| XLII. The Evolution of Mammals and the Rise of Mentality in the Cenozoic | Title page | XLIV. The Evolution of the Elephants |
[ p. 624 ]
¶ The Horses
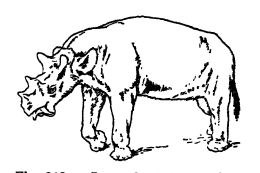
In demonstrating the truth of evolution, the horses, above all organisms, are the best illustration of the working out of this doctrine by means of natural selection and adaptation to environment. They are the “ show animals ” of evolution, since their history running back through millions of years is now well known. In this way the famous Yale University Collection, assembled by Professor Marsh, did much to establish the truth of Darwinism (study Fig., p. 627).
Huxley many years ago said that the horse must have been derived from some quadruped which possessed five complete fingers or toes (called digits) on each foot and which had the bones of the forearm and of the leg complete and separate. Moreover, that if the horse has thus been evolved, and the remains of the different stages of its evolution have been preserved, they ought to present us with a series of forms in which the number of digits becomes gradually reduced, the bones of the forearm and leg take on the equine condition, and the form and arrangement of the teeth successively approximate to those which obtain in existing horses. Since Huxley’s time, nearly all of the missing links in the evolution of the horses have been found and nowhere is this history so complete as in the Cenozoic formations of the Great Plains of the United States.
The horse is the most useful and beautiful of man’s domesticated animals, and has been one of the greatest factors in his civilization. In the early history of man he fed on the horse, and at Solutre, in the department of Sa6ne-et-Loire, France, there is one pile of horse bones estimated to represent 80,000 individuals. Later the horse became man’s chief means of travel and his beast of burden in agriculture and warfare. The horse is also among the most perfect and swiftest of organic running machines, as man loves to demonstrate in the race horse. As migrants into all continents, and in adapting themselves to varied environments — from torrid to arctic climes — horses have had but two equals — elephants and man. In [ p. 625 ] their wild state, horses are now restricted to the open arid plains of central Asia and Africa, since the mustangs and broncos of North and South America are descended from domesticated horses run wild since the days of the Spanish explorers.
Distinguishing Characters. — The horse family (Equidse, from Equus caballus, the living horse) includes the living horses, zebras, and asses. They belong to the odd-toed hoofed mammals (Perissodact 3 da), in which the axis of the foot lies in the third digit. Horses are characterized by their veiy long and slender feet, each composed of but a single functional toe, the third digit. The hoof is the equivalent of the nail or claw of the third finger or toe in other animals. Horses therefore walk upon the ver 3 ’ tip of the toe, in fact, on the third finger nail, the wrist being what horsemen call the “ knee ” and the heel the “ hock.” As the third toe in each limb supports the entire horse, it is necessarily much larger than in animals in which the weight is distributed among several digits. There is, however, on each side of the fxmctional digit, i.e. the “ cannon-bone,” a slender bone known as the “ splint bone.” These are the vestiges of the second and fourth toes of the original five in the ancestors of horses (Fig., p. 627). Curious as it may seem, domestic horses are sometimes born with two or three supernumerary toes on one or more feet, and such a one Caesar is said to have ridden in battle. The whole structure of the horse is preeminently adapted to swift running, and every part of the skeleton has been modified and specialized to that end.
The teeth of horses are as peculiar to them as are their one-toed feet (Fig., above). The molars are long, square prisms which grow [ p. 626 ] up from the gums as fast as they wear off on the crowns. This wear is accelerated because of the silica in the grasses eaten, and by the sand of the ground that is taken in while feeding. The grinding surface exhibits a peculiar and complicated pattern of edges of hard enamel, between which are softer spaces composed of dentine and of a material called cement, much like dentine in quality but softer and formed in a different way. The dentine is formed on the inside surfaces of the enamel while the tooth is still within the jawbone; the cement is deposited on the outside surfaces of the enamel after the tooth has broken through the jawbone and before it appears above the gum.

Evolution. — The horse family has been traced back to near the beginning of the Cenozoic mthout a single important break. When the little four-toed “ dawn horses ” (Eohippus), no larger than a small dog, appeared in western North America early in the Eocene (Wasatch), the land stood far nearer sea-level than it does now and the climate, though at first with winters, soon became warm and equable throughout the year. Then for a long time the seasons were very much alike and the climate tropical and moist enough [ p. 627 ] [ p. 628 ] to induce extensive areas of forests, at least over the Cordilleras (Laramide mountains). Over the Great Plains, however, the climate was drier and here were great grassy open plains. With the Miocene the whole of western North America began to rise, attaining culmination of elevation in the Pleistocene. Along with these topographic changes the climate became cooler, drier, and eventually icy cold. To all of these changes in the environment the horses adapted themselves or migrated into more favorable habitats, and in so doing changed from the smaller many-toed forms to the larger, fewer-toed, swifter, and more intelligent ones.
At first the many-toed horses browsed in the forests where they were an easy prey to the carnivores of the time, but with the diminishing of the forests and the appearance of the drier grassy plains, they spread for protection into the open plains, and here they developed more and more speed. With the elongation of the lower part of the limbs and the development of the sprinting habit of getting quickly up on their toes, came the gradual loss, through disuse, of the additional toes, and an equally remarkable change in teeth from a short-crowned browsing to a long-crowned (grazing) type. In this way the horses became one of the most highly specialized of animals adapted to a particxilar environment.
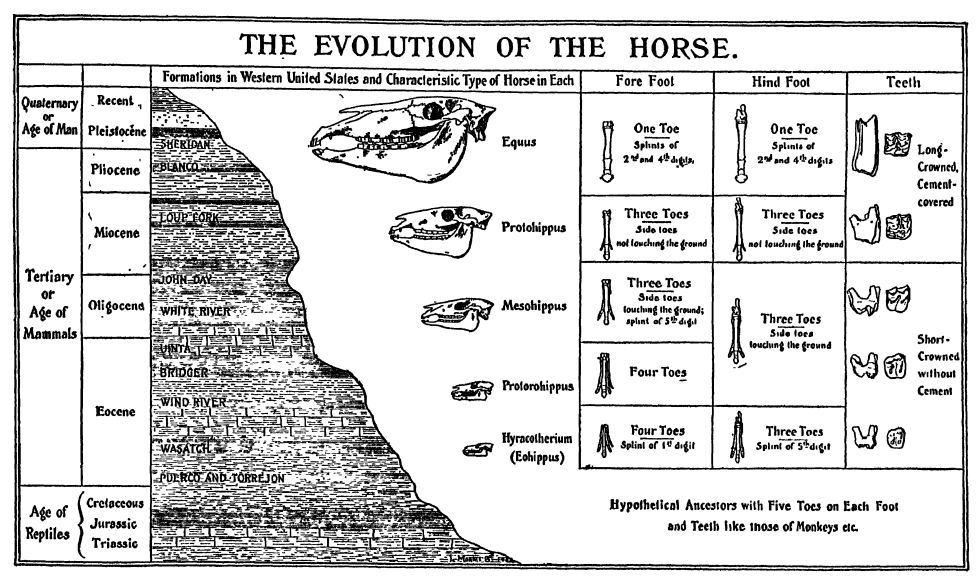
Where the horse family first originated is not known. The “ dawn horses ” appear at about the same time, and in the same state of evolution, in western Europe and North America. In Europe they soon died out (Eocene) but North America throughout the Cenozoic was their generating center. Curiously, however, even though horses were present throughout the Pleistocene in both North and South America, they had all died out at some time before the advent of the red men. Our present wild horses are feral, that is, had domesticated ancestors, and those of Asia, Africa and Europe are the descendants of early Miocene horses that spread from North America to Siberia by way of Alaska. Late in the Miocene the North American horses spread into South America.
Matthew recognizes twelve stages in the evolution of the horse family, found in as many different and successive geologic formations. Besides the main line of descent which led into the modern horses, asses, and zebras, there were several collateral branches which have left no descendants.
Scott summarizes the long and marvelous development of the horse family as follows: (1) In size there was a somewhat fluctuating increase, leading by slow gradations from the diminutive horses of the Lower Eocene, about the size of a fox-terrier, to the great [ p. 629 ] animals of the Pleistocene, about 14 hands tall. (2) The molar teeth, originally low, cusped, and with roots, gradually changed from the browsing type to the very long, prismatic, complex, grazing teeth, and the lower jaws grew in depth to accommodate this elongation. (3) The face grew relatively longer and the ej-es were progressively shifted farther back. (4) The short, neck was greatly elongated and the individual vertebrae modified so as to give flexibility with no loss of strength. (5) The limbs grew relatively much longer, the bones of the fore-arm and lower leg were fused together, the one on the inner side (radius and tibia) e nlar ging to csirry the entire weight, and the external one (ulna and fibula) becoming more or less atrophied. (6) The feet were much elongated and the median or third digit of each gradually enlarged until it carried the whole weight, at the same time modif 3 nng the shape of the hoof so as to fit it to be the sole support of the body. The other toes gradually dwindled and became functionless, though often retained as splints. The first digit was first lost, then the fifth, then the second and fourth were reduced to dew-claws and finally to splints. Thus the pentadactyl horses of the Lower Eocene were transformed into the monodactyl species of the Pliocene and Pleistocene.
Brain and Mentality. — The brain of living horses is large and richly convoluted, implying a high intelligence, but it is not equal to that of the elephant. The docility of the horse and its ability to learn are notable. On the other hand, it is emotional, and its psychology is largely linked up with its normal mode of defense — flight — since the first impulse of a domestic horse on seeing any incomprehensible thing is to run away. In the wild state this same impulse is of the greatest posrible aid as a means of survival. (Lull.)
Ancestor of Horses. — When the great English paleontologist, Richard Owen, described the little five-toed “ dawn horse ” of Europe (Hyracotherium) a type closely allied to the American “ dawn horse ” (Eohippus), he did not at all know that he had the ancestral stock of the horses, so unlike was it to the modern form. With the subsequent finding in North America of stages of development between it and other horses, it became plain that Hyracotherium was a generalized form descended from a stock that gave rise not only to horses but also to tapirs and rhinoceroses. In this is seen how an ancestral stock deploying into different environments leads eventually into animalR looking very unlike one another and yet having within their bodies structures which show their relationships. “ The conclusion is unavoidable that horse, rhinoceros and tapir, three races [ p. 630 ] widely different to-day, are derived through progressive changes from a common ancestral type ” (Matthew).
Geologic Succession. — The most ancient known member of the family EquidjB was the “ dawn horse,” which swarmed in the forests and glades of the Low’er Eocene (Wasatch). They were graceful little creatures no larger than a dog, or about 11 inches tall at the withers. They had on the front feet four functional digits and the vestige of another, while the hind feet had three toes and two tiny splints, the vestiges of the first and fifth digits. This evidence clearly indicates that Eohippus originated in ancestors with pentadactyl feet. The molars were cusped and short and of the browsing type; tiie neck was very short, the body long, with an arched back, the limbs and feet short, and the hind limbs much longer than the fore (Fig, p. 626).
The Oligocene horses were intermediate in development between those of the Eocene and Miocene. In the Lower Oligocene (White River) the largest ones were of the size of sheep (Mesokippus), and while the teeth were low-crowned and of the browsing type, they were changing toward those of the grazing animal. The front feet had three functional toes, and even though the middle one was the largest, the two lateral digits touched the ground; the hind feet were also threetoed, but there were no splints present (Fig., p. 627).
In the Middle Miocene the three-toed horses were still mostly browsers, though some of the progressive ones were changing rapidly toward grazers. It is here that we get the intermediate forms (Merychippus) and note the splitting up of the [ p. 631 ] horse family into several independently developing phyla. The Lower Pliocene had both progressive and conservative types of these animals. In the Middle Pliocene, among the herds of horses roaming the plains, there were three phyla of three-toed grazing horses, but the lateral digits no longer touched the ground; while they were “ dew-claws,” however, they were still at times of some supporting value. La the Upper Pliocene occurred the last of the three-toed browsing and grazing horses, and here lived also the first one-toed member of the genus Equus. In the earlier half of the Pleistocene there were at least ten species of - the same genus, and among them horses larger than any now living. (Study Fig., p. 630.)
[ p. 632 ]
¶ The Camels
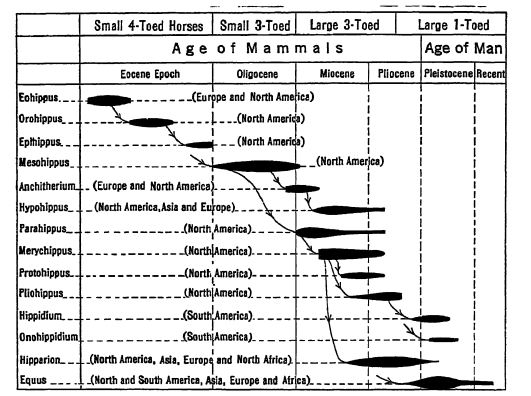
The family Camelidse now includes the two species of camels of the desert areas of central Asia and the Uamas and guanacos of the higher and colder parts of South America. These strange animals had their origin and essential evolution in western North America and their history runs parallel with that of the horses. They represent adaptations to dTj’ climates and open sandy plains. Camels belong to the even-t oed division of hoofed mammals, the Artiodactyla. Here the axis of the foot lies between the third and fourth digits and not, as in the horses, in the third digit.
Camels first appeared in the Upper Eocene in a form about the size of a domestic cat, and then there was a long line of them throughout the Cenozoic. In the Lower Miocene, the camels, like the horses, began to diversify, and large camels were still plentiful in North America during the earlier half of the Pleistocene but soon thereafter became extinct. The llamas spread southward through the tropics into South America in the Pliocene, and at the same time the camel stock radiated northward and finally across the AlaskaSiberia land bridge into Asia and thence into Africa.
Scott states that the mode of evolution shown by the camels differs in no significant regard from that seen in the horses. There was the same increase in bodily stature and in the relative lengths of limbs and feet, a diminution in the number of digits from the original five to two in camels (Fig., p. 631), and to one in horses (Fig., p. 627), and a similar development of the high-crowned grinding teeth from the low-crowned browsing type. The two families of camels and horses each arose in a single series.
¶ The Short-footed Amblypods
Among the archaic mammals of the Eocene there was a stock of browsers, sluggish in habit, clumsy in the mechanics of their skeleton, but abundantly represented by species and genera. These are called Amblypoda, a name which has reference to their short or blunt feet. Their legs were stout and pillar-like, and the short feet had five toes with elastic pads as in elephants. like all archaic mammals, their brains were small in proportion to the size of their bodies.
Amblypods probably originated in North America during the late Mesozoic, since the oldest forms are known in the Paleocene. They were common in the Wasatch, grew to larger and larger size and became more diversified during the later Eocene, and died out [ p. 633 ] before the close of this epoch in the Bridger. They are very striking and characteristic mammals in the North American Eocene, and but few of them (Coryphodon) migrated into western Europe.
The most striking of the ambhypods were the uintatheres, so named from the genus Uintatherium (see Fig., below), found in the Uinta Mountains of Utah, ilany of the kinds grew to the size of small elephants, 7 feet tall at the shoulders, and in the general shape of their broad legs and feet, but not their heads, resembled the proboscidians. The uintatheres, Scott says, were veritable giants and the most fantastic animals of their time. They were once regarded as proboscidians but it is now established that the two groups are in no way directly related. They were, however, the elephant-like animals of the North American Eocene.
The heads of uintatheres were large, and many-homed, and, curiously, the tops of the skulls were hollowed or basin-like, a feature that is characteristic of them. Of true horns sheathed with horn, as in rhinoceroses, there was a pair over the nose, and over the eyes and ears were other pairs of more or less high bony knobs thought to have been covered only with thick skin. Another striking characteristic, but restricted to the males, was the development of the upper canines into two formidable curved and sabre-like tusks. In order that these tusks should not be broken off in use, the lower jawdeveloped wide bony flanges for their protection. Their use is unknown. In both sexes the upper set of incisors were suppressed as in many living ruminants. The brain in uintatheres was “ absurdly small ” and their low mentality among mammals is comparable to that of dinosaurs among reptiles. It may have been the chief cause for their extinction, since they had to compete with the ever rising hordes of larger-brained and therefore more alert modernized mammals.
¶ The Giant-beasts or Titanotheres
In the Lower Eocene, among the immigrant modernized mammals there appeared an odd-toed imgulate (Eotitanops), smaller than a sheep and in appearance suggestive of a tapir. This ancestral form, Osborn says, evolved into eleven principal branches, the deployment [ p. 634 ] beginning in the Bridger, continuing in the higher Uinta, and vanishing at the climax of its development in early Oligocene (White River) time. The most significant evolution of these animals therefore takes place within 200 feet of Oligocene strata. Late in the Eocene they spread into Mongolia (Protitanotherium) but apparently did not live long there.
These are the titanotheres, “giant-beasts,” which are very characteristic North American Cenozoic mammals. It is only the later forms that attain the size of small elephants, but Brovtotherium, aside from the elephants, is “the most imposing product of Tna.rmna.1ia.n evolution.” Why the titanotheres failed when they were at their best is not known, but it may be that the drier climatic conditions of the Miocene, and the changing of their forest browsing habitats to open grass lands had already begun and if so may have been the cause of their xmdoing (see Fig., above).
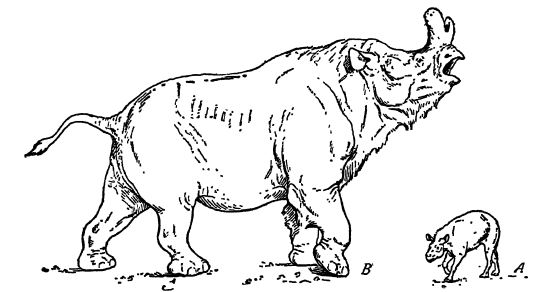
The titanotheres were heavy in body, with columnar legs and short feet, the latter supported on thick pads as in elephants. In all of them the front feet had four toes and the hind three, and in the older and smaller forms the toes and hoofs were more prominent. Their most characteristic single feature lies in the evolution of the head. In the older ones the skull was small, long and narrow, and devoid of knobs. The canine teeth, however, were prominent as tusks and the incisors were used for browsing on the vegetation. Evidently the tusks in the early forms were used as defensive weapons. In the Uinta, Scott sa 3 rs, the titanotheres were larger and had small knobs over the eyes that with time steadily enlarged and [ p. 635 ] shifted forward, until in the White River these bony horns had attained great size and were situated on the nose. While the horns were enlarging, the skull was being modified to support their weight and to endure better the shock of impact when they were put to use. When the horns had become weapons, the tusks dwarfed into insignificance and the front teeth were no longer used for cropping, this being done by the tongue and upper lip. In the fullness of the titanothere development, the heads were long, very broad, large, and massive; the profile deeply concave, resembling that of some fantastic rhinoceros with nasal horns. In body and limb as well these derived forms resembled large rhinoceroses.
The brain in the largest of the species was very small, no larger than a man’s fist, indicating that these great beasts “ must have been incredibly dull and stupid, surpassing even the modern rhinoceroses in this respect ” (Scott).
¶ The Rhinoceroses
The rhinoceroses belong to the odd-toed hoofed mammals; they are generally three-toed, and typically thick-skinned. As a rule, they have but little hair, although there was a woolly species contemporaneous with man during the Pleistocene in the cold climate of Siberia and northern Europe (Ceratotherium antiquitatis). They are browsers and grazers and live in forests, steppes, and marshes. They have large heads, short necks, and very long massive bodies, and their limbs are short, stout, and columnar, like those of elephants. The living forms stand from 4 feet to 6 feet and a half tall at the shoulders, and the single-homed species occur in India and Java, while those with two horns tandem, one on the nose and the other on the forehead, live in Africa and Sumatra. None of the fossil American forms was as tall or as heavy as the largest living form. The horns of rhinoceroses are peculiar in that they are neither hollow as in cattle, nor of bone, but are solid dermal growths made up of agglutinated hairs, and for this reason are never found fossil. Their presence in fossil forms is always indicated, however, by thickened and roughened nasal bones.
Since Middle Pliocene time there have been no rhinoceroses in North America, and yet this continent may have been not only the place of their origin, but that of their most significant evolution as well. The origin and development of the ancestral forms in North America and later of the trae rhinoceroses of the Old World is a very complex history, much more so than that of the horses and titanotheres. Ehinoceros-like Tnfl.TnTna1s appeared in America early in the Eocene [ p. 636 ] in small, active, and generalized forms (Hyradiyvs, Fig., p. 638) that in the course of the Cenozoic deployed into at least eight branches. These are again grouped into four main lines of evolution, namely, (1) the ancestral stock of small, defenseless and hornless, running rhinoceros-like forms; (2) a specialized and large, shortlived, aquatic type that lived in rivers and lakes; (3) the gigantic baluchitheres of Asia; and (4) the main stem of true slow-moving, huge, usually homed rhinoceroses. In Miocene and Pliocene times, these fl.niTna.1s had their widest distribution, living then in all continents except Australia and South America. The ancestral and aquatic stocks died out in North America during the Oligocene, and all of them early in the Pliocene.

Hornless forms arose first, and later on at different times in the Cenozoic appeared homed ones. The horns may be single, or double in a transverse pair (= pair-horned), or arranged one behind the other along the median line of the head (= tandem-homed); usually they are placed over the nose, but in some the horn is on the forehead. The presence of horns, as in the titanotheres, led to changes in the shape of the skull (see Figs., above, and p. 637).
The largest land mammal so far discovered was a hornless form of rhinoceros known as Bdiichitherium. It lived in either late Oligocene [ p. 637 ] or early Miocene time, and was first found in Baluchistan, but has since been collected in Turkestan and Mongolia. Osborn says it stood about 13 feet tall at the shoulders, and was about 25 feet long; in comparison all other rhinos are small. The neck was long and horse-like, the head long and narrow, and about 5 feet long, with two powerful tusks. The limbs were very long and stilted, enabling the animals to browse on the foliage of trees.
Of true rhinoceroses Scott says there are seven branches, three of which have living representatives. It appears that these striking animals had their origin in the North American genus Trigonias of the lower White River, a hornless form with four toes on the front feet instead of three as in all other rhinoceroses. Another hornless rhinoceros of the same time was CcenopuS, tridactyl on all feet, and this t3’pe persisted into the Pliocene. Conspicuous among the homed rhinoceroses were the small diceratheres (pair-homed), beginning in the Oligocene and persisting into the Miocene (Fig, p. 636). They also migrated into Eurasia, and are characterized by a transverse pair of horns on the nose. Pinally from the Old World there came to North America, in the Middle Miocene, Teleoceras, an aquatic type with a small horn on the nose, and grotesquely short legs, so that the belly almost touched the ground (Fig., above). This stock persisted in great numbers into the lower Pliocene.

In the Upper Miocene and Lower Pliocene of this continent there were at least four distinct types of rhinoceroses living in great [ p. 638 ] abundance, but none of them attained the size or bore the great horns of living species. Why they failed to live later, and why the woolly rhinoceros of Eurasia did not come to North America along with the mammoth is not known.
The hyracodonts are the ancestral hornless animals out of which all rhinoceroses came. The oldest one occurs in the Eocene (Bridger) and all of them died out in the Oligocene (White River). Hyrackyus (Fig., below), the oldest genus, is much generalized, and out of it might have also developed the horses and titanotheres. It was about as large as a sheep, but heavier. HjTacodonts were lightly built, with heavy clumsy heads, but slender and long necks and limbs; they are suggestive of horses rather than rhinoceroses. In fact, the feet of the White River forms resemble those of horses of the same time (Mesohippus). For safety they depended upon speed, and accordingly are called cursorial or running rhinoceroses. Scott says that it is interesting to reflect that had the White River hyracodonts and the ancestral tapirs continued to live up to the present, they would in the course of their evolution have developed one-toed feet as did the horses. In the hyracodonts, as in the horses, the evolution was mainly in the elongation of neck, limbs, and feet, and in the enlargement of the middle toes. With the increase in the size of their bodies in the course of their existence, the limbs became stouter, but all were of the cursorial or running type. In but one genus was a pair of small nasal horns developed (ColonocercLs).
The amynodonts were the specialized hornless aquatic rhinoceroses. They probably originated in Hyrachyus in late Bridger time and died out in the Oligocene. Amynodon was smaller and lighter than the later and larger Metamynodon of the White River. According to Scott, the latter was the heaviest and most massive creature of its time. Its head was large, depressed, and broad, [ p. 639 ] the nostrils high on the head in keeping with the aquatic habits, the neck short, the body long and massive, and the limbs short and stout, with four toes on the front feet and three on the hind ones.
¶ Collateral Reading
R. S. Lull, The Evolution of the Horse Family. American Journal of Science, 4th series, Vol. 23, 1907, pp. 161-182.
W. D. Matthew and S. H. Chubb, Evolution of the Horse. American Museum of Natural History", Guide Leaflet Series, No. 36, 1913.
H. F. Osborn, The Age of Mammals. New York (Macmillan’) , 1910.
H. F. Osborn, The Extinct Giant Rhinoceros Baluchitherium of Western and Central Asia. Natural Historj", Vol. 23, 1923, pp. 209-22S.
W. B. Scott, A History of Land Mammals in the Western Hemisphere. New York (Macmillan), 1913.
| XLII. The Evolution of Mammals and the Rise of Mentality in the Cenozoic | Title page | XLIV. The Evolution of the Elephants |