of paired fins.
Figs. 2 and 4 after Woodward, British Museum Guide Book; Fig. 3 after Hussakof.
Food of Fishes. — Among living fishes some feed exclusively on plants, some on plants and animals alike, some exclusively on animals, and others on the mud m which minute plants and animals occur. The majority of fishes, however, feed on other fishes, without much regard to species, devouring their own young as readily as those of any other kind. Teeth with blunt tips usually serve for crushing shells; teeth with serrated edges often signify plant-feeding forms; while strong incisors may indicate the choice of fina-ils and crustaceans as food. In general, the sharper the teeth and the larger the mouth, the more distinctly a species is a fish-eater. Small-mouthed fishes are apt to be plant- or mud-eaters.
Growth. — Fishes grow as long as they live and their size depends upon their ancestry plus the amount of food procurable: the length of life is variable; some exist many years, others are short-lived, and some appear to be annuals .
Ancestry of Fishes. — In spite of the facts that most fishes have bony skeletons and that fish-like animals are known as far back as the Middle Champlainian, their more primitive fossil ancestors are still undiscovered by paleontologists. Nor are the zoologists able to determine their ancestry among the vast horde of living forms and their embryos. All that the paleontologist can say is that the oldest fosal vertebrates are undoubted fishes, and seemingly of elasmobranch and ostracoderm kinds. Under these circumstances, [ p. 293 ] we will leave this matter to future discoveries and go at once to a description of the kinds of fishes.
Classification of Fishes. — For present purixises, fishes may be divided as follows:
- Class Pisces or true fishes.
- Subclass Elasmobranchii or gristle fishes
- Order Acanthodei or spinous sharks
- Order Selachii or true sharks and rays
- Subclass Ostracodermi or aberrant extinct shark
- Subclass Ganoidei or enamel-sealed fishes
- Subclass Teleostei or bony fishes
- Subclass Dipnoi or true lung-fishes
- Subclass Arthrodira or armored fishes
- Subclass Elasmobranchii or gristle fishes
¶ Subclass Elasmobranchii or Gristle Fishes
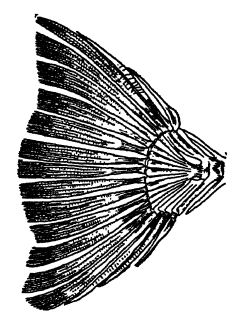
The word Elasmobranchii means having plate-like gills, and the group includes the ancient and modern sharks, sawfishes, sea-cats (Chimaera), skates, and rays. With few exceptions, the living forms are inhabitants of marine waters (see Pl., p. 295, Fig. 3). They are the most primitive fishes, and very early in the Paleozoic give rise to the higher types.
In most elasmobranchs the mouth is on the under side of the head, but in some it is at the forward end. The internal skeleton is of gristle or is cartilaginous, with more or less calcification. The tail is usually heterocercal. The skin often appears to be naked, but there are always dermal denticles (small pieces) tipped with hard, shining enamel, cored with dentine (phosphate and carbonate of lime with fluoride), and with a bony (phosphate of lime) base sunk in the deeper part of the skin. This type of scale is called placoid, and has the same structure as teeth, which originally also developed out of the skin. When the denticles are small and closely set they constitute the so-called shagreen (meaning a rough mosaic).
The five gill slits on either side of the body back of the head are, in living cartilaginous fishes, either open or covered by a skin flap, never by a bony operculum as in most other kinds. The median fins are often provided with long, sharp, homy or bony spines, this being particularly true of the dorsal ones. It is these spines and the teeth of sharks that are usually preserved as fossils; unfortunately, however, they give little information as to the general characteristics of the animals.
The brain is enclosed in a solid cartilaginous box or skull without sutures or joints, and in this feature the elasmobranchs differ from [ p. 294 ] nearly all other fishes (the exception being rarely among ganoids), in which the separate skull bones are easily distinguished. There is no swimming bladder.
The acanthodian or spinous sharks are small and the most primitive of true jfishes. They appear in the late Silurian, are particularly common in the Old Red sandstone of Scotland, and rarely exceed a foot in length (see Pl., p. 295, Fig. 1). They are differentiated from the true and most other Paleozoic sharks by their plated skin, in which are embedded small, flattened, closely fitting, bony plates composed of dentine and enamel, and by the further fact that the stout spines are not restricted to the median fins but are also developed in connection with the paired fins. The teeth are small and rather of the pointed type like those of modern sharks. The acanthodian sharks are either directly or indirectly the progenitors of the later sharks and the higher ganoids.
The shell-eating sharks were especially well represented in the later Paleozoic, by two families known as cestracionts and cochliodonts. The first group is now best represented by the Port Jackson shark, which lives off South Australia (see Pl., p. 295, Fig. 3), and is characterized by having two stout dorsal fin spines and, in the deeper parts of the mouth, many blunt and pavement-like teeth adapted for crushing shellfish and crustaceans (therefore also known as the “ oyster crusher ”). In the cochliodont sharks the individual teeth are fused into two large plates whose upper surface has spirally curved ridges suggesting the spirals of snail shells, the characteristic that gave rise to the family term (see Fig. A , p. 342) . It is these hard parts that are so often seen in the later Paleozoic and far less commonly in the Mesozoic; the cochliodonts, however, vanished with the Carboniferous.
The modern sharks are the true Selachii (a Greek term meaning sharks). The order includes the large, active, more or less spindle-shaped sharks, the sawfishes, and the flattened, sluggish, bottom-living rays and skates of the present time and of the Cenozoic and Mesozoic eras. This type of shark with sharpcutting teeth has been abundant. In the Miocene of South Carolina occiir teeth of this type, the largest of which are nearly 6 inches in length (Carcharodon), indicating sharks with a length of 60 feet. The living great blue shark attains 40 feet in length.
Geologic Occurrence. — In America the oldest shark remains are small fin spines, apparently of acanthodian kinds (Pl., p. 295, Fig. 1), found in the late Silurian (Cayugan and not Clinton as originally stated) of Pennsylvania. With the Middle Devonian, spines, teeth, and skin denticles of small and large sharks become common. In the Mississippian the shell-feeding forms were highly diversified (see p. 341), but lost their ascendency in the Pennsylvanian and practically vanished with the Paleozoic. The cutting type of tooth so general in living sharks is rare in the Paleozoic, is seen more often in the early Mesozoic, but is not common until Cretaceous time.
¶ SubcLass Ostracodermi or Aberrant Sharks
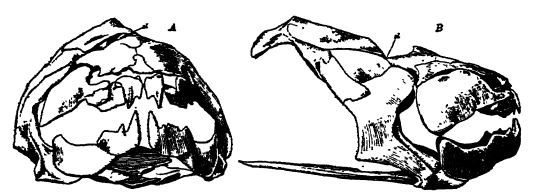
The Ostracodermi are heavily armored, and possibly a group of highly aberrant cartilaginous sharks. The subclass includes the oldest known fishes, beginning in the Middle Champlamian and vanishmg in the late Devonian. They represent a line of fish evolution [ p. 296 ] in the wrong direction, for they have left no descendants. They are. in fact, so different from other true fishes as to make impossible as yet to indicate their relationship to other vertebrates. The osiracoderms were small, none longer than 7 inches, probably bottom-feeders and of sluggish habits, apparently living mainly in the lagoon areas of the .seas and in the rivers. The head and anterior body was large, broad, and depressed, and armored by small placoid plates or scales, which were sometimes fused into more or less thick bony plates, while the posterior region was either naked or more or less covered by similar plates or scales. The tail was heterocercal, and there was no internal skeleton (see Figs., above, and p. 297).

Fig. 1, Lower Devonian acanthodian shark, from Scotland (Climatius X i: 2. Upper Devonian shark, from Ohio (Cladoselache fyleri), x J; 3, livin Port Jackson cestraciont shark, from Austraha (Cestracion philippi) x 1/10; livin lung-fish of Australia (Neocerotodus fosters) X A; 5, Devonian lung-fish,

In structure and form, there was considerable variety among the bony-skinned fishes. The older kinds, of very diverse construction, were without a pair of lateral paddles or swimming limbs (Pteraspis, CephalaspiS, Drepanaspis), and the eyes were either far apart or set closely together (see Fig., p. 297). Restricted to the Upper Devonian occur the forms with anterior armored paddles, decided armor over the anterior region, and eyes closely set (Bothriolepis, Pterichthys, once regarded as gigantic beetles, see Fig., above). It is possible, however, [ p. 297 ] that the latter genera — the “ winged fishes ” of Hugh Miller, who remarked that his first impression was that he had found in them “ the connecting hnk between the tortoises and the fish ” — do not belong in this subclass. The oetracoderms are widely distributed throughout Europe and North America.
¶ Subclass Ganoidei or Enamel-scaled Fishes
The term Ganoidei, which means bright appearance, has reference to the glossy surface of most of these fishes, as, for instance, in the living gar-pikes (see Pl., p. 291, Fig. 1). This glossiness is due to the more or less thick rhombic scales found in gar-pikes or the cycloid scales in Devonian forms, which are covered with bright enamel of the same character as the outer covering in teeth. The ganoid scales differ from the placoid ones of elasmobranchs in that they have but two layers, an outer enamel and a thicker bony layer beneath. In. some ganoids, however, there are true elastic cycloid scales, the kind so conomon in the food fishes, i. e., thin circular plates of phosphate of lime, but without enamel (see Pl., p. 291, Fig. 4). The ganoids are essentially fresh-water fishes, although some are marine and only ascend the rivers to spawn. In the geologic past [ p. 298 ] the marine members seem to have been considerably more common than now. The best known living examples aie the sturgeons and gar-pikes, but the origin of the subclass goes back at least to the Lower Devonian, for they constitute one half the known fish fauna in the Old Red sandstone fsee Pl., p. 291, Figs. 2-4) and continue to be abundant until early Cretaceous time. In their organization they form a connecting link between the acanthodian sharks and the bony fishes.

The skull is well protected hy dermal bones or is completely ossified, and while the interior skeleton is more or less bony, there are some forms in which it is wholly cartilaginous. In this subclass appears for the first time the perfected fish mouth, in which the lower jaw operates against the teeth of the perfected upper jaw, as in the higher vertebrates. An air bladder is always present and is connected with the gullet; in the gar-pikes it assists in respiration but is never cellular as in the lung-fishes. There is a spiral valve in the intestine, and the tail is heterocercal or diphycercal.
Fringe-filmed Ganoids. — According to the character of the fins, the ganoids are divided into two orders, namely (1) the fringe-finned or lobe-finned ganoids (Crossopterygii), and (2) the pillar-finned ganoids (Stylopterygii). In the latter kind the limbs have not the thick, muscular, scale-covered lobes of the other order, but are more like those of the food fishes. Of the first order there are now but two genera, both of which live in the fresh waters of Africa. Even though their air bladder also functions as a lung, these fishes cannot live out of water more than three hours. They were the common fishes in the Lower Devonian, the best known forms beiug the genera Holoptychius, Eusthenopteron, and Osieolepis (Pl., p. 291, Figs. 2-4). In these ganoids the layers of bone of the conical teeth are often deeply and complexly folded or labyrinthine, and in this are particularly interesting because the same kind of teeth occur in all of the Paleozoic amphibians. This similarity of tooth structure is another hint showing the probable origin of the Amphibia (order Stegocephalia) in these fiinge-finned Devonian fresh-water ganoids.
¶ Subclass Dipnoi or True Lung-fishes
The Dipnoi (from the Greek words me aning double breathing) are the “lung-fishes,” so termed because the three living genera have an air bladder which opens into the mouth and which is either single or double, vith numerous cellular spaces. This is an outgrowth of the anterior region of the digestive tract and serves as a lung, supplementing, and in times of drought supplanting, the functions of the gills as organs of respiration. From an evolutionary standpoint, the lung-fishes are very important, because the air bladder is comparable with the lung of the higher vertebrates [ p. 299 ] in that it returns the aerated blood direct to the heart, whereas in most other fishes the blood is carried from the air bladder through the general circulation before reaching the heart. The nasal sacs are on the exterior of the snout and open into the mouth, a condition met with also in Amphibia and higher vertebrates, but very rarely in fishes. It is in the lung-fishes, then, that we see the possibilities for the development of the higher vertebrates, though this does not necessarily mean that they were the progenitors of the latter. In many ways, they are intermediate in structure between the lower fishes and the amphibious salamanders, but they are not thought to be the direct connecting links between the fishes and the amphibians. It is probable that the fringe-finned ganoids (crossopterygians) were the progenitors of the amphibians.
From the living lung-fishes we learn that certain forms (Lepidosiren) inhabit muddy streams and marshes, where they feed on plants. Others feed on worms, insects, shellfish, crustaceans, and frogs. They are sluggish animals, rising occasionally to the surface for a new supply of air; in times of drought, Lepidosiren encases itself in a cocoon made of slime and clay, and breathes air through an opening made in the mud. During the rainy season, living lungfishes eat voraciously, storing up great quantities of fat between the muscles, and this fat sustains them during the dry’ season, so that they can live without food, sometimes for eight months, until the water returns.
In the living forms there are also either two or four pairs of gills covered by a movable bony operculum. They are generally eel-like in appearance, with the dorsal and ventral median fins usually continuous with the tail fin. Among the Devonian forms the tail is usually of the heterocercal type (Pl., p. 291, Fig. 2), but in recent forms it is diphycercal (Pl., p. 295, Fig. 4). The paired fins are long, broadly lobed, muscular, scaled, and fringed in the Dev’enian species and in living Ceratodus, but in other modern forms they are very slender, pointed, and without the fringe. The body is covered with overlapping cycloid scales.
Dipnoans are cartilaginous fishes in which there is, however, some ossification, but they have no vertebra differentiated as such. The skull consists of cartilage covered by a variable number of membrane bones that are thickest in Paleozoic genera. These are superficial bones developed in the membrane covering the cartilage, and are not formed in the cartilage itself. The teeth are few in number, usually in three pairs, of which two sets are large crushing plates.
[ p. 300 ]
The dipnoans are closely related to the fringe-finned ganoids, these two types of fishes being the most important offshoots of the primitive sharks and evohdng toward the higher vertebrates. The oldest dipnoans, in about fifteen species, occur in the Devonian of both Europe and America.
¶ Subclass Arthrodira or Armored Fishes
The most striking fishes of the Devonian seas were the highly armored Arthrodira, almost 40 per cent of all Devonian fishes being of this subclass. The term Arthrodira means jointed neck, and is given because the armor of the head slides somewhat over that of the body, thus allowing the head to move up and down, a rare feature among fishes and best developed in this subclass. Arthrodira are more or less heavily armored and in this respect remind us of the lung-fishes and the earliest amphibians. Their systematic relationship is still unknown, however, but apparently they are nearest the lung-fishes and the fringe-finned ganoids. It is because of this supposed relationship that they are called by A. Smith Woodward large-headed lung-fishes. Of course, nothing is known about the lungs and breathing habits of the Arthrodira.
The oldest arthrodires occur in the early Devonian fresh-water deposits of Germany, while those of the Middle and Upper Devonian of North America were marine. They were the largest and fiercest fishes of this period, but died out in the earliest Mississippian, a time when the shell-feeding sharks rose to the dominancy of the seas.
The arthrodires had no true teeth; the structures functioning as such were large cutting shears or crushing plates. The tail was heterocercai and there appears to have been a pair of pelvic fing. [ p. 301 ] Of an internal skeleton there was little, for the vertebral axis was not bony, though the neural and ventral arches were somewhat ossified.
The Arthrodira easily dominated the aquatic life of the Devonian seas, one American form, Dinickthys, the “terrible fish,” attaining a length of over 20 feet (Fig., p. 300). There are more than forty species known in North America, and one locality in Germany (Wildungen) has alone jdelded upward of fifty forms.
¶ Subclass Teleostei or Bony Fishes
The teleosts (means true bone) are the modern fishes and express the highest organization among these animals, for their evolution has tended to complete adaptation to the water (see Fig., p. 289). They constitute about 99 per cent of all living fishes. Most of the fishes of commerce and sport are of this subclass and occur abundantly in the fresh waters, seas, and oceans. Their origin was in the Ganoidei, apparently in late Paleozoic time, but they were not common imtil the Lower Cretaceous, with their best development in the Cenozoic and at present.
The bony fishes are nearly always covered with thin, elastic, cycloid scales, never with placoid, and very rarely with ganoid scales. In some the skin is naked and slimy. The caudal fin is usually homocercal and the paired limbs are never lobed. The gills are always covered by a movable operculum. The elongated air or swimming bladder, when present, is filled with gas and lies dorsally to the body ca-v-ity; it is not a lung but a hydrostatic organ. The internal skeleton is wholly ossified, the vertebrae are hollow at both ends (amphicoelous), there is a bony neural arch enclosing the spinal nerve-cord, and each vertebra has also a pair of ribs enclosing the viscera. Many of these characters are also seen, but less perfectly, in the Ganoidei, and therefore some naturalists regard both subclasses as representing but a single group.
¶ Origin of Fins and Limbs
Development of Fins. — In the higher marine animals, purposeful locomotion begins through lateral imdulating of a flexible body, to which were added fins to assist in progression through the water. Fins are of two kinds, the unpaired fin-folds and the paired fins. The originally flexible unpaired fln-folds arose out of the skin and later became rigid, giving the fish a more resistant surface. Such may be more or less continuous from the head along the mid-line of the back (dorsal fin), around the tail (caudal fin), and forward [ p. 302 ] along the under side as far as the vent (anal fin). It is usual in most fishes to see these unpaired fins discontinuous, that is, localized.
The paired fins, like the unpaired ones, in all probability originated. Lull says, in skin folds, growing out from the two sides of the body, and at firet served as keels that gave better balance to the narrow fish body, or checked its forward motion. They are well shown in the acanthodian sharks of the Silurian and Devonian (Pl., p. 295, Fig. 1), and here they were held e.vtended by stout bony spines, just as one sees the dorsal unpaired ones held vertically in modern sharks. In other forms, these lateral bony spines were replaced by internal stiffening rods. Osborn says that in the course of evolution these rods are concentrated to form the central axis of a fully jointed fin, and with further development they transform into the cartilages and bones of the limb girdles (see Fig., above).
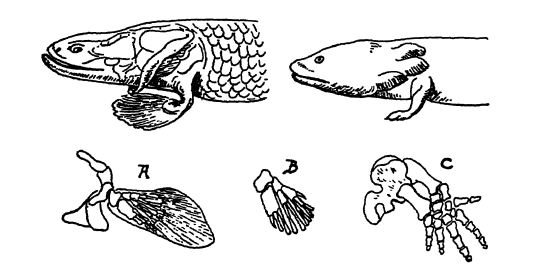
In the fishes, the paired fins with their girdles are the rudiments out of which legs and feet were developed through the enforced hobbling about of the fringe-finned ganoids in their search for water holes in desert regions. In them it is thought was developed a stracture prophetic of legs and feet, though the actxial transitional limb, either in fossil form or in embryos of living limbed vertebrates, is wholly unknown.
The oldest known footprint, Thinopus (Fig., p. 331), from the Upper Devonian, may give a hint of what this evolution was like. There appear to be here but two completely formed fingers, probably the first and second, separated [ p. 303 ] by a deft that extended deeply into the sole of the foot; the fossil indicates further that a nidimentary third toe was present and possibly even a fourth one. If the structure of this footprint can be relied upon, then the terrestrial foot began with two toes instead of five, as is usually held.
¶ Origin of Double Breathing in Fishes
Breathing Organs of Marine Animals. — A survey of tj-pical marine animals reveals many and diverse methods for the extraction of the free oxj-gen from the water, but whatever the kind of breathing organ, it is always of the same fundamental tj-pe, a localized organ or organs having delicate canals with very thin membranes in which the blood circulates, bringing the red corpuscles into almost direct contact with the surrounding water. The hsemoglobin of these red corpuscles has decided aflSnity for oxygen, which it extracts from the water and which the floating cells carry through the circulation of the blood to all parts of the animal’s body. Nowhere in the sea has any animal an additional organ for the extraction of oxygen directly from the air, excepting in such stocks as are known to have had land-living ancestors.
Evolution of Lungs. — There is in most fishes above the sharks an organ known as the air or swimming bladder, which is structurally a sac-like outgrowth, single or paired, of the alimentary canal. The principal function of this degenerate and secondarily modified structure in most living fishes is that of a hydrostatic organ, to give buoyancy to the animals, and it is controlled by muscles in such a way as to permit its possessor to remain at any desired level in the water. The air bladder is the homologue of the lungs of the terrestrial vertebrates, and is utilized by the so-called lung-fishes, and to a limited extent by other relic fishes (Amia, Lepidostevs, Pdypterus) as a respiratory organ supplementary to the gills.
The great range of modification of this structure shows it to be of extremely ancient origin, and its incipient condition is possibly shown by a pair of pouch-like outgrowths of the pharynx, or throat cavity, in the sharks. Stagnation of the water and a loss of free ojygen. would bring the fishes to the surface to gulp down air, and such pouches, if supplied with blood-vessels, would serve in a very rudimentary way to aid in aerating the blood. A premium placed upon such structures would, it is thought, stimulate their development to the condition seen in the modern lung-fishes.
Origin of Lungs in Anitnabs Living in Temporary Bodies of Water. — Animals originating and living in permanent bodies of [ p. 304 ] water, and especially in the ocean, have no need to breathe the air, and it is therefore held that the stimulus for such an alteration could have arisen onh- where the water periodically failed the animals. No fish permanently breathing air is known to inhabit the region between low and high tides, though several kinds live here for a time in search of food. On the other hand, in none of the marine deltas of the present does one find the climatic conditions necessarj- to force fishes to develop into lung-fishes, nor do they occur in the areas of the strand-lines or in the courses of the permanent rivers. Xowhere is there in these places a transition zone forcing the waterliving fishes to adapt themselves to the dry land.
Just as we have arid regions to-day, so it appears that si mila r land climates existed during much of geologic time. Under such climates, bodies of water come and go according to the season of rain and drought, and hence various methods are resorted to by the aniTnals to maintain their kind or themselves over the period of drought. During the arid season the struggle for existence is severe, not only because of the abnormal crowding of the individuals into constantly dimmishing spaces and the reduction in the amount of av’allable food, but even more so because of the increasingly saline and bitter character of the water. It is thought that under the stimulus of these changes gill-breathing fishes first adapted themselves to burrowing in the sand. Thus protected in water and mud holes there was for a time moisture to pass over the gills, but under such environments life was very precarious and in the struggle most of the individuals were destroyed. After innumerable failures in their efforts to gulp the air into the pharynx, efforts lasting through long geologic time, the ganoids and lungfishes were gradually developed and perfected, their first appearance being in earliest Devonian time.
Highest Mentality among Land-living Vertebrates. — Air breathing, once established among the fishes, has resulted in the land vertebrates attaining the highest mechanical and mental perfection, an evolution necessitated by adaptation to a wide range of environmental conditions as compared with the relative uniformity of the sea. The greatest mentality in the sea has been repeatedly derived from the continents, in that many stocks of land v’ertebrates have adapted themselves to the sea because of the ease with which they can there prey upon the less alert and intelligent. Out of such stocks, however, comes no higher mentality. They represent an adaptation in the wrong direction, that is, to an easier life, for the hipest mentality has been developed only on the land where [ p. 305 ] the strafe for existence is greatest because of the constant necessity of adaptation to changing environnient. Organic supremacy is attained only through constant vigilance.
¶ Collateral Reading
Joseph Barrell, Influence of Silurian-Devonian Climates on the Ri«e of Air breathing Vertebrates. Bulletin of the Geological Society of America, Vol. 17, 1916. pp. 387-436.
T. C. Chamberlin, On the Habitat of the Early Vertebrates, Journal of Gcolo©% Vol. 8, 1900, pp. 40(M12.
Bashford Dean, Fishes, Living and Fossil. New York and London (Macmillan), 1895.
C. R. Eastman, Devonic Fishes of the New Yoric Formations. New York State Museum, Memoir 10, 1907.
R. S. Lull, Organic Evolution. New York fMacmillan), 1917.
H. F. Osborn, The Origin and Evolution of Life, New York (Scribner), 1917.