| XXVII. The Pennsylvanian Period, the Time of Greatest Coal Making | Title page | XXIX. Coal and its Occurrence in Nature |
[ p. 373 ]
Plants of some kind occur in all lands and under all climates. They are not restricted to the moist soils of the valleys, but live also on the highest mountains and even sparingly in the driest deserts. In the shallow fresh waters are found many kinds of plants, and on the ocean bottoms down to a depth of about 600 feet grow the attached seaweeds. Fastened to rocks, trees, and other objects are the brown and gray lichens, a thin film of spreading plant matter. In the moist forests and on the meadows occur the fimgi, including the common toadstools, plants devoid of chlorophyl and feeding on the humic material (vegetable mold) of other dead and dying vegetation. A vast variety of microscopic plants, the bacteria, live everywhere — in the soil, in nearly all waters, in the oceans, on and in other plants and animals. They and the fungi are Nature’s main agencies for dissolving dead organisms, consuming oxygen and giving off carbon dioxide. The yeast bacteria convert sugar into alcohol by fermentation, and are used in the making of alcoholic liquors; other bacteria are useful in the making of cheese; still. others cause diseases, and are found even in all healthy organisms.
Paleozoic Floras. — From the geologic record we learn that the first known flora came into existence in the latter half of Devonian time, though a few land plants are known from the Silurian. We are probably right, however, in considering that the low-lying lands were clothed with a green verdure from at least the beginning of Champlainian time, but that the early plants had not yet acquired sufficient woody tissue in their organization to be preserved in the sediments, and hence have no significance in Historical Geology. Since Middle Devonian time land plants with woody tissue have become more and more prolific and have grown to greater dimensions and higher stature, all of which means that they also have gradually attained a longer individual growth.
Swamp Life of the Pennsylvanian. — The coal floras of late Paleozoic time are now well known, and it is estimated that upwards of three thousand species have been described (see Figs., pp. 360 and 374). They are conspicuous for their almost world-wide distribution [ p. 374 ] [ p. 375 ] and their luxuriance and abundance, along with a dense and varied undergrowth. Their most str iking representatives in number and size are the scale trees, a sort of evergreen having comparatively small needle-like leaves; some of these trees grew to over 100 feet in height, and to a diameter ranging up to 6 feet (see Pl., p. 377). There is in them an absence of rings of growth, indicating also an absence of seasons until near the close of Pennsylvanian time. Another remarkable group, the gigantic calamites or rushes (Pl., p. 381), grew to at least 60 feet long and 15 inches thick. They resembled the Uving cane brakes and bamboo thickets. These floras also included many fern-like forms, both delicate and hardy, some of which were climbing in habit, while others grew into majestic trees (Fig., p. 374) ; most of them bore seeds, but some were sporebearing and therefore true ferns.

In general, these forests must have reared their tops higher than 40 feet. They were of rapid growth and of soft and even spongy woods, as seen in the smooth, hard, and persistent bark, in the large and thin-walled cells of the woods, with a comparatively large amount of medullary and cortical tissue and large intercelliJar spaces, and in the many water-pores in the leaves.
Shades of green were the dominant color, and the monotone of the verdure was nowhere enlivened by bright flowers. Flowers, however, were present, but of a low order, insignificant in size and doubtless unattractive. Probably more than one half of the flora was spore-bearing (heterosporous) and we may safely regard most of the more common plants of the Coal Measures as seedless. Fertilization was not yet accomplished through the aid of honey- and pollen-eating insects as is so general among the hving flowering plants of the present, but was brought about in most of the coal floras by the rains and winds. At the time when the spore-producing trees and ferns were liberating their spores, the entire forest was covered with a greenish yellow or brown dust, and some of the coals are largely made up of these spores (White, and Jeffrey).
Spores (Pl., p. 377, Figs. 4-6) differ from seeds in that the latter give rise directly to sexed plants, while the former also develop into sexed ones, but these are short-lived and in turn give rise to the sexless, long-lived plants that bear the spores. In other words, the spore plants have an “alternation of generations,” while the seed plants have direct development. This matter will be taken up again later in the chapter.
The air was not scented with sweet odors, for there was no honey, but it is probable that resinous smells such as pervade living conifer forests were present.

Fig. 1, Alethopteris grandifolia, 2. Neuropteris fasciculata; 3, N. capitata; 4, Pseudopecopteris mazonana, with spore cases; 5, probably a seed-bearing fern-like tree, restored by Kidston; 6, Sphenopteris mixta; 7, Archaeopteris stricta; 8, Aneimites x 2; 9, same with a young seed. x 4; 10, same, one of the seeds x 3. Fig. 1—4, from the Illinois and Ohio State Surveys; Figs. 8—10 after David mite.
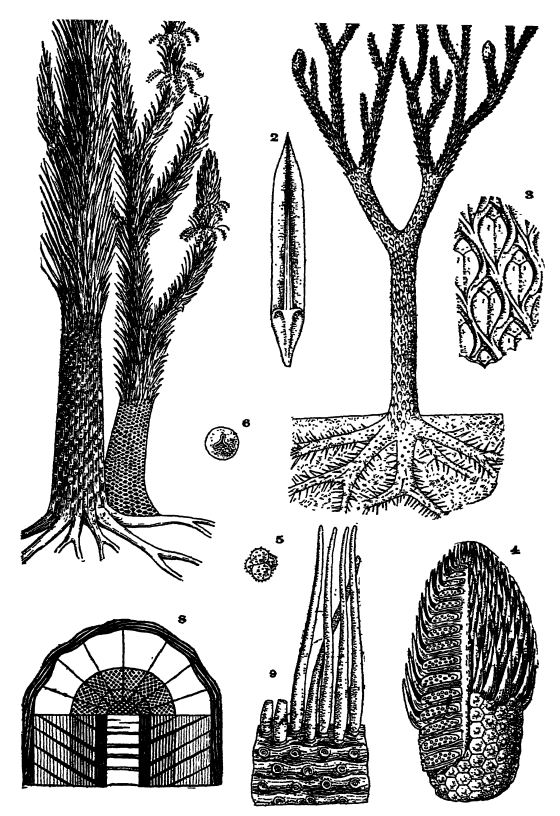
Fig. 1, restoration of the tree Lepidodendron, about x 1/100; 2, leaf of the sporebearing cone, about x ⅓, those of the branches being very similar but longer; 3, outer surface of tree, showing nature of leaf bases, about x ½ (L. cbovatum ) ; 4, sporebearing cone (Lepidostrobus) , with and without leaves, sectioned to show spores and spore-developing cases (sporangia) ; 5, bunch of four microspores; 6, macrospore; 7, two species of the tree Sigtllaria, restored, about x A; 8, section through trunk, showing relative thicknesses of outer leaf-bearing layer, two zones of cork-like bark, comparatively thin internal cylinder of wood, and medial pith; 9, roots of these trees (Stigmaria ficoides ) , with rootlets and scars of their attachment, figs. 1-6 after Steinmann; Figs. 7 and S after Zittel. (677)
[ p. 378 ]
In these forests of Pennsylvanian time might have been seen flying about the largest insects that have ever lived, great “ dragon-flies,” reaching a wing spread of 29 inches. Huge cockroaches abounded everywhere in great variety, giants of 4 inches in length not being rare (see Pl., p. 363, Fig. 4). As a rule, these insects were carnivorous, and did not transfer the pollen from one flower to another, with resulting cross-fertilization, as is so commonly done by living insects among present-day plants. The smaller insects were preyed upon by scorpions and spiders, the latter not making webs but living on the ground or in rotten logs, along with many myriapods, or thousand-legs. No insects of this time, so far as known, produced chirping or other sounds, and the soughing of the wind among the trees was interrupted only by the croak of amphibians in the marshes. Amphibians were common in the swamps, and it is probable that many small reptiles were running over the ground and about the trees. No larger land animals, such as we know, and no birds were to be seen.
Classification. — For our purposes, plants are most conveniently divided into (1) those which have no flowers but reproduce by spores and go through an alternation of generations (Cryptogams), as explained above; and (2) those which have flowers and reproduce directly from seeds (Phanerogams).
In this chapter we are to study the earliest land plants, and as these can be preserved as fossils only if they have fibrous or woody material, the paleontologist is not directly concerned with the lower Cryptogams.
All of the higher plants above the mosses in organization are characterized by having roots, and by the occurrence in their soft tissues of what are known as vascular bundles. These bundles are composed of tough, hollow, interlacing fibers, whose primary function is to serve as conducting channels (hence vascular, from vascidum, a small vessel) through which the various food materials pass, the water and salts ascending and the sugar and proteids descending; secondarily, they are of importance as forming supporting skeletons of the plants. All of the woody material is composed of these strands, and it is this tough substance which makes it possible for leaves and other parts of plants to impress upon the sediments the traces of their former existence.
The further classification may be expressed as follows:
Lower Cryptogams, spore-bearing plants
- Thallophytes, bacteria, fungi, most marine plants (algse)
- Bryophytes, mosses
Higher Cryptogams, also spore-bearing
- Pteridophytes, ferns (Pl., p. 376, Figs. 1-5)
- Arthrophytes, living rushes and Paleozoic calamites (Pl., p. 381, Figs. 1, 2)
- Lepidophytes, scale trees or lycopods (Pl., p 377)
[ p. 379 ]
Phanerogams, seed-bearing plants
- Gymnospenns, plants with inconspicuous, imperfect flowere
- Pteridospermophytes (or Pteridospermsj, fern-like plants (Pl., p. 376, Figs. 6—10)
- Cycadophytes, cycads (Figs., pp. 27 and 3S6)
- Coniferophytes
- Cordaites, ancient evergreen trees (Pl., p. 381, Figs. 3-6)
- Conifers, modern evergreen trees
- Gingkos, maidenhair trees (Pl., p. 381, Fig. 7)
- Angiosperms, true flowering plants
¶ Spore-bearing Flowerless Plants
¶ Ferns or Pteridophytes
Nearly everyone knows what a living fern looks like, but when this knowledge is applied to the Paleozoic plants of similar appearance, even the best of botanists cannot at present be certain in many cases that the form studied is a true fern; In the previous century, it was still believed that fully one half of the Carboniferous plants were true ferns and that these beautiful forms dominated the floras of that time. Due, however, to the accumulation of botanical knowledge, and more particularly to the discovery of specimens bearing the reproductive organs, it is now known that many of the so-called Paleozoic ferns bore seeds and therefore were of a higher organization and must be classified wholly apart from the true ferns. These were the seed-ferns, in many ways transitional between the spore-bearing ferns and the seed-bearing plants (see Pl., p. 376, Figs. 6-10). Probably more than one half of the Paleozoic plants formerly regarded as ferns (about one thousand species) win eventuafly be shown to be seed-bearing.
Living Ferns. — There are living to-day upward of six thousand different kinds of ferns, ranging from tiny, delicate forms to the beautiful tree-ferns of tropical lands, which grow to a height of 50 feet. The greatest variety occurs in the tropics, but herbaceous ferns are found under all climates, even in icy Greenland. As a rule, however, they prefer wooded humid regions. The fern plant is the sexless generation. Its spores are usually developed on the under side of the leaves in spore-cases, a number of which are always clustered together. In each one there are many microscopic spores which are all alike, and these are often developed in countless numbers (see Pl., p. 376, Fig. 4).
A few of these spores may germinate in the groimd, giving rise to a sexed plant, the prothallium, which is usually a heart-shaped, primitive, green shoot lying prostrate on the ground and often less [ p. 380 ] than 0.5 inch in length. On its under side are developed male organs that give rise to male cells or pollen, and female organs that when pollenized develop certain cells into sexless plants. We have here, therefore, an alternation of generations, that is, non-sexed plants giving rise to sexed plants, and these in turn producing the non-sexed ferns.
In the smaller herbaceous ferns there is a short, stout, undergrovmd stem that gives rise to many leaf stems, each of which develops curled up in a spiral, remaining so for a year or more. Subsequently the stems uncurl and grow rapidly into the fully developed leaves, generally called fronds. These may attain a great length, as in the tree-ferns, and are called fronds because the leaves are compounded of many leaflets.
In the tree-ferns there is but a single unbranched stem resembling a palm trunk, and the fronds, consisting of one or more leaves with their many leaflets, are developed at the top as a crown, each frond uncurling as it attains full growth. The trunk bears the scars of the fronds that have dropped off and beneath this outermost layer there is often a thick zone, the bark, which surrounds the more or less large, central, woody cylinder (Pl., p. 376, Fig. 5).
Fossil Ferns. — Ancestral ferns (Marattiales, or ferns developing from a single cell) were rare in the Devonian and are not common as fossils until the Pennsylvanian, where many species of the smaller herbaceous kinds occur associated with the tree-ferns (see Pl., p. 376, Figs. 1, 4). The latter kinds became conspicuous for the first time in the Pennsylvanian and were plentiful in the later half of this period and in the early Permian. The stem in some instances attained a height of 50 feet, and in certain forms the fronds developed in two or four parallel columns, but in most cases they were placed spirally around the trunk (see Pl., p. 376, Fig. 5).
¶ Bushes or Arthrophytes
Among living plants there is a small group of forms having a very wide distribution and popularly known as rushes (similar to Pl., p. 381, Figs. 1, 2). As a rule, they are small, less than 18 inches tall, appearing in wet places early in the year; in Central and South America and Cuba, however, a giant form occurs in groves, attaining a height of 40 feet, but with a stem that does not exceed an inch in diameter. This and all other living forms, about twentyfive in number, belong to the genus Equisetum (see also Fig., p. 468).
This living genus of rushes has in the ground a non-sexed, perennial, creeping and branching, horizontal root-stalk, that at intervals [ p. 382 ] gives rise to upright aerial stems and descending roots. The stems consist of a thin, externally striated, woody zone, wdth a large pith center. At regular intervals the stem is divided into internodia by transverse partitions called nodes, and from these arise the whorls of small or reduced leaves, vdth one or more longitudinal veins, and also the branches when such are present. The branches repeat on a smaller scale the general features of the stems. At the top of the stem or on the branches occur the spore-bearing cones.

Fig. I, calamite trees restored, the one on the left bearing a spore case, about x 1/100; 2, basal end of a calamite; 3, cordaite tree restored (Dorvcordaita), about x 1/100, also showing nature of catkin-like flowers; 4, different type of cordaites about x 1/40; 5, Trigonocarpon ornatum, a fruit or seed of a cordaite; 6. Rhabdocarpus apiculatus, seed of a oordaite; 7, branchlet of the living maidenhair tree (Ginaba biloba) male flowers, and on the Hght the mature seed.
Fossil Calamites. — In Paleozoic time, from the Devonian to the Permian, but more especially in the Pennsylvanian, lived a great variety of ancient rushes, the largest of which are known as the calamites (Pl., p. 381, Figs. 1, 2). They were prolific plants and White says that in America they attained a diameter of up to 12 inches and a length of more than 30 feet. In them the wood cylinder was far thicker than in living rushes, and specimens are kno’wm having 2 inches of bark outside of the same thickness of wood. The layers of secondary wood were added externally beneath the bark, a type of growth found in modern trees. The nodes were not equidistant throughout as in the living forms, but in the earliest growth were progressively farther apart, and the stems thickened rapidly. Calamites were also far more abundantly branched than is the case in living rushes, and their leaves were proportionately larger. The top of the trunk consisted of a crowded tuft of closely set circles of leaves, and the cones with reproductive spores occurred more or less abundantly on the smaller leafy twigs or at the top of the main stem. As a rule, the spores were all alike in the Paleozoic Arthrophytes, but in some cases they were differentiated into small and large types (heterospores, see Pl., p. 377, Figs. 5, 6).
¶ Scale Trees or Lepidophytes
Living Lycopods. — Lycopods is a group name for the very widely distributed, primitive, herb-like, evergreen plants known as ground pines, club mosses, and running pines, the latter being extensively used in floral decorations. There are about one hundred living species of the genus Lycopodium, the name, from the Greek for wolfs foot, being given because of the appearance of the roots. In the existing floras the lycopods never grow in dense masses to the exclusion of other plants, and therefore are ffot conspicuous. Their general appearance suggests the larger mosses or smallest branches of pines, since the leaves are always remarkably small.
Fossil Lepidophytes. — In the Paleozoic from Devonian to Middle Permian time, but more especially during the Pennsylvanian, the [ p. 383 ] Lepidophytes were the dominant plants, and attained gigantic proportions when compared mth the greatly reduced living lycopods, for the former were then at the climax of their evolution (Figs., pp. 360 and 374). These trees in Pennsylvanian times liberated their heterosporous reproductive germs in such tremendous amounts as to be a marked factor in coal making (see Pl., p. 377, Figs. 4-6).
In the late Paleozoic there were two main types of Lepidophytes, known as Lepidodendron or scale trees and Sigillaria or seal trees, terms which have reference to the scale-like appearance of the leafbases on the trunks and branches of these trees. Upward of one hundred species of Lepidodendron have been described, ranging from the Devonian to the Permian (Pl., p. 377, Figs. 1-3). The trunk tapered slowly and in some forms grew to a height of over 100 feet and a diameter of 3 feet. These trees in their general appearance differed from all modern ones in that the trunk and many of the branches divided into two forks rather regularly. The more slender branches were terminated, as a rule, by linear or oval spore-bearing cones 1 to 12 inches long (Pl., p. 377, Fig. 4).
The leaves of Lepidodendron were needle-like, always comparar tively small, but having a width at the bottom of 0.5 inch and a length of 6 to 7 inches (Pl., p. 377, Figs. 1, 2). On their xmder sides there were two lateral grooves, and in these hoEows were situated the minute but numerous breathing organs for the extraction of the carbon dioxide from the atmosphere. The leaves soon dropped off, and are usually found attached only to the young growing twigs. The leaf-bases were diamond-shaped and arranged on the branches and trunks in spirals, thus forming the characteristic mar k in g s of the trees. The leaf was attached in the upper part of the rhomb (Pl., p. 377, Fig. 3).
Of Sigillaria (Pl., p. 377, Fig. 7), about one hundred species have been described, and they differ from Lepidodendron in being rarely branched. Trunks have been found with a diameter, just above the roots, of 6 feet, and one attained a height of nearly 100 feet and was unbranched. At the tip, for about 10 feet, these trees were clothed with erect, rigid, grass-like leaves, which in most cases were like those of Lepidodendron, but sometimes were far larger and wider.
The leaf scars in Sigillaria were arranged in vertical rows with the scars of adjacent series alternating with one another. In many forms the surface of the tr unks was longitudinally ribbed, each of the ribs bearing a single row of leaf scars (see Pl., p. 377, Fig. 7).
[ p. 384 ]
Sigillaria are unknown in the Devonian and died out before the close of the Pennsylvanian.
The structure of the trunk and branches in the fossil Lepidophyies was peculiar in that the greater thickness beneath the outer layer with the leaf scars was composed of two kinds of cork-like bark surrounding a comparatively thin internal woody cylinder that may have been entirely filled with inward growing wood — the endogenous type of growth — or with more or less of pith (Pl., p. 377, Fig. 8)Secondary wood up to 2 inches thick was added to the outer cork beneath the leaf layer, and as this cork was also tough, the trees had considerable resisting power against the winds and storms of that time but were less stiff than the modern forest trees.
Basally, both Lepidodendron and Sigillaria terminated in from four to seven trunk roots that are thought to have spread almost horizontally in the ground, and there was no vertical or tap root continuing the trunk as in modern trees. These trunk roots each divided once or twice and tapered to a point, burrowing in every direction through the mass of decaying vegetation in the swamp. On their surface were circular scars arranged in sets of five, and to these were attached round tapeiing appendages radiating in all directions; these latter attained a length of 15 inches and served for the extraction of food and water from the ground (Pl., p. 377, Fig. 9). Such roots are known as Stigmaria (from the Latin stigma, a mark), and are very frequently seen in the underclays beneath coal beds.
The cones of Lepidophytes, while they looked much like those of modern evergreens or conifers, did not have seeds but were filled with spores (Pl., p. 377, Figs. 1, 4-6). The latter grew in large cases, which contained either minute spores, the microspores or fertilizing male parts (Pl., p. 377, Fig. 5), or large ones, the female macrospores (Pl., p. 377, Fig. 6), often forty times larger than the microspores. These two kinds of spores were developed either in different cones or in different parts of the same cone, and there were many more of the minute spores than of the larger ones.
¶ Seed-bearing Flowering Plants or Phanerogams
The seed-bearing plants, which are exceedingly varied and numerous, fall into two great divisions. The geologically older and more primitive group are known as Gymnosperms (or naked seed plants), in which the male and female parts may be in separate flowers or combined into one, but are always inconspicuous and often without bright color. The seeds develop in the open ovaries, which are [ p. 385 ] said to be naked because the pollen falls directly upK)n the ovules (eggs), while in the higher or true flowering plants (Angiosperms) the ovary is covered by a stigma which receives the pollen.
¶ Seed-bearing Ferns or Pteridosperms
In discussing the ferns on a previous page, it was stated that there were in the Paleozoic many plants having all the appearance of being true ferns but known to have borne seeds instead of spores. In other words, these plants had no alternation of generations to complete the developmental cycle, but were sexed plants, that is, they grew directly from the seed into either males developing pollen or females developing seeds which were fructified and developed into an embryo while attached to the plant. They had the general appearance of ferns and had seeds very much like cycads. The discovery of these Pteridospermophyta, or, in common parlance, Pteridosperms, is one of the outstanding achievements of the paleobotanists of the early part of this century. In the origination of seeds, a great forward step had been taken by the Pteridosperms, and it became the dominant feature in later floras (see Pl., p. 376, Figs. 6-10).
The seeds and pollen were borne on independent sexed plants. The seeds may have been small and occurred at the tips of the leaves or hung down on the xmder side at the outer ends of the lobate leaves, or, if large, were attached to the thick midrib of the frond; they never occurred in cones. The seeds varied in size from 0.25 inch up to at least 2 inches in length, often being long drawn out at the free end, where the entrance to the pollen chamber was located. The embryo or nut was surroimded by a husk that in some forms had a thick, fleshy exterior and a hard, stony, inner layer. Similar seeds also occurred in the cordaites, in the gingkos (Pl., p. 381, Figs. 5-7), and in the cycads.
Some of the seed-fems had long and slender stems, and climbed about other plants; many were herbaceous, and others (Psaronius) were as tall and stout as the tree-ferns. They had their origin in primitive ferns earlier than the Middle Devonian, and their climax of development was in the Pennsylvanian and early Permian, but none are as yet known beyond the Paleozoic.
¶ Cycads or Cycadophytes
In the modern warm-climate floras, the living cycads, of about 110 species, are the remainders of a once more diversified group, never very common in the Paleozoic floras, but dominant in those of the earlier half of the Mesozoic era. A striking living example [ p. 386 ] is the “ sago palm ” of Ceylon, the source of our edible sjigo. The significance of these ancient seed-bearing plants in relation to the other flowering plants we have learned largely from the studies of G. R. Wieland; they appear to have arisen out of the Pteridosperms, possibly as early as the Devonian. Cycadaceous plants appeared with the Aliddle Pennsylvanian but were not common until Triassie times.
The salient feature of cycads is that their short columnar woody stems, having very large pith centers, are encased in a thick armor of persistent leaf bases, with an intermediate felt-like mass. The trunks vary in size from very small ones to those of 60 feet in length in living Cycas. Cycads grow slowly and do not become mature for many years, and a trunk 6 feet long may be 1000 years old. They are at times wonderfully preserved in Mesozoic strata. The leaves are of various kinds but commonly of the pinnate paJm-leaf type, and are much used in floral decoration (see Figs., p. 27 and above).

¶ Conifers or Coniferophytes
Cordaites, or Large-leaved Conifers. — In the Paleozoic, beginning at least with the Upper Devonian, occur logs whose structure is [ p. 387 ] not very unlike that of modern pines or conifers. In the Pennsylvanian, casts of such logs are often seen in the sandstones, and in the roof shales of the coals the long strap-like leaves of these trees abound; in fact, they often form a good part of the coal. These plants, which are known as cordaites (after the paleobotanist Corda), included a variety of forms, and were the dominant Gjtunosperms of Paleozoic time. They were softwood evergreen trees, tall and slender, sometimes 120 feet tall and 3 feet thick. In such tall trees fully two thirds of the trunk was without branches, the upper third or fourth of the stem having a dense crown of them and abounding in simple leaves of large size (see Pl., p. 381, Figs. 3-6).
The leaves in the cordaites were always large, and some are known with a length of 6 feet and a width of 6 inches. Some were strapshaped, with the free ends either pointed or blimt; others were grass-like, with a length of 20 inches and a width of 0.5 inch. The leaf substance was thick and had parallel veins as in the living yucca (see Pl., p. 381, Figs. 3, 4).
While the woody trunk of the cordaites was very much like that of modern pines, it is distinguished from the latter by always having a central pith which may be of any diameter up to 5 inches. The new wood was formed beneath the thick bark, in other words, was added externally to the previous layers (exogenous growth).
The cordaites were related to the conifers, but, as is seen from the above description, the two groups were very different in their composition. The former were much branched only at the top, did not have cones with seeds but developed them in catkins, the leaves were not needle-like but strap-like, and finally the trunks had a pith center. The cordaites were most closely related to the gingkos.
The gingkos differ from the cordaites at once in the much smaller and lobate leaves, while the trunks are branched throughout nearly their whole length (see Pl., p. 381, Fig. 7).
Conifers or Evergreen Trees. — The evergreen trees, or true conifers of living floras, whose seeds are developed in prominent female cones and the pollen in male cones, are not known to be older in origin than the Permian, but are now the commonest of Gymnosperms. The order embraces the araucarias, the sequoias or California “ big trees ” (closely related but smaller trees can be traced back to the Permian), the pine, spruce, fir, juniper, larch, cypress, and yew.
[ p. 388 ]
¶ Angiosperms or True Flowering Plants
As the true flowering plants do not appear in Historical Geology until the Cretaceous, their description is deferred to the chapter on that time.
¶ Collateral Reading
E. W. Berry, Paleobotany: A Sketch of the Origin and Evolution of Floras. Annual Report of the Smithsonian Institution for 1918, 1920, pp. 289-407.
C. J. Chamberlaix, The Living Cycads. Chicago (University of Chicago Press), 1919.
J. M. Coulter and C. J. Chamberlain, Morphology of Gjunnosperms. Chicago (University of Chicago Press), 1910.
D. H. Scott, An Introduction to Structural Botany, Part II, Flowerless Plants. London (Black), 1904.
A. C. Seward, Links with the Past in the Plant World. Cambridge (University Press), 1911.
D. White, Fossil Flora of the Lower Coal Measures of Missouri. U. S. Geological Survey, Monograph 37, 1899.
D. White, The Stratigraphic Succession of the Fossil Floras of the Pottsville Formation in the Southern Anthracite Coal Field, Pennsylvania. U. S. Geological Survey, Twentieth Annual Report, Pt. II, 1900, pp. 749-918.
G. R. Wieland, Araucariales, Cycadales, and Cordaitales. Encyclopedia Americana, 1918, pp. 135-138, 351-360, 683-686.
| XXVII. The Pennsylvanian Period, the Time of Greatest Coal Making | Title page | XXIX. Coal and its Occurrence in Nature |