[ p. 49 ]
ANCESTRY OF THE ANTHROPOID APES — PLIOCENE CLIMATE, FORESTS, AND LIFE OF WESTERN EUROPE — TRANSITION TO THE PLEISTOCENE, OR AGE OF MAN — THE FIRST GLACIATION, ITS EFFECTS ON CLIMATE, FORESTS, AND ANIMAL LIFE — THE PREHUMAN TRINIL RACE OF JAVA — THE EOLITHS OR PRIMITIVE FLINTS — THE SECOND GLACIATION — THE HEIDELBERG, EARLIEST KNOWN HUMAN RACE — THE THIRD GLACIATION
The partly known ancestors of tke anthropoid apes and the unknown ancestors of man probably originated among the forests and flood-plains of southern Asia and early began to migrate westward into northern Africa and western Europe.
As early as Oligocene times a forerunner of the great apes (Propliopithecus), most nearly resembling the gibbons, appears in the desert bordering the Fayum in northern Egypt. Early in Miocene times true tree-living gibbons found their way into Europe and continued throughout the Pliocene in the forms known as Pliopithecus and Pliohylobates, the latter being a true gibbon in its proportions; it ranged northward into the present region of Germany. Another ape which early reached Europe is the Dryopithecus; it is found in Miocene times in southern France ; the grinding-teeth suggest those of the orang, the jaw is deep and in some ways resembles that of the Piltdown man. A third ape (Neopithecus) occurs in the Lower Pliocene near Eppelsheim, in Germany, and is known only from a single lower molar tooth, which recalls the dentition of Dryopithecus and more remotely that of Homo. In the Pliocene of the Siwalik hills of Asia is found Palaeopithecus, a generalized form which is believed to be related to the chimpanzee, the gorilla, and the gibbon ; the upper premolars resemble those of man.
None of these fossil anthropoids either of Europe or of Asia can be regarded as ancestral to man, although both Neopithecus [ p. 50 ] and Dryopithcciis have been placed in or near the line of human ancestry by such high authorities as Branco and Gaudry. When Dryopithecus was first discovered by Lartet, Gaudry1 considered it to be by far the most manlike of all the apes, even attributing to it sufficient intelligence for the working of flints, but fuller knowledge of this animal has shown that some of the living anthropoids are more manlike than Dryopithecus. This animal is closely related to the ancestral stock of the chimpanzee, gorilla, and orang. The jaw, it is true, resembles that of the Piltdown man (Eoanthropus), but the grinding-teeth are much more primitive and there is little reason to think that it is ancestral to any human type.[1]

[ p. 51 ]
Among these fossil anthropoids, as well as among the four living forms, we discover no evidence of direct relationship to man but very strong evidence of descent from the same ancestral stock. These proofs of common ancestry, which have already been observed in the existing races of man, become far more conspicuous in the ancient Palaeolithic races ; in fact, we cannot interpret the anatomy of the men of the Old Stone Age without a survey of the principal characters of the existing anthropoid apes, the gibbon, the orang, the chimpanzee, and the gorilla.

The gibbon is the most primitive of living apes in its skull and dentition, but the most specialized in the length of its arms and its other extreme adaptations to arboreal life. As in the other anthropoids, the face is abbreviated, the narial region is narrow, i. e., catarrhine, and the brain-case is widened, but the top of the skull is smooth, and the forehead lacks the prominent ridges above the orbits; thus the profile of the skull of [ p. 52 ] the gibbon (Fig. i6) is more human than that of the other anthropoid apes. When on the ground the gibbon walks erect and is thus afforded the free use of its arms and independent movements of its fingers. In the brain there is a striking development of the centres of sight, touch, and hearing. It is these characteristics of the modem gibbon which preserve with relalively slight changes the type of the original ancestor of man, as noted by Elliot Smith.2

The limbs of the orang are less elongated and less extremely specialized for arboreal life than those of the gibbon but more so than those of the chimpanzee and the gorilla. The skull is rounded and of great vertical height, with broad, bony ridges above the orbits and a great median crest on top of the skull in old males. The lower jaw of the orang is stout and deep, and, although used as a fighting weapon, the canine tusks are much [ p. 53 ] less prominent than in either the gibbon, chimpanzee, or gorilla.
In the chimpanzee we observe the very prominent bony ridges above the eyes, like those in the Trinil and Neanderthal races of men. Of all the anthropoid apes the lower jaw of the chimpanzee most nearly resembles that of the Piltdown man. The prognathous or protruding tooth rows and receding chin suggest those in the Heidelberg, Piltdown, and Neanderthal races. When the chimpanzee is walking (Fig. 18) the arms reach down below the level of the knees, whereas in the higher races of man they reach only half-way down the thighs.

[ p. 54 ]
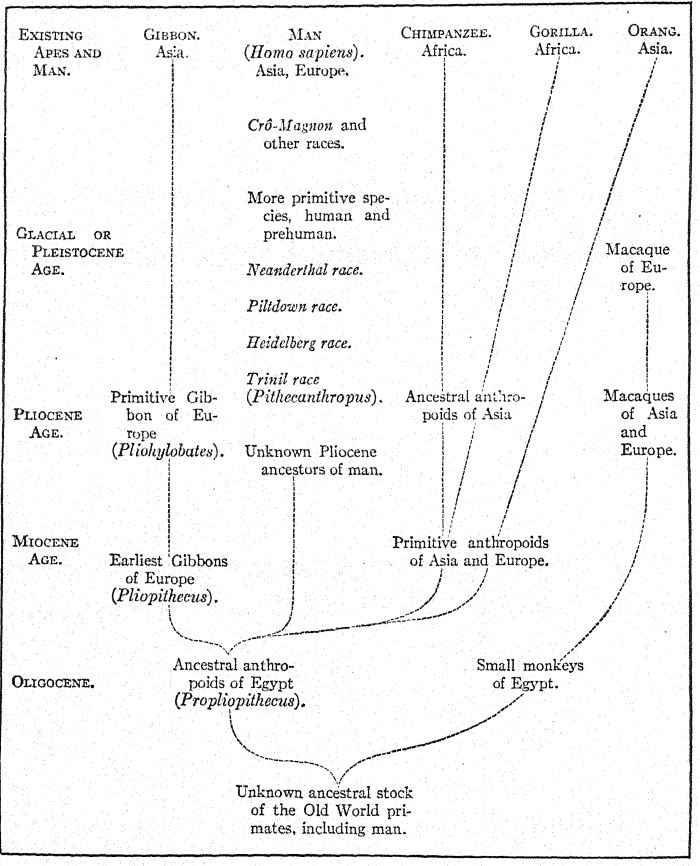
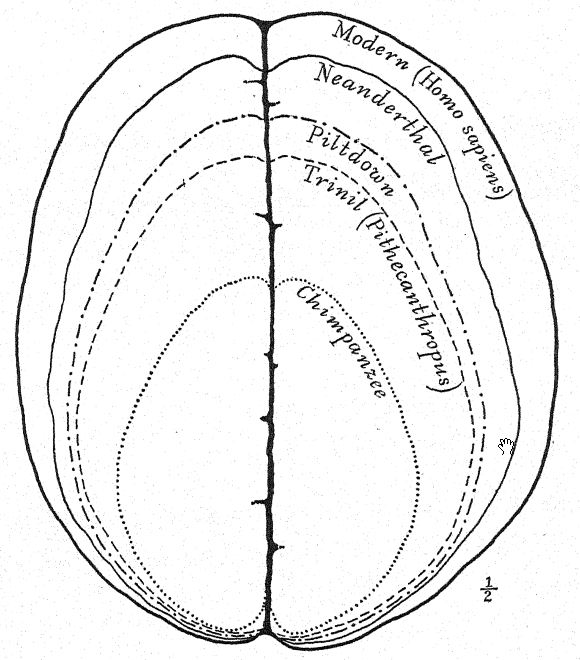
From the unknown and ancestral stock of the anthropoid apes and man the gibbon was the first to branch off in Oligocene times; the orang then branched oflf in a widely different direction. The stem of the chimpanzee and of the gorilla branched off at a more recent date and is more nearly allied to that of man. Five early human races have been found in Europe in Glacial or Pleistocene times, but no traces of other primates except the macaques, which are related to the lower division of the baboons, have been found in Europe in Pleistocene times. Modified after Gregory. (For latest discovery see Appendix, Note VII.)
Thus, the fore limb, although much shorter than that of the gibbon, is relatively longer than that of any human race, recent or ancient. We observe also in the walking chimpanzee (Fig. 18) [ p. 55 ] that the upper part of the leg, the thigh-bone, or femur, is relatively long, while the lower part, the shin-bone, or tibia, is relatively short. Indeed, both in the arm and in the leg the upper bones are relatively long and the lower bones are relatively short. These proportions, which are inheritances of arboreal life, are in very marked contrast to those observed in the arms and legs of the Neanderthal race of men, in which the limbs are of the terrestrial or walking type.

We observe also in the chimpanzee a contrast between the grasping power of the big toe, which is a kind of thumb, and the lack of that power in the hand, in which the thumb is nearly useless ; in all apes this function is characteristic of the foot, in man of the hand alone. The opposable thumb, with its power of bringing the thumb against each of the fingers, is the one character which is lacking in every one of the anthropoid apes and which was early developed arnong the ancestors of man.
The skull of the chimpanzee is longer than that of the orang, the most prominent feature in the top view being the extreme protuberance of the orbits, which are surrounded by a supraorbital [ p. 56 ] and cireumorbital bony ridge, wlrich is also strongly developed in the Neanderthal skull as well as in the Pithecanthropus or Trinil skull but, so far as we know, is entirely lacking in that of Piltdown. As in the orang and the gorilla, a crest develops along the middle of the top of the skull for the insertion of the powerful muscles of the jaws, a crest which is wholly wanting in the gibbon and probably wanting in all the true ancestors of man.
The gorilla illustrates in the extreme the specializations which are begun in the chimpanzee, and which are attributable to a life partly arboreal, partly terrestrial, with the skull and jaws used as powerful fighting organs. The head is lengthened by the forward growth of the muzzle into an extreme prognathism. The limbs and body of the gorilla show a departure from the primitive, slender-limbed, arboreal type of apes and are partly adapted to a bipedal, ground-dwelling habit.
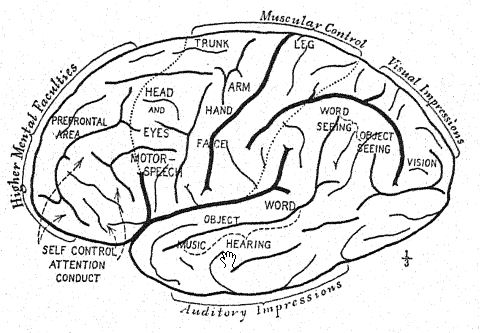
As regards psychic evolution,3 Elliot Smith observes that the arboreal mode of life of the early ancestors of man developed quick, alert, and agile movements which stimulated the progressive development of the posterior and lateral portions of the brain. The sense of smell had been well developed in a previous terrestrial life, but once these creatures left the earth and took [ p. 57 ] to the trees, guidance by the olfactory sense was less essential, for life amidst the branches of the trees is most favorable to the high development of the senses of vision, touch, and hearing. Moreover, it demands an agility and quickness of movement that necessitate efficient motor centres in the brain to co-ordinate and control such actions as tree life calls for. The specialization of sight awakens curiosity to examine objects with greater minuteness and guides the hands to more precise and skilled movements.
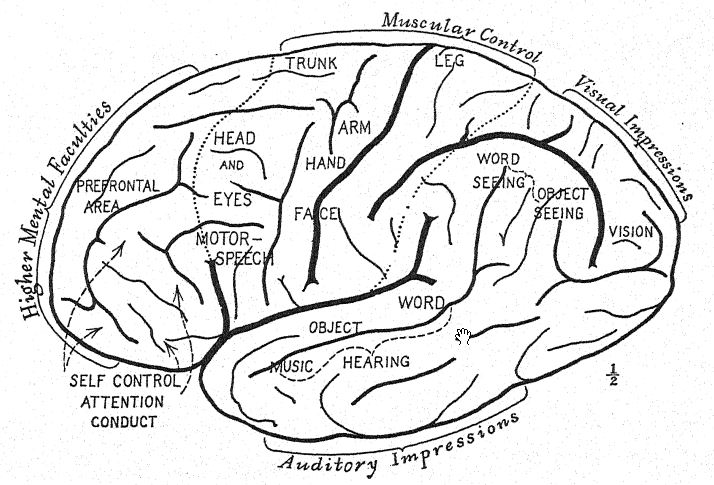
The anatomy of man is full of remote reminders of this original arboreal existence, which also explains the very large and early development of the posterior portions of the brain, in which the various senses of sight, touch, and hearing are located.
The first advance from arboreal to terrestrial life is marked by the power of walking more or less erect on the hind limbs and thus releasing the arms; this power is developed to a greater or less degree in all the anthropoid apes ; with practice they become [ p. 58 ] expert walkers. The additional freedom which the erect attitude gives to the arms and to the movements of the hands and the separate movements of the fingers is especially noticeable in the gibbon. The cultivation of the powers of the hand reacts upon the further growth and specialization of the brain ; thus the brain and the erect attitude react upon each other. In the gibbon there is a marked increase in the size of those portions of the brain which supply the centres of touch, vision, and hearing.
Discussion as to how the ancestors of man were fashioned has chiefly dealt with the rival claims of four lines of structural evolution: first, the assumption of the erect attitude; second, the development of the opposable thumb; third, the growth of the brain; and fourth, the acquisition of the power of speech. The argument for the erect attitude suggested by Lamarck, and ably put by Munro4 in 1893, indicates that the cultivation of skill [ p. 59 ] with the hands and fingers lies at the root of man’s mental supremacy. Elliot Smith’s argument that the steady growth and specialization of the brain itself has been the chief factor in leading the ancestors of man step by step upward indicates that such an advance as the erect attitude was brought about because the brain had made possible the skilled movements of the hands.
The true conception of prehuman evolution, which occurred during Miocene and Pliocene times, is rather that of the coincident development of these four distinctively human powers. It appears from the limb proportions in the Neanderthal race [ p. 60 ] that the partly erect attitude and walking gait were assumed much earlier in geologic time than we formerly imagined. The intimate relation between the use of the opposable thumb and the development of the higher mental faculties of man is sustained to-day by the discovery that one of the best methods of developing the mind of the child is to insist upon the constant use of the hands, for the action and reaction between hand and brain is found to develop the mind. A similar action and reaction between foot and brain developed the erect gait which released the hand from its locomotive and limb-grasping function, and by the resultant perfecting of the motion of thumbs and fingers turned the hand into an organ ready for the increasing specialization demanded by the manufacture of flint implements.
This is the stage reached, we believe, in late Pliocene times in which the human ancestor emerges from the age of mammals and enters the age of man, the period when the prehistory of man properly begins. The attitude is erect, the hand has a well-developed opposable thumb, the centres of the brain relating to the higher senses and to the control of all the motions of the limbs, hands, and fingers are well developed. The power of speech may still be rudimentary. The anterior centres of the brain for the storing of experience and the development of ideas are certainly very rudimentary.
¶ Change of Environment in Europe
Considering that the origin and development of any creature are best furthered by a struggle for existence sufficiently severe to demand the full and frequent exercise of its powers of mind and body, it is interesting to trace the sequence of natural events which prepared western Europe for the entrance of the earliest branches of the human race. The forests and plants portray even more vividly than the animals the changing conditions of the environment and temperature which marked the approach and various vicissitudes of the great Ice Age.
[ p. 61 ]
The forests of central France in Pliocene times, as well as those of the valley of the Arno in northern Italy, were very similar to the forests of the middle United States at the present time, comprising such trees as the sassafras, the locust, the honeylocust, the sumach, the bald cypress, and the tulip. Thus the regions which harbored the rich forest and meadow fauna of northern Italy in Upper Pliocene times abounded in trees familiar to-day in North and South Carolina, including even such distinctively American forms as the sweet gum (Liquidambar styraciflua), the sour gum (Nyssa sylvatica) , and the bay, beside those above mentioned. To the south, along the Mediterranean, there also flourished trees incident to a more tropical climate, the bamboo, the sabal palm, and the dwarf fan-palm ; most interesting is the presence of the sabal, which now flourishes in the subtropical rain forests of central Florida. The sequoia also was abundant. Toward the close of the Pliocene the first indications of the coming Glacial Epoch were a lowering of the temperature, and, in the higher mountainous areas perhaps, a beginning of the glacial stages.
The ancestors of the modern forests of Europe predominated in central France; the oak, the beech, the poplar, the willow, and the larch. It is these forests, which survived the vicissitudes of glacial times, that gave descent to the forests of Postglacial Europe, while all the purely American types disappeared from Europe and are now found only in the temperate regions of the United States.5
We have seen that few anthropoid apes have been discovered either in the Middle or Upper Pliocene of Europe ; the gibbon-ape line disappears with the Pliohylobates of the Upper Pliocene. These animals are, however, rarely found in fossil form, owing to their retreat to the trees in times of flood and danger, so that we need not necessarily assume that the anthropoids had actually become extinct in France. The primates which are found in the Upper Pliocene belong to the lower types of the Old World monkeys, related to the living langur of India and to the macaque and baboon. The evidence, as far as it goes, indicates that the [ p. 62 ] ancestors of man were at this time evolving in Asia and not in Europe. This evidence, nevertheless, would be completely offset if it could be proven that the eoliths, or primitive flints, found in various parts of Europe from Oligocene to Pleistocene times are really artifacts of human or prehuman origin.
The mammals of Europe in Pliocene times were derived by very remote migrations from North America and, more directly, from southern Asia. The Oriental element is very strong, including types of rhinoceroses now peculiar to Sumatra and southern Asia, numerous mastodons very similar to the south Asiatic types of the times, gazelles and antelopes, including types related to the existing elands, and primitive types of horses and of tapirs. Among the carnivores in Europe similar to south Asiatic species were the hysenas, the dog bears (Hyaenarctos), the civets, and the pandas (Ailurus) ; there were also the sabre-tooth tigers and numerous other felines. In the trees were found the south Asiatic and north African monkeys ; and in the forests the axis deer, now restricted to Asia. But the most distinctive African-Asiatic animal of this period was found in the rivers; namely, the hippopotamus, which arrived in Italy in the early Pliocene and ranged south by way of the Sicilian land bridge into northern Africa and east along the southern shores of the Black Sea to the Siwalik hills of India. Thus, many of the ancestors of what we have termed the African-Asiatic mammal group of Pleistocene times had already found their way into Europe early in Pliocene times. In middle and late Pliocene times there arrived three very important types of mammals which played a great rôle in the early Pleistocene. These are:
The true horses (Equus stenonis) of remote North American origin.
The first true cattle (Leptobos elatus), originating in southern Asia.
The true elephants, first Elephas planifrons and later E. meridionalis, better known as the southern mammoth, both originating in Asia.
[ p. 63 ]
The forests and river borders of the valley of the Arno, near Florence, contained all these African-Asiatic animals in Upper Pliocene times. Here they received their names which remind us of this region of Italy as it is today, such as the Etruscan rhinoceros (Dicerorhinus etruscus), the Florentine macaque (Macacus florentinus), Steno’s horse (Equus stenonis), the Etruscan cattle (Leptobos etruscus), which was the earliest ox to reach Europe.
In Italy and France these African-Asiatic mammals were mingled with ancestors of the more hardy Eurasia tic forest and meadow group. Of these the most graceful were a variety of deer with very elaborate or many-branched antlers, hence known as the ‘polycladine’ deer. In the forests roamed the wild boars of Auvergne (Sus arvernensis) , also the bears of Auvergne (Ursus arvernensis), lynxes, foxes, and wildcats. In the rivers swam the otter and the beaver, closely allied to existing forms. Among the rocks of the high hills were the pikas or tailless hares (Lagomys), also hamsters, moles, and shrews.
Many of the most characteristic animals of the dry modern plateaus of Africa had disappeared from Europe before the close of Pliocene times, namely, species of gazelles, antelopes, and the hipparion horses, all of which were adapted to the dry uplands or deserts of Africa. In the remaining faune Pliocene récente of French authors we find evidence that the Pliocene in all of western Europe closed with a moist, warm, temperate cHmate, with widespread forests and rivers interspersed with meadows favorable to the life of a great variety of browsing deer as well as of grazing elephants, horses and cattle. The flora of the Middle Pliocene as found at Meximieux indicates a mean annual temperature of 62° to 63° Fahr.
One of the proofs of the gradual lowering of temperature toward the close of Pliocene times in Europe is the southward retreat and disappearance of the apes and monkeys ; the Upper Miocene gibbon is found as far north as Eppelsheim, near Worms, Germany; in Lower Pliocene times the monkeys and apes are found only in the forests of the south of France; in Upper [ p. 64 ] Pliocene times they are recorded only in the forests of northern Italy; the evidence, so far as it goes, indicates a gradual retreat toward the south.
Finally, at the end of the Pliocene there existed very close geographic relations eastward with the mammalian life of India by way of what was then the isthmus of the Dardanelles and southward with the mammalian life of Africa by way of the Sicilian land bridge. This would indicate that the long lines of eastward and westward migration were open and favorable to the arrival in western Europe of new migrants from the far east, including perhaps the most primitive races of man. There is not the least evidence that Pliocene man or ancestors of man existed in Europe, excepting such as may be afforded by the problematic eoliths, or most primitive flints.
¶ The First Glaciation
In Upper Pliocene times cold marine currents6 from the north began to flow along the southeastern coast of England, with indications of a gradually lowering temperature culminating at a time when the sea abounded in the arctic mollusks, which have been preserved in the ‘Weyboum Crags,’ a geologic formation along the coast of Norfolk. This arctic current was the herald of the First Glacial Stage.
It does not appear that a glacial cap of any considerable extent was formed in Great Britain at this stage, but about this time the first great ice-cap was formed in British North America west of Hudson Bay, which sent its ice-sheets as far south as Iowa and Nebraska. In the latter State forests of spruce and other coniferous species indicate the appearance of a cool temperate flora in advance of the glaciation. In the Swiss Alps the snow descended 1,200 meters below the present snow-line, and in Scandinavia and northern Germany the first great ice-sheets were formed from which flowed the glaciers and rivers conveying the ‘Old Diluvium,’ or the ‘oldest drift.’ Accompanying the cold wave along the eastern coast of England we note, in the famous fossil deposits known as the ‘Forest Bed of Cromer,’ [ p. 65 ] which overlie the Weybourn Crags, the arrival from the north of the fir-tree (Abies). This is most significant, because it had hitherto been known only in the arctic region of Crinnell Land, and this was its first appearance in central Europe. Another herald of northern conditions was the first occurrence of the musk-ox in England, which is attributed7 to the ‘Forest Bed’ deposits.
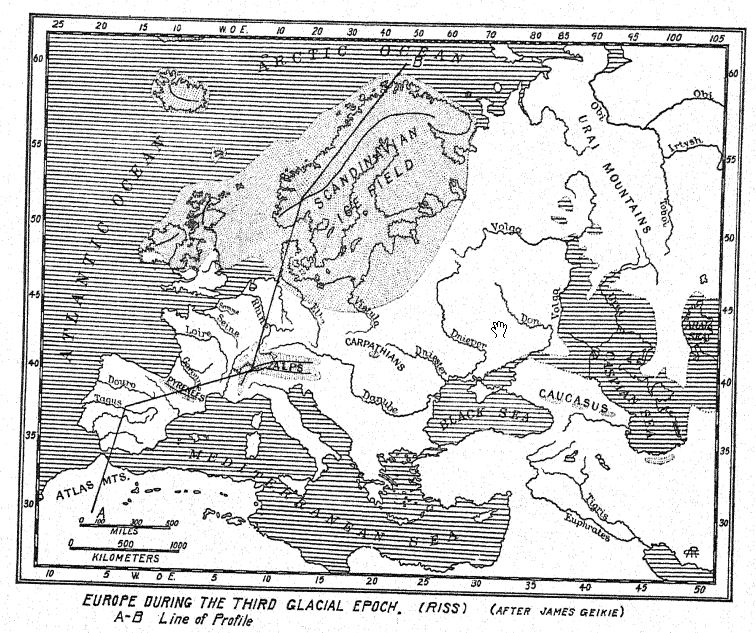
While Great Britain was less affected at this time than other regions, there is no doubt as to the vast extent of the First Glacial Stage in British America, in Scandinavia, and in the Alps ; in the latter region it has been termed ‘the Günz stage’ by Penck and Bruckner. The ‘drift’ deposits have a general thickness of 98½ [ p. 66 ] feet (30 m.), but they are largely covered and buried by those of the far more extensive Second Glacial Stage. The Scandinavian ice-sheet8 not only occupied the basin of the Baltic but overflowed Scania —the southern part of Sweden— and extended as far south as Hamburg and Berlin. In the Alps the glaciers passed down all the great mountain valleys to the low grounds of the foreland, implying a depression of the snow-line to 4,000 feet below its present level.

¶ The First Interglacial Stage. Eoliths
Proofs that a prolonged cool wave passed over Britain during the first glaciation are seen in its after effects, namely, in the modernization of the forests and in the disappearance both in Britain and France of a very considerable number of animals which were abundant in Upper Pliocene times. Yet by far the greater part of the Pliocene mammal life survived, a fact which tends to show that, while very cold conditions of climate and great precipitation of moisture may have characterized the regions immediately surrounding the ice-fields, the remainder of western Europe at most passed through a prolonged cool period during [ p. 67 ] the climax of the First Glacial Stage. This was followed during the First Interglacial by the return of a period somewhat warmer than the present.
This First Interglacial Stage is known as the Norfolkian, from the fact that it was first recognized in Europe in the deposits known as the ‘Forest Bed of Cromer,’ Norfolk, which contain rich records not only of the forests of the period, but of the noble forms of mammals which roamed over Great Britain and France in Norfolkian times. The forests of Norfolk, in latitude 52° 40 N. mainly abounded in trees still indigenous to this region, such as the maple, elm, birch, willow, alder, oak, beech, pine, and spruce, a forest flora closely corresponding to that of the Norfolk and Suffolk coasts of England at the present time, although we find in this fossil flora several exotic species which give it a slightly different character.9 From this tree flora Reid concludes that the climate of southeastern England was nearly the same as at present but slightly warmer.
We note especially that a very great change had taken place in the entire disappearance in these forests of the trees which in Pliocene times were common to Europe and America, as described above ; in other words, the flora of Europe was greatly impoverished during the first cold wave.
In southern France, as at the present time, the interglacial climatic conditions were milder, for we find numerous species of plants, which are now represented in the Caucasus, Persia, southern Italy, Portugal, and Japan. Thus the First Interglacial Stage, which was a relatively short one, enjoyed a temperature now belonging about 4° of latitude farther south.
This First Interglacial Stage is also known as the St.-Prestien, because among the many localities in France and Italy which preserve the plant and mammal life of the times that of St. Prest, in the Paris basin, is the most famous. Here in 1863 Desnoyers10 first reported the discovery of a number of mammal bones with incision lines upon them, which he considered to be the work of man. These deposits were regarded at the time as of Pliocene age, and this gave rise immediately to a wide-spread theory [ p. 68 ] of the appearance of man as early as the Pliocene. The human origin of the incisions discovered by Desnoyers has long been a matter of dispute and is now regarded as very improbable. Similar lines may be of animal origin, namely, marks left by claws or teeth, or due to accidental pressure of sharp cutting surfaces. However, we do not pretend to express an opinion of any value as to the cause of these incisions. Supposed confirmation of the evidence of Desnoyers of the existence of Pliocene man was the alleged finding by Abbott of several worked flints, two in situ, in the ‘Forest Bed of Cromer,’ Norfolk. Many years later in similar deposits at St. Prest were discovered the supposed ‘eoliths’ which have been referred to Étage Prestien by Rutot. The age of the St. Prest deposits is, therefore, a matter of the very highest interest and importance.

[ p. 69 ]
St. Prest is not Pliocene; it is rather the most ancient Pleistocene deposit in the basin of Paris,11 and these incised mammal bones probably date from the First Interglacial Stage. The bed which has yielded the incised bones and the rich series of fossils consists of coarse river sands and gravels, forming part of a ‘high terrace,’ 98½ feet (30 m.) above the present level of the river Eure. This, like other ‘high terraces,’ contains a characteristic First Interglacial fauna, including the southern mammoth (E. meridionalis) , and Steno’s horse (E. stenonis). We also find here other very characteristic early Pleistocene mammals, such as the Etruscan rhinoceros (D. etruscus), the giant hippopotamus of early Pleistocene times (H. major), the giant beaver of the early Pleistocene (Trogontherium), three forms of the common beaver (Castor), and one of the bison (Bison antiquus). This mammalian life of St. Prest is very similar to that of Norfolk, England ; to that of Malbattu in central France, Puy-de-Dome ; of Peyrolles, near the mouth of the Rhdne, in southern France ; of Solilhac near Puy ; of Durfort, Gard ; of Cajarc, Lot-et-Garonne ; and finally to that of the valley of the Arno, in northern Italy.
One reason why certain authors, such as Boule and Depéret, have placed this stage in the Upper Pliocene is that the mammals include so many surviving Pliocene forms, such as the sabre-tooth tigers (Machaerodus), the ‘polycladine’ deer with the elaborate antlers (C. sedgwicki), the Etruscan rhinoceros, and the primitive Steno’s horse. But we have recently discovered that, with the exception of the ‘polycladine’ deer, these mammals certainly survived in Europe as late as the Second Interglacial Stage, and there is said to be evidence that some even persisted into the Third Interglacial Stage.
It is, therefore, the extinction or disappearance from Europe of many of the animals very abundant even in late Pliocene times which marks this fauna as early Pleistocene. Anthropoid apes are no longer found; indeed, there is no evidence of the survival of any of the primates, except macaques, which survive in the Pyrenees to late Pleistocene times; the tapir has entirely disappeared from the forests of Europe; but the most significant [ p. 70 ] departure is that of the mastodon, which is believed to have lingered in north Africa and which certainly survived in America into very late Pleistocene times. The animal life of western Europe, like the plant life, has lost one part of its Pliocene aspect while retaining another part, both in its mammalian fauna and in its forest flora.
The living environment as a whole, moreover, takes on a novel aspect through the arrival, chiefly from the north, of the more hardy animals and plants which had been evolving for a very long period of time in the temperate forests and meadows of Eurasia to the northeast and northwest. From this Eurasiatic region came the stag, or red deer (Cervus elaphus), also the giant deer (Megaceros), and from the northerly swamps the broadheaded moose (Alces latifrons). The presence of members of the deer family (Cervidae) in great numbers and representing many different lines of descent is one of the most distinctive features of First Interglacial times. Beside the new northerly forms mentioned above, there was the roe-deer (Capreolus), which still survives in Europe, but there is no longer any record of the [ p. 71 ] beautiful axis deer (Axis), which has now retreated to southern Asia. The ‘polycladine’ deer, first observed in the valley of the Arno, is represented in First Interglacial times by Sedgwick’s deer (C. sedgwicki), in Norfolk, and by the species C. dicranius of northern Italy, "where there also occurs the deer of the Carnutes’ (C. carnutorum).

We observe that browsing, forest-living, and river-living types predominate. Among the forest-frequenting carnivores were the wolverene, the otter, two kinds of bear, the wolf, the fox, and the marten; another forest dweller was a wild boar, related to the existing Sus scrofa of Europe.
Thus in the very beginning of Pleistocene times the forests of Europe were full of a wild life very similar to that of prehistoric times, mingled with which was the Oriental element, the great elephants, rhinoceroses, and hippopotami connecting Europe wdth the far east. Among these eastern migrants in the early Pleistocene were two new arrivals, the primitive wild cattle (Bos primigenius), and the first of the bison (Bison priscus).
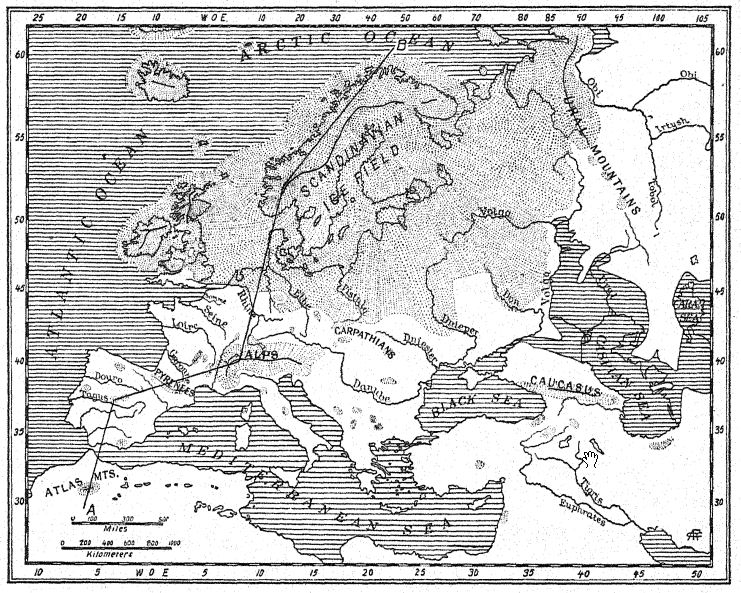
The theoretical map of western Europe during First Interglacial times (Fig. 12, also Fig. 56) enables us to understand these migrations from the northeast and from the Orient. As indicated by the sunken river channels discovered on the old continental shelf, the coast-line extended far to the west to the borders of the continental plateau which is now sunk deep beneath the ocean ; the British Isles were separated from France not by the sea but by a broad valley, while the Rhine, with the Thames as a western tributary flowed northward over an extensive floodplain, which is the present floor of the North Sea basin. It is not improbable that the rich mammalian life deposits in the ‘Forest Bed of Cromer,’ Norfolk, were washed down by tributaries of this ancient Rhine River.
In all the great rivers of this enlarged western Europe occurred the hippopotami, and along the river borders and in the forests browsed the Etruscan rhinoceros. Among the grazing and meadow-living forms of the Norfolk country of Britain were species of wild cattle (Bos, Leptohos), together with two species [ p. 72 ] of horses, including a lighter form resembling Steno’s horse (E. stenonis cocchi) of the Val d’Arno and a heavier type probably belonging to the forests. The giant elephant of this period is the southern mammoth (E. meridionalis trogontherii), a somewhat specialized descendant of the Pliocene southern mammoth of the valley of the Arno; this animal is best known from a superb specimen discovered at Durfort (Fig. 42) and preserved in the Paris Museum. It is said to have attained a height of over 12 feet as compared with 11 feet 3 inches, the height of the largest existing African elephants. It is probable that all these south Asiastic migrants into Europe were partially or wholly covered with hair, in adaptation to the warm, temperate climate of the summers and the cool winters. To the south, in the still milder climate of Italy, the arrival of another great species, known as the ‘ancient’ or ‘straight-tusked elephant’ (E. antiquus), is recorded. This animal had not yet reached France or Britain.
Preying upon the defenseless members of this heterogeneous fauna were the great machasrodonts, or sabre-tooth tigers, which ranged over Europe and northern Africa and into Asia. It does not appear that the true lions (Felis leo) had as yet entered Europe.
An intercommunication of life over a vast area extending 6,000 miles from the Thames valley on the west to India on the southeast is indicated by the presence of six or more similar or related species of elephants and rhinoceroses. Twenty-five hundred miles southeast of the foot-hills of the Himalayas similar herds of mammals, but in an earlier stage of evolution, roamed over the island of Java, which was then a part of the Asiatic mainland.
¶ The Trinil Race of Java
The human interest in this great life throng lies in the fact that the migration routes opened by these great races of animals may also have afforded a pathway for the earliest races of men. Thus the discovery of the Trinil race in central Java, amidst a [ p. 73 ] fauna closely related to that of the foot-hills of the Himalayas and more remotely related to that of southern Europe, has a more direct bearing upon our subject than would at first appear.
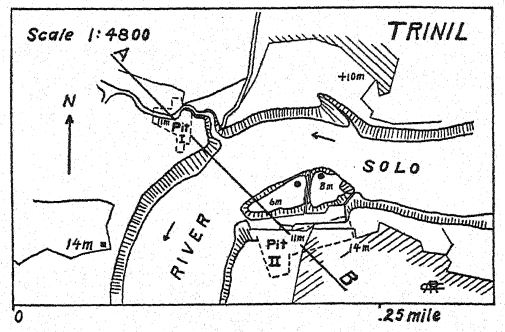
On the Bengawan River in central Java, a Dutch army surgeon, Eugen Dubois, had been excavating for fossils in the hope of finding prehuman remains. In the year 1891 he found near Trinil a deposit of numerous mammal bones, including a single upper molar tooth which he regarded as that of a new species of ape. On carefully clearing away the rock the top of a skull appeared at about a meter’s distance from the tooth. Further excavation at the close of the rainy season brought to light a second molar tooth and a left thigh-bone about 15 meters from the spot where the skull was found, imbedded and fossilized in the same manner. These scattered parts were described by Dubois13 in 1894 as the type of Pithecanthropus erectus,[2] a term signifying the [ p. 74 ] upright-standing ape-man. The specific term erectus refers to the thigh-bone, of which the author observes: “We must therefore conclude that the femur of Pithecanthropus was designed for the same mechanical functions as that of man. The two articulations and the mechanical axis correspond so exactly to the same parts in man that the law of perfect harmony between the form and function of a bone will necessitate the conclusion that this fossil creature had the same upright posture as man and likewise walked on two legs. … From this it necessarily follows that the creature had the free use of the upper extremities— now superfluous for walking— and that these last were no doubt already far advanced in that line of differentiation which developed them in mankind into tools and organs of touch. . . . From a study of the femur and skull it follows with certainty that this fossil cannot be classified as simian. . . . And, as with the skull, so also with the femur, the differences that separate Pithecanthropus from man are less than those distinguishing it from the highest anthropoid. … Although far advanced in the course of differentiation, this Pleistocene form had not yet attained to the human [ p. 75 ] type. Pithecanthropus erectus is the transition form between man and the anthropoids which the laws of evolution teach us must have existed. He is the ancestor of man.”


Thus the author placed Pithecanthropus in a new family, of the order Primates, which he named the Pithecanthropidae.
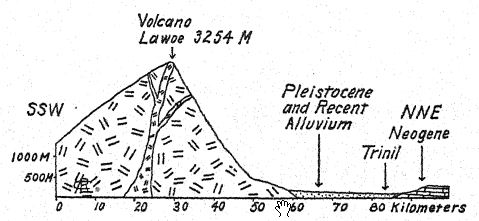
The geologic age of the bones referred to is a matter of first importance. The remains of Pithecanthropus lay in a deposit about one meter in thickness, consisting of loose, coarse, tufaceous sandstones, below this a stratum of hard, bluegray clay, and under that marine breccia. Above the Pithecanthropus layer were the ‘Kendeng’ strata, a many-layered tufaceous sandstone, about 15 meters in thickness. This geologic series was considered by Dubois and others to be of late Tertiary or Pliocene age ; Pithecanthropus accordingly became known as the longawaited ‘Pliocene ape-man.’ Subsequent researches by expert geologists have tended to refer the age to the early Pleistocene.17 According to Elbert18 the Kendeng strata overlaying the Pithecanthropus layer correspond to an early pluvial period of low temperature and, in point of time, to the [ p. 76 ] Ice Age of Europe. For even in Java one can distinguish three divisions of the Pleistocene period, including the first period of low temperature to which the Pithecanthropus layer is referred.
The fossil mammals contained in the Pithecanthropus layer have also been thoroughly studied,19 and they tend to confirm the original reference to the uppermost Pliocene. They yield a very rich fauna similar to that of the Siwalik hills of India, ineluding the porcupine, pangolin, several felines, the hyaena, and the otter. Among the primates beside Pithecanthropus there is a macaque. Among the larger ungulates are two species of rhinoceros related to existing Indian forms, the tapir, the boar, the hippopotamus, the axis and rusa deer, the Indian buffalo, and wild cattle. It is noteworthy that three species of late Pliocene elephants, all known as Stegodon, and especially the species Stegodon ganeza, occur, as well as Elephas hysudricus, a species related to E. antiquus, or the straight-tusked elephant, which entered Europe in early Pleistocene times. Fossils of the same animals are found in the foot-hills of the Himalayas of India, about 2,500 miles distant to the northwest. The India deposits are considered of uppermost Pliocene age,20 for this is the closing life period of the upper Siwaliks of India.
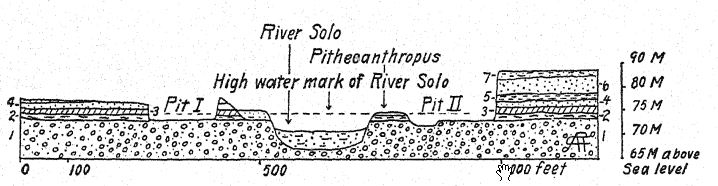
Recent 7 River wash, blue-black clay.
Pleistocene: 6 Light-colored sandstone, like tuff.
5 Gray tuff with balls of clay, fresh-water shells.
4 White streaked sandstone resembling tufa.
3 Blue-black clay with plant remains.
2 Bone-bearing stratum. Pithecanthropus.
I Lahar conglomerate.
[ p. 77 ]

Certainly Java was then a part of the Asiatic continent, and similar herds of great mammals roamed freely over the plains from the foot-hills of the Himalaya Mountains to the borders of the ancient Trinil River, while similar apes inhabited the forests. At this time the orang may have entered the forests of Borneo, which are at present its home ; it is the only ape thus far found in the uppermost Pliocene of India. We may, therefore, anticipate the discovery, at any time, in India of a race similar to Pithecanthropus.
The geologic age of the Trinil race is, therefore, to be considered as late Plio- cene or early Pleistocene.
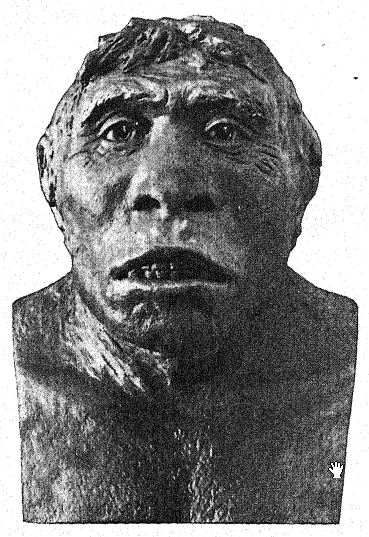
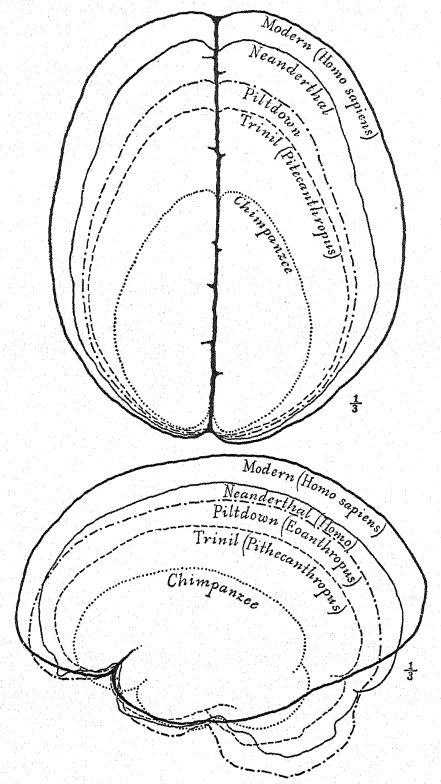
This great discovery of Dubois aroused wide-spread and heated discussion, in which the foremost anato- mists and palteontologists of the world took part. Some regarded the skull as that of a giant gibbon, others as prehuman, and Still others as a transition form. We may form our own opinion, however, from a fuUer understanding of the specimens themselves, always keeping in mind that it is a question whether the femur and the skull belong to the same individual or even to the same race. First, we are struck by the marked resemblance which the top of the skull bears, both on viewing it from the side and from above, to that of the Neanderthal race. This fully justifies the opinion of the anatomist Schwalbe21 that the skull of Pithecanthropus is nearer to that of Neanderthal man than to that of even the [ p. 78 ] highest of the anthropoid apes. As measured by Schwalbe, the index of the height of the cranium (Kalottenhöheindex) may be compared with others as follows :
| Lowest human race | 52 per cent. |
| Neanderthal man | 40.4 per cent. |
| Pithecanthropus, or Trinil race | 34.2 per cent. |
This accords with the estimate of the brain capacity[3] of 855 c. cm. (Dubois) as compared with 1,230 c. cm., the smallest brain capacity found in a member of the Neanderthal race. Second, as seen from above, we are struck with the great length of the calvarium as compared with its breadth, the cephalic index or ratio of breadth to length being 73.4 per cent (Schwalbe) as compared with 73.9 per cent in the Neanderthal type skull ; this dolichocephaly accords with the fact that all of the earliest human races thus far found are long-headed, although according to Schwalbe22 all anthropoids are broad-headed. This is a very important distinction. The third feature is the prominence and width of the bony eyebrow ridges above the orbits, which are almost as great as in the chimpanzee and greatly exceed those [ p. 79 ] of the Neanderthal race and of the modern Australian. The profile of the Trinil head restored by McGregor (Fig. 38) exhibits this prominent bony ridge and the low, retreating forehead. In the latest opinion of Schwalbe23 Pithecanthropus may be regarded as one of the direct ancestors of Neanderthal man and even of the highest human species, Homo sapiens. He also considers that when the lower jaw of the Trinil race becomes known, it will be found to be very similar to that of the Heidelberg man, the final conclusion being that Pithecanthropus and the nearly allied Heidelberg man may be regarded as the common ancestors of the Neanderthal race, on the one hand, and of the higher races on the other. There are, however, reasons for excluding Pithecanthropus from the direct ancestral line of the higher races of man.

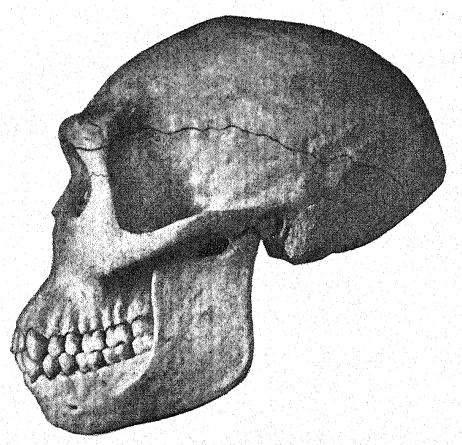
This prehuman stage has, none the less, a very great significance in the developmental history of man. In our opinion it is the very stage which, theoretically, we should anticipate finding in the dawn of the Pleistocene. A similar -view is taken by Buchner, who presents in an admirable diagram (Fig. 117) the [ p. 80 ] result of his comparison of twelve different characters in the skulls of Pithecanthropus, the Neanderthals, the Australians, and the Tasmanians. One of the main objects of Buchner’s research was a very detailed comparison of the Trinil skull with that of the lowly and now extinct Tasmanian race, which, we observe in the diagram, occupies a position only a little higher than that of the Spy-Neanderthal race.
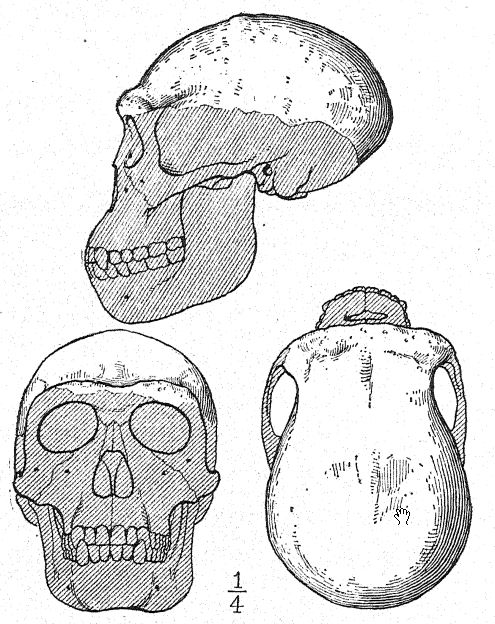
If the femur belongs with the skull, the Trinils were a tall race, reaching a height of 5 feet 7 inches as compared with 5 feet 3 inches in the Neanderthals. The thigh-bone (Fig. 122) has a very slight curvature as compa,red with that of any of the apes or lemurs, and in this respect is more human; it is remarkably elongate (455. mm.), surpassing that of the Neanderthals; the [ p. 81 ] shin-bone (tibia) was probably correspondingly short. The two upper grinding-teeth preserved are much more human than those of the gibbon, but they do not resemble those of man closely enough to positively confirm the prehuman theory. Dubois observes:25 “That the tooth belongs to some hominid form needs no further demonstration. Aside from its size and the greater roughness of the grinding surface, it differs from the human grinder in that the less developed cusp of Pithecanthropus is the posterior cusp next the cheek, while in man it is generally the posterior cusp next the tongue. The simplification of the crown and the root of the Trinil grinder is quite as extensive as it usually is in man.”
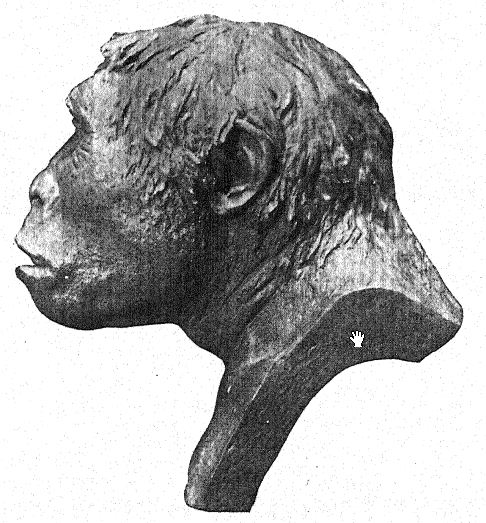
Various efforts have been made to supplement the scattered and scanty materials collected by Dubois. The Selenka expedition of 1907-8 brought back a human left lower molar as the only result of an express search for more Pithecanthropus remains. [ p. 82 ] Dubois is also said to possess the fragment of a primitive-looking lower jaw from the range knovni as the Kendeng Hills, at the southern base of which lies the village of Trinil.
It remains for us to consider the stage of psychic evolution attained by the Trinil race, and this naturally turns upon the erect attitude and what little is known of the size and proportions of the brain.
The assumption of the erect attitude is not merely a question of learning to balance the body on the hinder extremities.26 It involves changes in the interior of the body, the loss of the tail, the freeing of the arms, and the establishment of the diaphragm as the chief muscle of respiration. The thigh-bone of Pithecanthropus is so much like that of man as to support the theory that the erect position may have been assumed by the ancestors of man as early as Oligocene times. It would appear that Pithecanthropus had free use of the arms and it is possible that the [ p. 83 ] control of the thumb and fingers had been cultivated, perhaps in the fashioning of primitive implements of wood and stone.
The discovery of the use of wood as an implement and weapon probably preceded that of the use of stone.
Elliot Smith describes this stage of development as follows:27 “. . . The emancipation of the hands from progression threw the whole responsibility upon the legs, which became more efiicient for their purpose as supports-once they lost their prehensile powers and became elongated and specialized for rapid progression. Thus the erect attitude became stereotyped and fixed and the limbs specialized, and these upright simians emerged from their ancestral forests in societies, armed with sticks and stones and with the rudiments of all the powers that eventually enabled them to conquer the world. The greater exposure to danger which these more adventurous spirits encountered once they emerged in the open, and the constant struggles these first semihuman creatures must have had in encounters with definite enemies, no less than with the forces of Nature, provided the factors which rapidly weeded out those unfitted for the new conditions and by natural selection made real men of the survivors.”
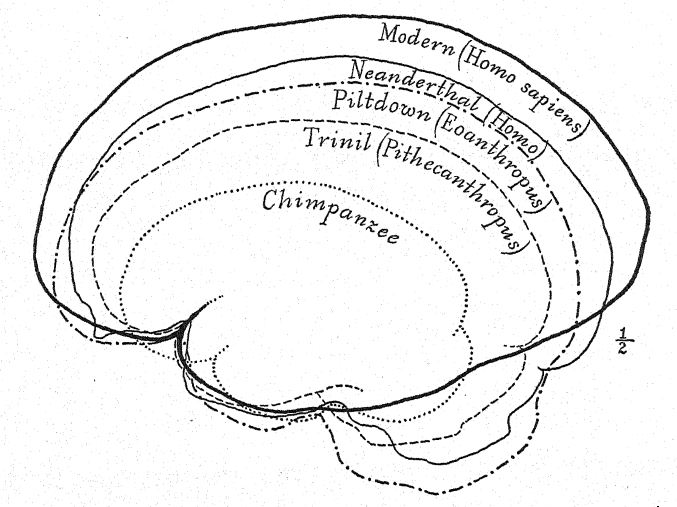
The undeveloped forehead of Pithecanthropus and the diminutive frontal area of the brain indicate that the Trinil race had a limited faculty of profiting by experienee and accumulated tradition, for in this prefrontal area of the brain are located the powers of attention and of control of the activities of all other [ p. 84 ] parts of the brain. In the brain of the ape the sensory areas of touch, taste, and vision predominate, and these are well developed in Pithecanthropus. The central area of the brain, which is the storehouse of the memories of actions and of the feelings associated with them, is also well developed, but the prefrontal area, which is the seat of the faculty of profiting by experience or of recalling the consequences of previous responses to experience, is developed to a very limited degree.28 Thus, while the brain of Pithecanthropus is estimated at 855-900 c. cm., as compared with 600 c. cm. of the largest simian brain, and 930 c. cm. of the smallest brain recorded in the human race, it indicates a very low stage of intelligence.
¶ Absence of Palaeoliths and Presence of Eoliths in Western Europe
Returning to First Interglacial conditions in Europe, we observe that the river courses flowed through the same valleys as at present but that in early glacial times the channels were far [ p. 85 ] broader and were elevated from loo to 150 feet above the present relatively narrow river levels. The vast floods of the succeeding glaciation filled these valleys, but some of the ‘high terraces’ were already formed. It is extremely important to note that Pre-Chellean flints or true palaeoliths have never been found in the sands or gravels of these ‘high terraces.’
Eoliths found on this ‘high-terrace’ level at St. Prest belong to the Prestien culture of Rutot,29 who regards this station as of Upper Pliocene age. These, like other supposed Eolithic flints, are very rough, but, rude as they are, they generally exhibit one part shaped as if to be grasped by the hand, while the other part is edged or pointed as for cutting. It is generally admitted that these flints are mostly of accidental shapes, and there has been little or no proof of their being fashioned by human hands. On this point Boule30 observes : “ As to the eoliths, I have combated the theory not only because it seems to me improbable but because a long geological experience has shown me that it is often impossible to distinguish stones split, cut, or retouched by purely physical agents from certain products of rudimentary workmanship.”
On the other side, it is interesting at this point to quote the words of MacCurdy;31 “My opinion, based on personal experience, … is that the existence of a primitive industry, antedating what is commonly accepted as Palaeolithic, has been established. This industry occurs as far back as the Upper Miocene and continues on through the Upper Tertiary into and including the Lower Quaternary. The distinguishing characters of the industry remain but little changed throughout the entire period, the subdivision of the period into epochs being based on stratigraphy [geologic stages] and not on industrial characters. The requirements in the way of tools being very simple and the supply of material in the way of natural flakes and fragments of flint being very plentiful, the inventive powers of the population remained dormant for ages. Hammer and knife were the original tools. Both were picked up ready-made. A sharp-edged, natural flake served for one, and a nodule or fragment served for the other. When the edge of the flake became dulled by use, the [ p. 86 ] piece was either thrown away or the edge was retouched for further use. If hammer or flake did not admit of being held comfortably in the hand, the troublesome points or edges were removed or reduced by chipping. The stock of tools increased slowly with the slowly growing needs. As these multiplied and the natural supply of raw material diminished, the latter was supplemented by the manufacture of artificial flakes. When the lesson of associating definite forms of implements with definite uses was learned, special types arose, notably the amygdaloid implement and the poniard. Then came the transition from the Eolithic to the Palaeolithic, a stage that has been so thoroughly investigated by Rutot.”
It is not improbable that the Trinil race was in a stage of Eolithic culture ; it is highly probable that the prehuman races of this very remote geologic age used more than one weapon of wood and stone.
¶ The Great Second Glaciation (Fig. 25, p. 65)
In early Pleistocene times a general elevation of southern Europe united the islands of the Mediterranean with Europe on the north and with Africa on the south, forming broad land connections between the two continents which afforded both northward and southward migration routes. At this time certain characteristically African mammals, such as the straight-tusked elephant and the lion, were probably finding their way north; Sicily at this time gained its large fauna of elephants and hippopotami, and the island of Malta was connected with the mainland, as well as the easterly islands of Cyprus and Crete. It appears probable that the connection between the Italian mainland and Malta was renewed more than once.
The approach of the second glaciation is indicated along the southeast coast of Great Britain by the subsidence of the land and the rise of the sea, accompanied by a fresh arctic current, bringing with it an invasion of arctic moUusks which were deposited in a layer of marine beds directly over those which contain the [ p. 87 ] [ p. 88 ] [ p. 89 ] rich warm fauna and flora of the ‘Forest Bed of Cromer,’ Norfolk.®^ It also appears prdbable that a cold northern current swept along the western coasts of Europe, and Geikie estimates that a lowering of temperature occurred of not less than 20° Fahr., a change as great as is now experienced in passing from the south of England to the North Cape.

The second glaciation was by far the greatest both in Europe and America. In the region of the Pyrenees, which at the very much later period of the Third Interglacial Stage became a favorite country with Palaeolithic man, there were glaciers of vast extent. This is reahzed by comparison with present conditions. The largest of the present glaciers of the Pyrenees is only 2 miles in length and terminates at a height of 7,200 feet above the sea. During the greatest glaciation the snow appears to have descended 4,265 feet below its present level. From the Pyrenees through the Gallego valley into Spain there flowed a glacier 38 miles in length, while to the north the glacier in the valley of the Garonne flowed for a distance of 45 miles to a point near Montrejeau. Even in its lower reaches this glacier was over half a mile in thickness. To the east was a glacier 38 miles in length, filling the valley of the Ariege and covering the sites of such great Palaeolithic caverns as that of Niaux ; it is probable that at this time the formation of this cavern began. That these glaciers were all prior to the period of the Lower Palaeolithic Acheulean culture is proven by the fact that Acheulean implements are frequently met with lying on the surface of the moraines laid down by these ancient ice-floes.33
To the north was the vast Scandinavian ice-field, which swept over Great Britain and beyond the valleys of the Rhine, Elbe, and Vistula, reaching nearly to the Carpathians. Even the lesser mountain chains were capped with glaciers, including the Atlas Mountains in northern Africa.
In North xAmerica from the great centre west of Hudson Bay the ice-cap extended its drift southward into Missouri, Iowa, Kansas, and Nebraska, beyond the limits of earlier and subsequent glaciations.
[ p. 90 ]
The materials of the chief ‘high terraces’ of the great rivervalleys of western Europe were deposited at this time.
¶ Life of the Warm Second Interglacial Stage
The long warm period which followed the great glaciation is remarkable in presenting the first proofs of the presence of man in western Europe. It is the period of the Heidelberg race of man (Homo heidelbergensis) , known only from a single jaw discovered by Schoetensack in the Mauer sands near Heidelberg, in 1907. No other proofs of the existence of man have been found in any of the deposits which took place during this vast interval of geologic time, unless we accept the theory of Penck and of Geikie that the Pre-Chellean and Chellean quarries of the River Somme belong in the Second Interglacial Stage.
The vast duration of this interglacial time is evidenced both in Europe and America by the deep cutting and wearing away of the ‘drifts’ brought down by the second glaciation. Penck believes that this ‘long warm stage’ represents a greater period of time than the entire interval between the third glaciation and the present time. The climate immediately following the retreat of the glaciers was cool and moist in the glaciated regions, but this was followed by such a prolonged period of heat and dryness that the glaciers on the Alps withdrew to a point far above their present limits.
In one of the old ‘high terraces’ of the River Inn, in the north Tyrol, is a deposit containing the prevailing forest flora of the period, from which Penck concludes that the chmate of Innsbruck was 2° C. higher than it is at the present time. Corresponding with this the snow-line stood 1,000 feet above its present level, and the Alps, save for the higher peaks, were almost completely denuded of ice and snow. A characteristic plant is the Pontic alpine rose (Rhododendron ponticum), which, flourishes now in an aimual temperature of 57°-65° Fahr.,34 indicating that the climate of Innsbruck was as genial as that of the Italian slopes of the Alps to-day. This rhododendron is now found in the Caucasus. Other southern species of the time were a buckthorn, [ p. 91 ] related to a species now living in the Canary Islands, and the box. There were also more hardy plants, including the fir (Pinus sylvestris), spruce, maple, willow, yew, elm, beech, and mountain-ash. The forests of the same period in Provence were, for the most part, similar to those now found in that region ; out of thirty-seven species twenty-nine still occur in this part of southern France. On the whole, the aspect of southern France at this time was surprisingly modern. The forests included oaks, elms, poplars, willows, lindens, maples, sumachs, dogwood, and hawthorn. Among the climbing plants were the vine and the clematis. Here also were some forms which have since retreated to the south, such as species of the sweet bay and laurel which are now confined to the Canary Islands. The great humidity of the time is indicated by the presence of certain species of conifers which require considerable moisture. As in First Interglacial times, the presence of the fig indicates mild winters.
It is difficult to imagine forests of this modern character, which farther northward included a number of still more temperate and hardy species, as the setting of the great African and Asiatic life that roamed all over western Europe at this time. It was the presence of hippopotami, elephants, and rhinoceroses which gave to Lyell, Evans, and other early observers the impression that a tropical temperature and vegetation were characteristic of this long life period. These animals were formerly regarded as proofs of an almost tropical climate, but the more trustworthy evidence of the forests, strengthened by that of the presence of very numerous hardy types of forest and meadow animals, has set aside all the early theories as to extremely warm temperatures during Second Interglacial times.
The remains of what is still conveniently known as the ‘faune chaude,’ or warm fauna, are chiefly found in the sands and gravels of the ancient beds of the Neckar, Garonne, and Thames, and other rivers of the north and south, also in Essex, England. The most surprising fact is that the mammal life of western Europe remained entirely unchanged by the vast second glaciation just described ; the few extinctions which occurred as well as a number [ p. 92 ] of new arrivals may be attributed to new geographical connections with Africa on the south and to the steady progress of migration from the far east.
There were four very important and distinctive new arrivals from the African- Asiatic world, namely, the straight-tusked or ancient elephant (E. antiquus), the broad-nosed rhinoceros (D. merckii), the African lion (Felis leo), and the African hycena (H. striata), which bespeak close geographical connections with [ p. 93 ] northern Africa. Of these the ancient elephant and the broadnosed rhinoceros were close companions; they enjoyed the same regions and the same temperatures, their remains are very frequently found together, and they survived to the very end of the great life stage of western Europe, which closed with the advent of the fourth glaciation. They are in contrast to the other pair of great mammals which was already present in Europe in Pliocene and First Interglacial times, namely, the southern mammoth, at this stage known as trogontJierii, which had a preference for the companionship of the hippopotamus (H. major) ; it would seem that these animals Were less hardy because both [ p. 94 ] disappeared from Europe a little earlier than the ancient elephant and Merck’s rhinoceros.
The African lion would appear to have been a competitor of the sabre-tooth tiger, for the latter animal now becomes less abundant, although there is reason to believe that it survived until the Third Interglacial Stage. With the. ancient Pliocene type of the sabre-tooth were also foimd the Etruscan rhinoceros, the primitive bear of Auvergne (Ursus arvernensis), ojidi the giant he&Yev (Trogontherium cuvieri).
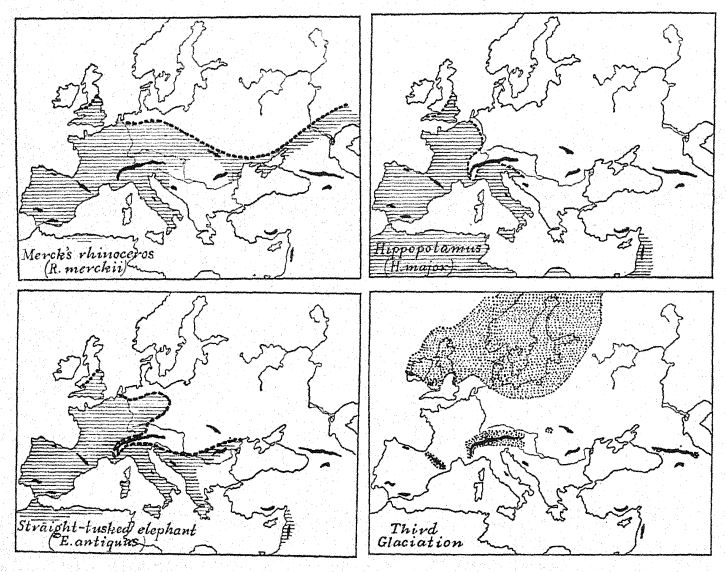
The northern forests of the time were frequented by the broad-faced moose, the giant deer, and the roe-deer, as well as by noble specimens of the stag (Cervus elaphus). In the open forests and meadows the wild cattle (Bos primigenius) began to be more [ p. 95 ] numerous and the bison (Bison priscus) also occurred. Among the meadow or forest frequenting forms were horses of larger size, such as the horses of Mosbach and of Süssenborn. In this assemblage of northern and southern types it is noteworthy that the Eurasiatic forest and meadow types of mammals greatly predominate in numbers and in variety over the xAfrican- Asiatic types; this, together with the flora, is an indication that the climate was of a temperate character ; it is probable, therefore, that all the mammals were well protected with a hairy covering and adapted to a temperate climate. The fact that the fauna as a whole remained practically unchanged throughout the second glaciation is a proof not that it migrated to the south and then returned but that the non-glaciated regions of western Europe were temperate rather than cold.
¶ The Heidelberg Race
Heidelberg man. Ancient elephant. Etruscan rhinoceros. Mosbach horse. Wild boar. Broad-faced moose. Red deer, or stag. Roe-deer. Primitive bison (wisent). Primitive o.v (Aurochs, urus). Auvergne bear. Deninger’s bear. Lion. Wildcat. Wolf. Beaver.
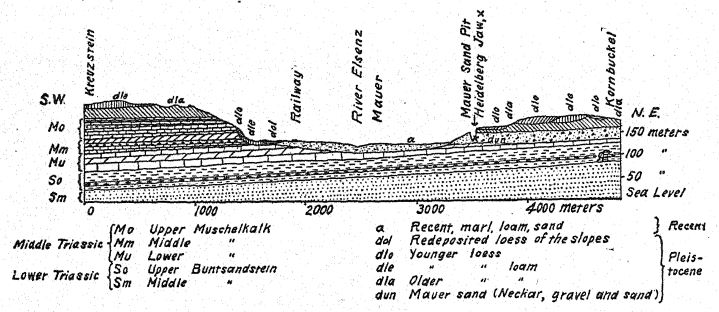
To us by far the most interesting mammalian life is that found south of the mouth of the Neckar along the ancient stream Elsenz, where were deposited the lower ‘sands of Mauer,’ containing the lower jaw of the Heidelberg man and the remains of many animals of the period. The enumeration of this entire fauna is very important, as indicating the temperate climatic conditions which surrounded the first true species of man which has thus far been discovered in Europe. The discoverer, Schoetensack,35 referred these mammals and the Heidelberg man to the First Interglacial Stage, and a similar opinion has recently been expressed by Geikie. The presence of the Etruscan rhinoceros would appear to point to such great antiquity, but the evidence afforded by this primitive animal is overborne by that of three mammals which are highly characteristic of Second Interglacial times ; these are the straight-tusked or [ p. 96 ] ancient elephant (E. antiquus), the lion, and the Mosbach horse. Excepting only the Etruscan rhinoceros, all these species frequenting the ancient stream Elsenz and deposited with the ‘sands of Mauer’ occurred also in the forests and meadows of the region now known as Baden, where the fossil mammal deposits of Mosbach near the Neckar are found. A similar mammalian life of a somewhat more recent time occurs in the river gravels of Süssenborn, near Weimar. The horses of Mauer, of Mosbach, and of Süssenborn[4] were of much larger size and of more specialized character than Steno’s horse of First Interglacial times.
Thus the Heidelbergs, the first human race recorded in western Europe, appear in southern Germany early in Second Interglacial times, in the midst of a most imposing mammalian fauna of northern aspect and containing many forest-living species, such as bear, deer, and moose; in the meadows and forests browsed the giant, straight-tusked elephant (E. antiquus) , from the simple structure of its grinding-teeth is regarded as similar in habit to the African elephant now inhabiting the forests of central Africa; the presence of this animal indicates a relatively moist climate and well-forested country. The Etruscan [ p. 97 ] rhinoceros differed from the larger Merck’s form in the possession of relatively short-crowned grinding-teeth, adapted to browsing habits and a forested country ; on the head were borne two horns ; it was a long-limbed, rapidly moving type ; the herds [ p. 98 ] of bison and of wild cattle (urus) which roamed over the plains were now subject to the attack of the lion.

a-b. ‘Newer loess,’ cither of Third Interglacial or of Postglacial times.
b-c. ‘Older loess’ (sandy loess) of the close of Second Interglacial times.
c-f. The ‘sands of Mauer.’
d-e. An intermediate layer of clay.
The white cross (X) indicates the spot at the base of the ‘sands of Mauer’ at which the jaw of Heidelberg was discovered.
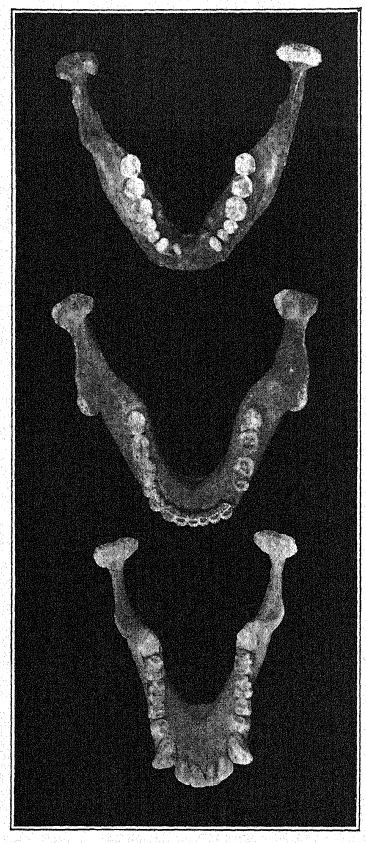
The discovery in 1907 of a human lower jaw in the base of the ‘Mauer sands’ is one of the most important in the whole history of anthropology. The find was made at a depth of 79 feet (24.10 m.) from the upper surface of a high bluff (Fig. 46), in ancient river sands which had long been known to yield the very old mammalian fauna described above. For years the workmen had been instructed to keep a sharp lookout for human remains. The jaw had evidently drifted down with the river sands and had become separated from the skull, but it remained in perfect preservation. The author’s description may first be quoted.36 The mandible shows a combination of features never before found in any fossil or recent man. The protrusion of the lower jaw just below the front teeth which gives shape to the human chin is entirely lacking. Had the teeth been absent it would have been impossible to diagnose it as human. From a fragment of the symphysis of the jaw it might well have been classed as some gorilla-like anthropoid, while the ascending ramus resembles that of some large variety of gibbon. The absolute certainty that these remains are human is based on the form of the teeth — molars, premolars, canines, and incisors are all essentially [ p. 99 ] human and, although somewhat primitive in form, show no trace of being intermediate between man and the anthropoid apes but rather of being derived from some older common ancestor. The teeth, however, are somewhat small for the jaw; the size of the border would allow for the development of much larger teeth ; we can only conclude that no great strain was put on the teeth, and therefore the powerful development of the bones of the jaw was not designed for their benefit. The conclusion is that the jaw, regarded as unquestionably human from the nature of the teeth, ranks not far from the point of separation between man and the anthropoid apes. In comparison with the jaws of Neanderthal races, as found at Spy, in Belgium, and at Krapina, in Croatia, we may consider the Heidelberg jaw as pre-Neanderthaloid; it is, in fact, a generalized type.
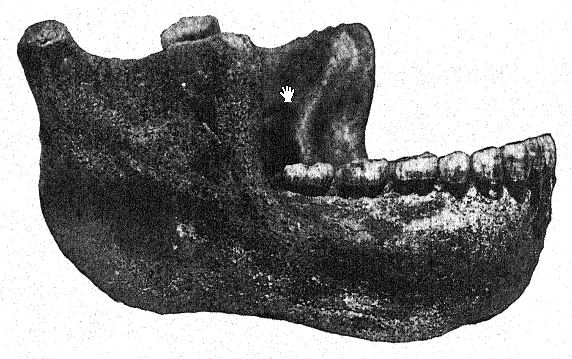
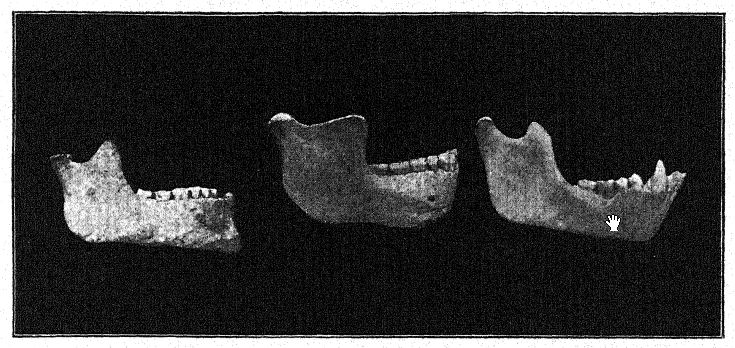
In a conservative spirit, Schoetensack named the type represented by this jaw Homo heidelbergensis. Other authors have regarded it as of distinct generic rank ; thus it has been termed Palaoanthropus heidelbergensis by Bonarelli.37 The jaw itself is extremely massive; the canine teeth, unlike those of the anthropoid apes and of the Piltdown race, do not project beyond the line of the other teeth and were therefore not used as weapons of offense and defense as in the anthropoids, in which these teeth [ p. 100 ] are prominently developed as tusks. As noted by Schoetensack, the teeth are not very massive in proportion to the jaw itself, which is the most powerful human jaw known, even exceeding the largest Eskimo jaw and indicating a skull of very massive and primitive character. It resembles that of the ape in the recession of the chin, hence it has been termed amentalis. There is a large development of the coronoid process of the mandible for the attachment of the temporal muscle. This jaw may well have been used as a tool in the last stages of the preparation of hides, as is the practice of the Eskimo races. We observe that the powerful bony branches of the jaw, when regarded from above, close in upon the space left for the tongue; in fact, the bone closes in to such an extent as to interfere seriously with the free use of the tongue in articulate speech.
It would seem that in the jaw, and probably in all other characters of the skull, as they become known, the Heidelberg race will be found to be a Neanderthal in the making, that is, a primitive, more pow^erful, and more ape-like ancestral form. In the matter of the retreating chin, the true Neanderthals of Spy, Malarnaud, Krapina, [ p. 101 ] and La Chapelle rank exactly half-way between the most inferior races of recent man and the anthropoid apes.
Not only among the Eskimos, but generally throughout the savage races of Australia and of other countries, the jaws are used as tools; among the Australians the teeth are very much worn down but are in admirable preservation. When seen from above, we observe that the ‘Heidelberg’ grinding-teeth form a perfect arch, or horseshoe-shaped arrangement, whereas in all the apes the two lines of grinding-teeth are almost parallel with each other. Thus, while there may be wide differences of opinion as regards the relationships of the Heidelberg man, all agree that Schoetensack’s discovery affords us one of the great missing links or types in the chain of human development.

The typical mammalian life of Second Interglacial times as found at Mosbach and Sussenborn belongs perhaps to a somewhat more recent stage of Second Interglacial times than that of the ‘Mauer sands,’ for in these localities the Etruscan rhinoceros [ p. 102 ] is wanting and the more specialized broad-nosed rhinoceros is abundant ; this animal differs from the Etruscan form in the possession of relatively long-crowned grinding-teeth, which were better adapted to grazing habits. On the head were borne two horns. A variety of the southern mammoth (E. trogontherii) is, so highly characteristic of Second Interglacial times that Pohlig refers to this life period as the E. trogontherii stage. From the structure of its grinding-teeth it is regarded as similar in habit to the Asiatic elephant, which now inhabits the forests of India, but it has the peculiar concave forehead distinctive of the mammoth and quite unlike the convex forehead of the Indian elephant. The bears of this period belong to the primitive species U. deningeri and U. arvernensis, for so far there is no certain record of the presence of the true brown bear of Europe (U. arctos). The sabre-tooth tiger of this time is preserved in the caverns of the Pyrenees near Montmaurin, associated with the remains of the striped hyaena (H. striata), a species which was widely distributed over western Europe in early Pleistocene times. This species was contemporary with, and later replaced by, the spotted hyaena (H. crocuta), from which the very hardy cave-hyaena (H. crocuta spelcea) of the ‘Reindeer Period,’ descended. We observe that the ‘polycladine’ deer of Upper Pliocene and First Glacial times has disappeared from western Europe; nor are there any traces of the axis deer. The hippopotamus is stiff represented by the giant species, H. major.
¶ Early Northern Migrations of the Reindeer
The animals that we have described belong in the warmer and more temperate regions of Europe. In the regions near the glaciers the reindeer was already to be found; in fact, this characteristically northern animal is recorded in the gravels of Süssenborn, near Weimar.
There is evidence of a succession of climatic changes in the region of Heidelberg. The Heidelberg jaw with its temperate mammalian fauna occurred at the very base of the Mauer bluff. [ p. 103 ] but higber up the bluff (Fig. 46) on a corresponding level are found the remains of mammals which indicate a marked lowering of temperature and which are referred by some authorities to the period of chilling climate that characterized northern Europe toward the close of Second Interglacial times. The reindeer also occurs in the ‘high terrace’ gravels of the River Murr, near Steinheim ; thus, at Mauer, at Sussenbom, and at Steinheim, we find proof that the reindeer had begun to spread over the colder regions of Europe, and there is some ground for belief that it found its way even as far south as the Pyrenees.
The evidence of the first cold, arid period which for the time greatly affected the climate of western Europe is also found in the layer of so-called ‘ancient .loess’ which lies in the bluff above the ‘sands of Mauer.’ This loess covers the warm mammalian deposits of the ‘sands of Mosbach’ as well as the ‘high terraces’ of many of the ancient river-valleys. Both in Europe and America the climatic sequence of the Second Interglacial Stage from moist to dry appears to have been the same.
Thus, after the recession of the ice-fields of the second glaciation, the climate was at first cold and moist ; then followed a long warm stage, favorable to the spread of forests; this was finally succeeded by a period of aridity in which the most ancient ‘loess’ deposits occurred. In Russia, also, the third glaciation was preceded by an arid and steppe-like cUmate with high winds favorable to the transportation of ‘loess.’
No palaesoliths or other proofs of human occupation have been found in this cold, dry period, for there is no evidence in any part of Europe of camping stations in this ‘ancient loess’ such as we find in the ‘loess’ which was deposited during the similar arid period toward the close of Third Interglacial and again during Postglacial times. Nor have we any record of the mammalian life in this ‘ancient loess’ of Europe.
[ p. 104 ]
¶ The Third Glaciation
This arid period in northern Europe and in North. America was followed by the moist, cool climate of the third glaciation. It is estimated by Penck that the advance of these new ice-fields began 120,000 years ago and that the period of advance and retreat of the glaciers was not less than 20,000 years. In the Alps the snow-line descended 1,250 metres below the present level; consequently this glaciation was more severe than the first but somewhat less severe than the second. In northern Europe the Scandinavian ice-field did not cover so wide an area as during the second glaciation, although Britain and Scandinavia were again deeply buried by ice; the glacial cap and glaciers flowed in a westerly and southwesterly direction across Denmark and the southern portion of the basin of the Baltic into Holland and northern Germany. In the Alps the third glaciation sent vast ice-floes along the valley of the Rhine, into eastern France, and into the valley of the Po, where this glaciation was even more extensive than the second. But the greatest glacier of this time was that of the Isar, a southern tributary of the Danube, which rises in the Bavarian Alps.38
During the Third Glacial Stage certain of the ‘middle terraces’ along the Rhine and other rivers flowing from the Alps were formed. In Britain,39 whereas during the second glaciation the ice-fields extended as far south as the Thames, during the third glaciation they did not extend beyond the midlands ; yet an arctic climate prevailed over southern England, with tundra conditions and temperature, as indicated by the plant deposits at Hoxne40 in Suffolk. Even before the third glaciation began in Europe a great ice-cap had formed over Labrador, on the eastern coast of North America, and the ice-sheets flowing to the south and southwest extended as far as Illinois, depositing the great Illinoian ‘drifts.’
[ p. 105 ]
Along the borders of these great ice-fields in both countries a cold and moist climate prevailed, for a prime condition of glaciation is the heavy precipitation of snow. In northern Europe, between the great Alpine and Scandinavian ice-fields of the third glaciation a cold climate undoubtedly prevailed; in the region of the Neckar River, near Cannstatt, is found a deposit knowm as mammoth loam,’ which Koken believed to be contemporaneous with the period of the third glaciation, although the evidence is certainly not convincing.41 Here are found fossil remains of the bc^dinavian reindeer, also of two very important new arrivals in Europe from the tundra regions of the far northeast, ar,imglg [ p. 106 ] wHch had wandered along the southern borders of the Scandinavian ice-sheet from the tundras of northern Russia and Siberia* This is the first appearance in western Europe of the woolly mammoth (E. primigenius) and the woolly rhinoceros (D. antiquitatis). In this tmammoth loam’ there also occur two species of horse, the giant deer (Megaceros), the stag, the wisent, and the AurochSc If the woolly mammoth and the woolly rhinoceros actually entered eastern Germany at this time, they certainly retreated to the north with the approach of the warm temperate climate of the Third Interglacial Stage, because no trace of these animals has been found again in Europe until the advent of the fourth glaciation.
¶ Bibliography
(1) Gaudry, 1890.1.
(2) Smith, G. E., 1912.1, p. 582.
(3) Op. cit.
(4) Munro, 1893.1.
(5) Osborn, 1910.1, pp. 306, 307.
(6) Geikie, J., 1894.1, pp. 329-336; 1914.1, p. 227.
(7) Dawkins, 1883.1, pp. 576-579.
(8) Geikie, J., 1914.1, p. 248.
(9) Reid, C., 1908.1.
(10) Desnoyers, 1863.1.
(11) Haug, 1911.1, p. 1S07.
(12) Geikie, J., 1894.1, p. 682; 1914.1, p. 250.
(13) Dubois, 1894.1.
(14) Fischer, 1 913. 1.
(15) Schwalbe, 1899.1; 1914.1.
(16) Büchner, 1914.1.
(17) Volz, 1907. 1.
(18) Elbert, 1908.1.
(19) Selenka, 1911.1.
(20) Pilgrim, 1913.1.
(21) Schwalbe, 1899.1, pp. 227, 228. (22) Op. cit., p. 223.
(23) Schwalbe, 1914. i, pp. 601-606.
(24) Büchner, 1 914.1, p. 129.
(25) Dubois, 1894,1, p. 14.
(26) Keith, 1912. 1.
(27) Smith, G. E., 1912. 1, p. 595.
(28) Op. cit.
(29) Rutot, 1907.1.
(30) Boule, 1913. 1, pp.‘266, 267.
(31) MacCurdy, 1 905.1, pp. 468, 469.
(32) Geikie, J., 1914.1, p. 251.
(33) Op. cit. p. 255.
(34) Op. cit. p. 238.
(35) Schoetensack, 1908.1.
(36) Op. cit. pp. 25-43.
(37) Bonarelli, 1909.1.
(38) Penck, 1909. 1.
(39) Geikie, J., 1914.1, p. 258.
(40) Op. cit., pp. 257-262.
(41) Schmidt, 1912.1, p. 181
¶ Footnotes
A recent article by A. Smith Woodward describes the fourth known specimen of Dryopithecus, lately discovered in northern Spain (see Woodward, 1914.2). ↩︎
There is a vast Pithecanthropus literature. That chiefly utilized in the present description includes Dubois,13 Fischer14 Schwalbe,15 Buchnerd16. ↩︎
In the Trinil skull as restored by McGregor (Fig. 36) the cranial capacity is 900 c. cm. ↩︎
These horses are now identified respectively as E. mauerensis, E. moshachensis, and E. süssenbornensis. ↩︎
This glaciation as it occurs in northern Europe has been termed Polandian by Geikie ; in the Alps Penck has termed it the in America it is known as the Iliinoian from the great drifts it deposited over the State of Illinois. ↩︎