[ p. 405 ]
In the middle Paleozoic, and especially in the Devonian, when the dry’ lands had become fully clothed by plants — the substratum on which all animals are dependent for food — there was established a habitat capable of sustaining animal life, and destined to be mastered by worms, snails, spidem, insects, and other invertebrates, and finally by various groups of vertebrates. Among the latter, the first to find the land habitable were the ancestral Amphibia, and they ruled their various environments certainly from late Devonian until well into Pennsylvanian time. Out of these animals came more complex ones, a most wonderful variety of reptiles, which not only became lords of the lands, seas, and oceans before the close of the Permian, but shortly afterward invaded the air, and by their dominancy gave the Mesozoic or medieval world its title of the Age of Reptiles.
¶ Amphibia, Living and Fossil
Amphibia differ from fishes in having legs and not fins, each leg bearing fingers or toes. Nearly all of them breathe by gills when very young and may retain these organs throughout life; lungs and functional nostrils are, however, nearly always present in the adult, although among the salamanders limgs may be reduced to vestiges or completely suppressed (see Pl., p. 407, Figs. 1-4). They have a three-chambered heart, and a mobile muscular tongue.
Living Amphibia. — There are about nine hundred forms of living Amphibia, most of which are of the frog kind (Fig., p. 406). Frogs, toads, newts, sirens, mud-puppies, water-dogs, and land salamanders are types of living amphibians (Pl., p. 407). All are cold-blooded animals. The class name Amphibia, which means living a double life, was given them because many live both on the land and in the fresh water. Others, however, live entirely in the water, and, as a rule, all amphibians in their younger stages are wholly restricted to this element.
The sexes are always separate and in the great majority of species the small eggs are fertilized in the water and develop there [ p. 406 ] without further care from the parents. The development is, therefore, very much the same as in the fishes and very un l i k e that of the higher vertebrates. The eggs of frogs and toads, for example, develop into little animals commonly known as tadpoles or polliwogs, with a more or less large and rounded head and body terminating in a long and very flexible tail, which they wriggle in swimming, as do the fishes (see Fig., below). These tadpoles have gills which at first project from the sides of the head, but are later covered by an operculum. In about two months they attain a stage which is the equivalent of the lung-fishes. They then undergo a marked metamorphosis, the hind legs appearing first and later the front pair, which are hidden under the operculum. The long tail shortens through internal absorption and the gills are also absorbed or drop off. The lungs then appear, and for a time the young creatures are fully amphibious, breathing water through their gills and extracting the free oxygen from it, and also taking in air through the lungs; but soon the small frogs or toads take to the land and breathe air only (see Fig., above). In this metamorphosis, requiring from a few weeks to at most a few months, we see a recapitulation of Paleozoic history that consumed millenniums of selection of the most fit for their environment. In those forms that remain permanently in the water the transformation is not so great, and some of them resemble fishes in bodily form through [ p. 407 ] [ p. 408 ] out life, among these being the so-called mud-puppies and waterdogs common to the rivers of the Ohio and Alississippi valleys (see Pl., p. 407, Fig. 1).
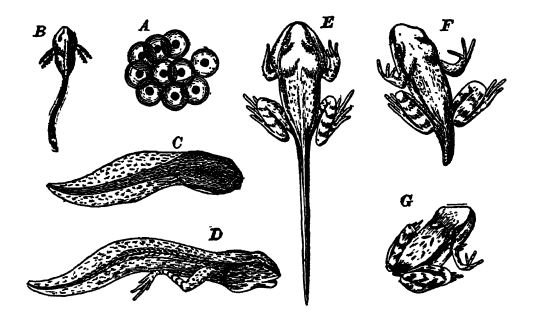
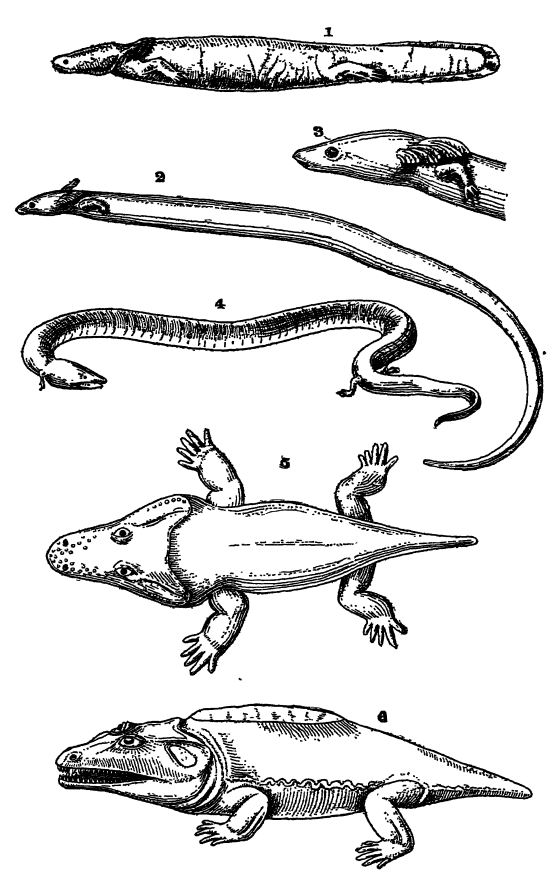
Fig. 1, mud-puppy (Necturus maculaius) of the Ohio and Missisippi rivers; 2 and 3, Siren lacertina, a small salamander, note the three pairs of gills and the absence of hind legs; 4, Amphiuma tridactyla, with both gills and lungs; 5, Trematops milleri, a stegocephaJian, 3 feet long, from Texas; 6, Cacops aspidephorus, a stegocephalian, 20 inches long, from Texas. Figs. 1 and 4 from Parker and Haswell; Figs. 2 and 3 after Lankester. British Museum Guide Book; Figs. 5 and 6 after
Respiration. — All Amphibia in their youth are provided with two or three pairs of external gills or internal ones with external gill-clefts, soft feathery outgrowths situated at the back part of the head and rich in blood-vessels (see Pl., p. 407, Figs. 1-3). Such gills are also present in the lung-fishes, and in the sirens and mudpuppies they may persist throughout life, though this condition is rather exceptional. In the salamanders of the land, and in all of the tailless Amphibia (frogs, etc.), the gills disappear and adult respiration is carried on wholly by limgs, as in the higher vertebrates. While the gills are present, the air passages to the lungs through the nose do not open into the mouth, but assume this position as soon as the gills vanish and the lungs become functional. The air is then taken in through the nasal openings and in the frogs is forced into the lungs in a swallowing manner. Many of the Amphibia can live a long time without food, and in the colder countries all the land forms hibernate through the winter in the mud at the bottom of streams and ponds, breathing during this time only through the skin.
Hearing. — In none of the fishes are there functional ears, though there are internal ones; in the amphibians, however, there are distinct external organs of hearing. These are best seen in the frogs, where the tympanic membrane of the ear-drum is a more or less large, circular disc embedded in the outer skin on the sides of the head. The cavity behind the disc, or drum, connects with the back part of the mouth by a tube known as the Eustachian tube, which is also present in all of the higher vertebrates.
Classification. — The living forms are divided into two subclasses on the basis of their body form: when tailless in the adult stage, they are called Anura (without a tall, see Fig., p. 406), while the primitive forms retaining the tail are called Urodela (having a distinct tail, see Pl., p. 407, Figs. 1-4). The Paleozoic amphibians, to be discussed later, are included under the term Stegocephalia.
The Anura include the frogs and toads so commonly known. They always have four legs, the hind pair of which are long and powerful, and the adults are without external gills or gill-clefts. They are almost cosmopolitan, occurring in all countries except the cold polar regions. This subclass is of comparatively modern origin, since the oldest fossil form is known in the late Jurassic, though in Cenozoic strata they occur far more frequently. They are the [ p. 409 ] most diversified of living Amphibia, and also the most specialized, but are not of particular interest in Historical Geology.
The Urodela or tailed Amphibia are of fax more significance in this connection, and even though fossil forms are not known older than the Jurassic, the subclass must have had its origin in the Paleozoic Stegocephalia. The Urodela, now practically restricted to the temperate parts of the northern hemisphere, are amphibians which as adults (salamanders, tritons, sirens, mud-puppies and axolotls) are often strikingly fish-like in appearance (see Pl., p. 407, Fig. 1). In the mud-puppies and axolotls there is an unpaired fin on the dorsal side, continuing around the diphycercal tail and along the posterior part of the ventral side, as in the lung-fishes. However, this fin does not have skeletal or bony rays, as in the fishes. Even though the Urodela are fish-like in appearance, they are readily distinguished by the fact that their paired appendages are not fins but fully developed legs, though small and weak, with all of the essential skeletal characteristics of the limbs of the higher vertebrates (study Pl., p. 407, Figs. 1-4). As a rule, there are two pairs of these limbs, the front or pectoral, and the hind or pelvic. In some forms, however, only the anterior pair is present, the posterior one having been lost, and in a few species all of the limbs are absent, these latter being the burrowing, worm-like Csecilians of tropical lands. The number of toes on each foot varies between five, the usual number, and two. Some forms, like the land salamanders, when matured may live wholly on the land and lose all trace of the gills, while others, such as the mud-puppies, remain in the water and preserve the gills throughout life. The Japanese and Chinese giant mud-puppy is the largest of all Urodela, attaining a length of 5 feet. One individual lived in captivity for over fifty years.
Stegocephafia, the Paleozoic Amphibia. — The term Stegocephalia means covered or mailed head and has reference to the fact that the upper surface of the actual skull was roofed over by more or less thick dermal bones. They are also known as the solid-skulled and armored Amphibia, and it is this armoring that at once distinguishes the Paleozoic and Triassic forms from all living ones. The chest was likewise provided with dermal armor, consisting of three large sculptured plates which represented a part of the bones of the pectoral arch of the fishes and the shoulder girdle of higher vertebrates. In some forms the entire body was covered with small overlapping scales, those of the ventral side being the thickest, or the scales being on the ventral side alone, while others (Branchiosauria) [ p. 410 ] were essentially naked and devoid of scales. Nearly all living Amphibia are naked and scales are but rarely present (Csecilians.) One group of the ancient forms have been called Labyrinthodonts, because the dentine of the large and conical teeth was much folded in labjTinthine undulations such as are first seen in fishes (p. 298).
The first trace of a limbed vertebrate, probably stegocephalianlike, is a foot impression (Thinopus) from the Upper Devonian (Fig., p. 331), but skeletons of the armored amphibians are not known in North America until Pennsylvanian times. They were then and in the Permian in their heyday. The whole stock vanished before the close of Triassic time.
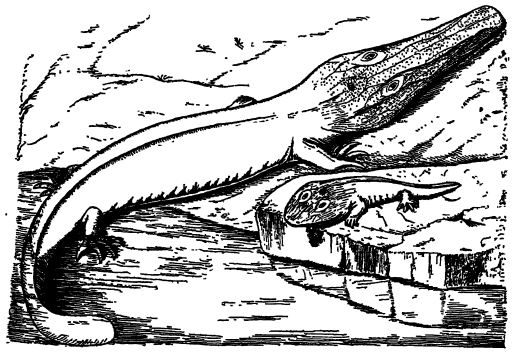
The most primitive Amphibia had small heads and weak limbs on long bodies that terminated in propelling tails like those of modern salamanders, but unlike these retained more or less of the bodilyarmor that came to them from their scaled and armored fish forebears. The large-headed forms seen chiefly in the late Pennsylvanian and early Permian were precocious descendants of primitive forms. The marked evolution of the ancient amphibians falls in with the time of their deplosnnent into all the varied en-vironments, passing from water and swamp habitats to transitional ones, and finally to those of the dry lands, and here becoming still more diversified [ p. 411 ] by burrowing beneath the surface and rising above it into the trees.
There are many kinds of stegocephalians known, the Coal Measures of North America alone yielding ninety species. In size they range from about an inch to more than 10 feet (Fig., p. 360). Some were lizard-like, small and active, with well-developed walking legs and relatively short tails; some were active swimmers, with long tails; others of medium size were thick-set and sluggish in habit, like the crocodiles (Fig., p. 410); while a few had the appearance of being gigantic, almost legless, tadpoles wrigghng about in the water. A tail was invariably present.
In the great majority of Stegocephalia there were two pairs of limbs, though leg-less forms like snakes are also known (see Fig., p. 360). The limbs were well developed but short and stumpy (see Fig., p. 410, and Pl., p. 407, Figs. 5, 6) . The bones of the front legs were like those of living salamanders in number, form, and disposition, and the hands were provided with four, or rarely five, usually short fingers. The hind limbs were nearly always longer and heavier, and bore five toes, of which either the second or third was the longest.
The heads were often broad and flat, with very wide mouths almost the full width of the skull, as in frogs and salamanders; but in some forms the heads were more or less elongated and even pointed, as in crocodiles. The heads of the latter, however, are also broad in the young stages, so that the broad head must be the primitive type. Of the former kind, Eryops of the Permian of Texas had a head 2 feet long and 18 inches wide and resembled a huge tadpole with a wide flat head, no neck, a thick heavy body, short legs, and a heavy flattened tail (see Branchiosaurus, Fig., p. 410). As Huxley has said, they “ pottered with much belly and little leg, like Falstaff in his old age, among the coal-forests.”
The Stegocephalia probably all lived either in fresh water or on the dry land, and some fed along the beaches of the Triassic seas. Certain of the small, active types are found in hollow logs of the Pennsylvanian of Nova Scotia. Probably the great majority of Stegocephalia were carnivorous and fed on shell-fish, worms, and other water invertebrates, but more particularly on fishes, reptiles, and small members of their own tribe.
The Third Eye of Stegocephalia. — In all well-preserved stegoeephalian sknlls the armor or roof bones and those of the true skull were pierced not only by the large lateral orbits in which the paired eyes were situated and by the pair of anterior nasal openings, but also by a single small orifice through the bone over [ p. 412 ] the brain (see Fig., p. 410). This aperture is of great interest, for in it was situated a third eye known as the pineal eye. Such an opening is also found in many fossil and some living reptiles (Sphenodon), and while the organ it contains can hardly be regarded as acting like a true eye in living reptiles, its great significance lies in the fact that it is a vestigial organ whose ancestry can be traced back at least to Pennsylvanian time. The rudiments of this eye are present in the brain of all living vertebrates, including man.
Ancestry of Sfegocephalia. — It is becoming increasingly difficult to distinguish the late Paleozoic reptiles from their stegocephalian associates. The most ancient of the solid-headed reptiles (Cotylosauria and Pareiasauria) are very aimilftr to the soli d-headed Stegocephalia. Of this type Osborn says: Bone by bone its parts indicate a common descent from the skull type of the fringefilmed fishes (Crossopterygia).” In other words, evidence is rapidly accumulating to show that the stegocephalians were not the most primitive limbed land vertebrates, but that they arose in an older stock (Protopoda), which gave rise on the one hand to the water-loving amphibians, and on the other to the reptiles, which became completely adapted to the 6ry land.
¶ Reptilia, Living and Fossil
In all of the vertebrates so far studied — fishes and amphibians — we have seen that their habitat is either wholly in the water, or that at the very least the small eggs are there laid and fertilized, and that the young are also born and spend the days of their youth in this element. All of the higher vertebrates remove themselves more and more from this habitat and none are developed in it directly from the egg. In other words, the reptiles, as a rule, are oviparous, laying large eggs like those of birds and provided with a more or less great quantity of food (yolk), and these eggs are fertilized before they are laid upon the land, where they hatch under the warmth of the sun. This is the most important and fundamental difference between the lower vertebrates, the fishes and amphibians on the one hand, and the higher vertebrates, the reptiles, birds, and mammals, on the other.
Living Reptiles. — All of the living animals known as turtles and tortoises, lizards and snakes, alligators and crocodiles, and the extinct types, the giant dinosaurs of Mesozoic time and the flying reptiles called pterodactyls, belong to the class Reptilia. The word reptile means creeping or crawling and has reference to an animal that goes on its belly like the snake, or moves with difficulty on short sprawling legs, like the alligator. There are, however, many reptiles that are in no sense creeping and crawling animals, as, for instance, many of the fleet-footed lizards, certain of the dinosaurs with their pillar-like legs, and the winged pterodactyls.
All living reptiles are cold-blooded animals like the fishes and amphibians, and their skin is never soft, but always more or less [ p. 413 ] [ p. 414 ] hardened by horny or bony material that occurs more often as scales than as armor plate. Even though many reptiles spend a large part of the time in the water, they are essentially land animals, as they are born on the land and breathe exclusively through lungs. Each animal has a pair of lungs, but in the elongated snakes the left lung is rudimentary and almost lost. In the snakes and in some lizards, legs are either wholly absent or mere vestiges buried in the flesh, and in such reptiles locomotion takes place by means of a wriggling movement either over the ground or through the water. Wherever legs are present, the Angers and toes have claws, a feature that is very rare among Amphibia.
In many of the reptiles eyelids are present, and in most lizards and fossil reptiles there is also the pineal eye. The organs of smell and hearing are also well developed, but in the reptiles the vibrating, or tjTnpanic, membrane of the ear-drum is no longer external, as in the frogs, but lies in a depression. With the exception of the turtles, nearly all reptiles have teeth; in Kving forms these are usually pointed and often recinved to serve for the holding of their animal prey. In many fossil forms, however, the teeth were adapted for cutting and more rarely for the mastication of food. In snakes and some lizards the tongue is slender and bifurcated and is protruded in a darting manner. In other reptiles the tongue is flat and immovable, as in crocodiles, being then attached to the floor of the mouth.
Development of Reptiles. — In some living lizards and snakes, and rarely among extinct forms, the females are viviparous, that is, they give birth to fully formed, living young. The great majority of reptiles, however, are oviparous. The eggs resemble those of birds, but are, as a rule, rounder and have a tough, parchment-like, porous covering, or in some cases a calcareous shell. As these eggs are very different in their development from those of fishes and amphibians, they need to be described in more detail.
Since the eggs of reptiles are large and develop on the dry land, they must naturally be very different in intemal structure from the small ones of amphibians which hatch in the water. In the latter the embryos have functional gills for use in the water, but in the reptiles a wholly different organ has been originated to provide the developing young within the egg with the necessary oxygen. This sac-like embryonic organ, known as the allantois, passes on to the embryo the oxygen received through the porous shell, and carries off by the reverse process the carbonic acid gas; in other words, its function is respiratory (see Figs. A-E, p. 415),
[ p. 415 ]
“ The unhatched or unborn reptile breathes by raeans of a vascular hood spread underneath the eggshell and absorbing dry air from without. It is an interesting point that this vascular hood, called the allantois, is represented in the amphibians by an unimportant bladder growing out from the hind end of the food-canal. A great step in evolution was implied in the origin of this ante-natal hood or foetal membrane and another one — of protective significance — called the amnion, which forms a water-bag over the delicate embryo. The step meant total emancipation from the water and from gill-breathing, and the two foetal membranes, the amnion and the allantois, persist not only in all reptiles but in birds and mammals as well ” (J. A. Thomson in Outlines of Science).
Reptile eggs are large and contain a great deal of yolk, on which the embryo lives and grows. At one pole of the egg lies the fertilized germinal vesicle which develops into the embryo (Fig. A, opposite). During its earliest growth there is formed at either end of the elongate embryo a two-layered, crescent- shaped fold, called the amniotic fold, which arches over the embryo and finally unites to cover it with a protective [ p. 416 ] hood. This latter is the amnion, and between it and the embryO there is a shallow cavity containing a watery amniotic fluid, bathing and protecting the outer surface of the embryo (Fig. B, p. 415). As the amnion occurs only in the eggs of reptiles, birds, and mfl.mma.ls , these groups are known collectively as the Amniota, and the fishes and amphibia, which lack it, are called Anamnia, meaning without the amnion.
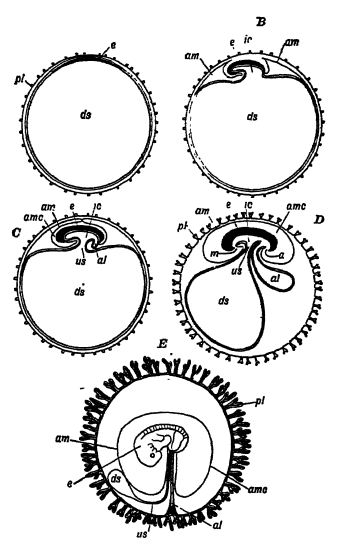
a. gnus; at, allantois; am, amnion; amc, amniotic cavity, ds, cavity of embryonic vesicle, later the yolk sac; e, embryo; e, intestinal cavity; m, mouth; placenta, an organ found only in mammals, formed Of walls of the uterus and embryonal membrane; both are abundantly supplied with blood-vesels, and while there is no direct continuity in the two blood streams, nutrient materials and respiratory gase are exchanged by osmoäs; u, umbilical stalk. After Köniker.
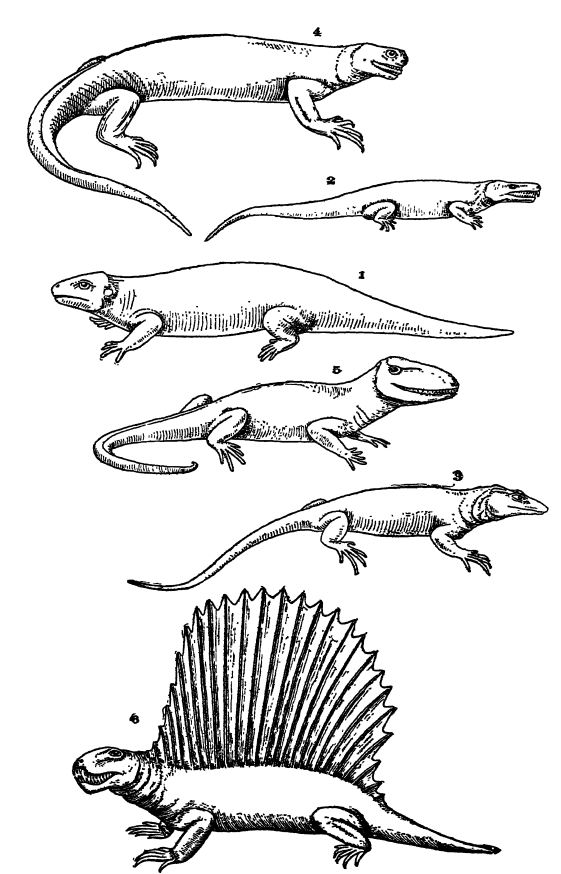
Paleozoic Reptilia. — Most of the Pennsylvanian and early Permian reptiles were plump, sluggish, more or less sprawling animals, basking often pn the land in the hot sun. In many ways they still resembled the stegocephalians, their associates, but had a marked tendency toward a reduction in the size of skull and toward loss of body armor. In all the forms the feet terminated in five fingers or toes. Few appear to have been swift of foot, and some, the “ ship lizards ” or “ fin-backed lizards,” bore a curious, very high, dorsal, median crest (Pelycosauria, Pl., p. 413, Fig. 6). “ These structures ” say Osborn, “ may have developed through social or racial competition and selection within this reptile family rather than as offensive or defensive organs in relation to other reptile families.” Most of the late Paleozoic forms were fierce-looking animals because of their large, recurved, holding teeth, and all of the American representatives seem to have been carnivorous in habit, feeding on insects, armored fishes, armored amphibians, and other reptiles. Others had crushing teeth, indicating a diet of shellfish and crustaceans.
Osborn says that the environment which changed the primitive Amphibia of the late Paleozoic into the Reptilia of the Permian was a warm, terrestrial, and semi-arid region, favorable to the development of a sensitive nervous system, alert motions, scaly armature, slender limbs, a vibratile tail, and the capture of food both by sharply pointed recurved teeth and by the claws of a five-fingered hand and foot. This evolution is as marvellous and extreme as the subsequent one of the mammals. Before the close of the Permian the reptiles had mastered all the parts of the lands, and were taking possession as well of the waters of the lands and seas.
It seems probable that the Reptilia arose even earlier than earliest Pennsylvanian time, for in the latter part of this period occurred not only true reptiles but also highly specialized forms. From the Pennsylvanian and older Permian of Texas, Oklahoma, and New Mexico, Williston and Case have made us acquainted with many different kinds of primitive Amphibia, and associated with them is even a greater and more complex society of primitive [ p. 417 ] Reptilia, animals that attained a ?TiayiTrmni length of 8 feet (see Pl., p. 413).
The order Cotylosauria embraces the solid-skulled reptiles, so called because the head was armored with sculptured plates (Pl., p. 413, Figs. 1, 2). They were the most primitive of reptiles. In build they were low and stout, slow of movement, and clumsy in walking, but since they had rather long tails, they swam well. All were carnivorous, though some fed partially on plants. Their life range was from late Pennsylvanian to late Triassic, and they are known chiefly in North America and Europe, though some occur in South Africa.
The order Theromorpha (means wild beast), which includes the Pelycosauria fin-backed lizards a highly specialized group of wholly land-living forms. They were active and Kzard-like, with rathet long tails, and some among them attained to a length of 8 feet. The fin-backed lizards ” were plump and bore a more or less high dorsal bony crest or fin. All were fiercely carnivorous. Their life range was from late Penns 3 dvanian into late Permian and they were in greatest abundance in North America, and less common in Europe and Africa.
South Africa is another region in which are preserved many excellent skeletons of ancient amphibians and reptilians. They occur commonly in the very thick Karoo series (up to 9500 feet) of continental shales and sandstones, and of Permian and Triassic times. According to Osborn, the greatest number of forms are found in the early and middle Permian, when the world already had a wonderful society of reptiles.
This African reptile assemblage included great round-bodied herbivorous forms (pareiasaurs), having massive limbs and round solid heads; agile ones with large eye-sockets and adapted to swift running (dromosaurs=ninning reptiles), terrible reptiles” with mammal-like heads (Theriodontia= carnivorous teeth), and many slender-limbed predatory tirpes with sharp canine-like teeth. There were also giant carnivorous reptiles (dinocephaIians= “terrible headed”), very massive animals with a highly arched back, broad swollen forehead, and short wide jaws provided with marginal teeth. Surpassing even these in size were the “ lawless toothed reptiles ” (anomodonts), in which the skull ranged from [ p. 418 ] a couple of inches to a yard in length, and the jaws were sheathed in horn and hooked like those of turtles. Finally in the early Triassic appeared the dogtoothed ” reptiles (cynodonts), the most mammal-like of all.
Progenitors of Higher Reptiles. — As has been said, the most primitive reptiles are the Cotylosauria. Lull tells us that this order, as seen in Limnoscelis (Fig. 2, p. 413), may well have been the central stock out of which could have evolved directly or indirectly the lizards, alligators, and dinosaurs. The cotylosaurians were swamp-living, sluggish, long-tailed reptiles having four legs of equal proportions.
Progenitors of Mammals. — In the Permian and Triassic of Africa and North America are found carnivorous reptiles of the suborder Theriodontia. They range in size from small forms to those as large as a tiger. They are of great interest because the African forms are regarded as having given rise to the lowest or egg-laying mammals, while in the American forms originated the higher reptiles. Their teeth were differentiated and localized, as in mammals, into incisors, canines, and molars (see Fig., p. 417).
¶ Collateral Reading
E. C. Case, The Permo-Carboniferous Red Beds of North America and their Vertebrate Fauna. Carnegie Institution of Washington, Publication No. 207, 1915.
E. C. Case, The Environment of Life in the Late Paleozoic in North America; a Paleogeographic Study. Ibid., Publication No. 283, 1919.
R. L. Moodie, The Coal Measures Amphibia of North America. Ibid., Pub lication No. 238, 1916.
H. F. Osborn, The Origin and Evolution of Life, pp. 177-233. New York (Scribner), 1917.
S. W. Williston, American Permian Vertebrates. Chicago (University of Chicago Press), 1911.
S. W. Williston, Water Reptiles of the Past and Present. Chicago (University of Chicago Press), 1914.
S. W. Williston, The Phylogeny and Classification of Reptiles. Journal of Geology, Vol. 25, 1917, pp. 411-421.