Public domain
[ p. 678 ]
¶ Formations and Physical
History During the closing stages of the Paleozoic, when the sea was excluded from the area between the growing Appalachians and the Great Plains, Appalachia appears to have suffered deformation. One result of this deformation was the development of elongate troughs upon its surface, roughly parallel to the. present coast. These troughs became the sites of deposition, and the sediments laid down in them constitute the only representative of the Triassic system in the eastern part of the continent. The open sea seems to have been completely excluded from the western interior by the beginning of the Triassic period, though sedimentation was in progress over considerable areas between the meridians of 100° and 113°. Some of these areas appear to have been the sites of salt seas and some of fresh lakes, while still others were probably without standing water. Between the meridians named, many areas of relatively high land probably interrupted the continuity of sedimentation. On the western coast, the ocean began to gain on the continent about the close of the Paleozoic, and the shore of the Pacific was presently shifted eastward to the vicinity of the 117th meridian in the latitude of Nevada.
In keeping with these changes in geography, Triassic strata are known in three regions: (1) The Atlantic slope east of the Appalachians; (2) the western interior; and (3) the Pacific coast. The strata in these three regions are in many ways unlike.
The Eastern Triassic — The Newark Series
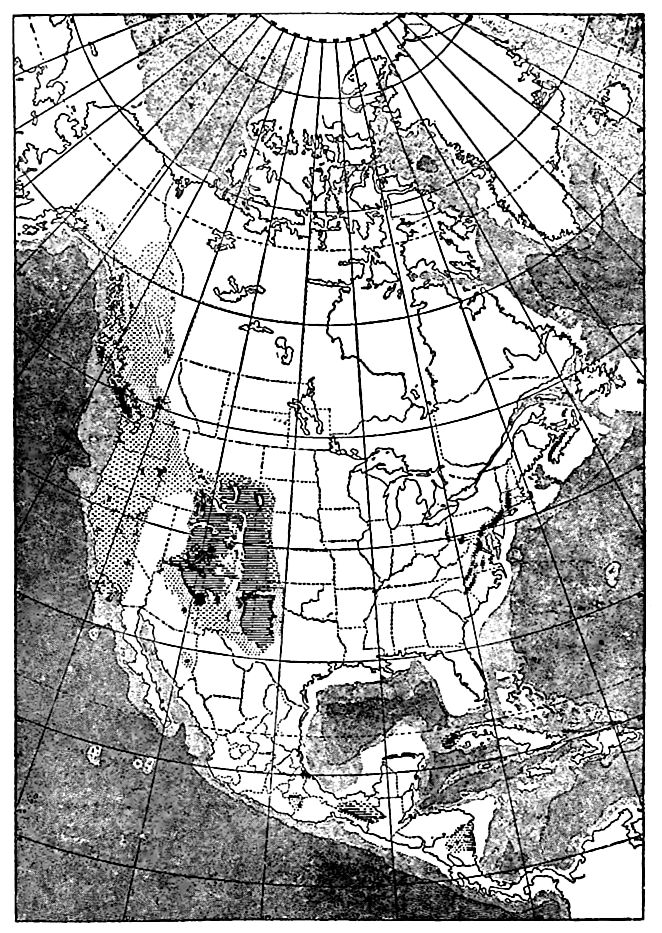
Distribution. The Triassic system of the east occurs in spots from Nova Scotia to South Carolina, as shown in Fig. 464. Its [ p. 679 ]several areas are mostly elongate in a northeast-southwest direction. The principal areas are: (1) About the Bay of Fundy; (2) in the Connecticut River valley; (3) from the Hudson River to Virginia; and (4) disconnected areas in Virginia and North Carolina. The beds of these several areas have been grouped under the mime Newark[1] (Newark, N. J.).
[ p. 680 ]
The rocks of the Newark series[2] include all the common varieties of fragmental rocks, some of which are developed in unusual phases. Sandstones and shales predominate, but there are abundant conglomerates, some breccias, and, locally, limestone and coal.
The conglomerates. Conglomerate lies at the base of the system in many places, and is made up chiefly of material from the underlying crystalline schist. But the conglomerates are not simply basal. Some of them represent the border phase of beds which grade laterally into sandstone, and even into shale. The chief constituent of the conglomerate is quartz, the most resistant part of the underlying terranes; but quartzite and crystalline schist appear in the conglomerate, and locally limestone is its principal constituent.
The exceptional coarseness of the conglomerate in some places has been thought to call for unusual conditions of origin. It has been conjectured that it was formed at a time when glaciers existed in the region. It should be noted, however, that this suggestion was based on the supposed demand for some exceptional agent of transportation, rather than on any direct evidence of glaciation. In general, the materials of the conglomerate are too well assorted to be the immediate product of glaciation, and the stones and bowlders do not bear the marks of ice. These points would lose much of their force if the conglomerate were deposited by glacial drainage; but in the absence of all certain evidence of glacial or glacio-fluvial origin, it seems more prudent to regard the conglomerate as a formation of terrestrial or shallow-water origin.
The sandstone and shale. Sandstone and shale make up the [ p. 681 ] great body of the Newark series, and both possess distinctive characteristics. Their prevalent color is red, though there are shales which are black, and sandstones which are gray. Except locally, the series is poor in fossils. Some of the sandstone is arkose, that is, contains a considerable amount of feldspar, and both sandstone and shale contain much mica. Both these constituents abound in the metamorphic rocks from which the Newark sediments were chiefly derived.
Conditions of origin. The character of the Newark formations and their fossils, mainly land plants, footprints of reptiles, and fresh- or brackish-water fishes, indicate that they are of continental rather than marine origin, though the precise manner in which they were laid down is not known. That deformation of the surface of Appalachia, which had been reduced nearly to planeness by erosion, gave rise to elongated depressions in which the Triassic sediments were deposited, seems certain. The depressions may have been due to warping or to faulting, or partly to the one and partly to the other (Figs. 465 and 466), and their development may have progressed as deposition went on. Some of them may have been broad river valleys, which, in the general uneasiness which marked the close of the Paleozoic era, became sites of deposition. However formed, these depressions in the surface of the present Piedmont region became the sites of lakes, bays, estuaries, dry basins, or aggrading rivers. Lacustrine, estuarine, and fluviatile conditions may have alternated from time to time in the various places where sedimentation was in progress, and perhaps the sea gained access to some of them from time to time.
The considerable thickness of the sediments, taken in connection with their decisive evidences of shallow-water or subaerial origin, such as ripple-marks, sun-cracks, tracks of land animals, etc., indicate either that inclined deposition prevailed, or that subsidence of the areas of sedimentation accompanied the deposition. For the adequate supply of the detrital material, it would seem that the lands bordering the areas of deposition were raised, relatively, as the troughs were filled. The general conditions of accumulation may have been similar to those under which the Catskill formation was deposited at an earlier time.
[ p. 682 ]
Former extent. It is possible, and perhaps probable, that the areas of the Newark series from Virginia to South Carolina were once connected with one another, and with the Virginia-New York area, though such connection has not been demonstrated. It has even been suggested that the Newark of the New York- Virginia area was once connected with that of the Connecticut valley, and this with that of Acadia, and that the separation was effected by erosion; but this suggestion does not seem well founded.[3]
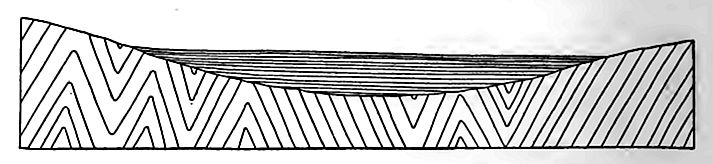
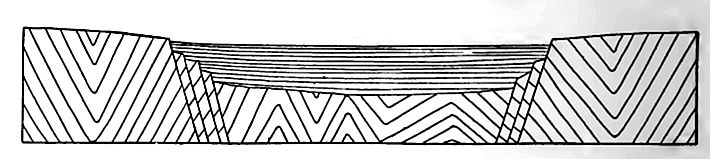
Igneous rocks. Igneous rocks are associated with the sedimentary beds in dikes and in sheets interbedded with the shales and sandstones. Some of the sheets are extrusive, having been poured out on the surface of the inferior beds and subsequently covered by sediment; others are intrusive (sills), having been forced in between the layers of sedimentary rocks after the latter were deposited. Certain isolated bodies of igneous rock may represent volcanic plugs. The sheets of igneous rock (usually called trap, though largely basalt) vary in thickness from a few feet to several hundred.
[ p. 683 ]
Structure. The structure of the Newark series is generally monoclinal. In the Connecticut River valley the dip is about 20° (10° to 25°) to the eastward. In the New York- Virginia area[4] also the structure is monoclinal, the dip being to the northwest (10°-15°). This contrast of dips between the New England and New Jersey areas .has been thought to support the suggestion that the strata of the two areas are parts of one huge anticline, from the broad crest of which the beds have been removed. The strata are otherwise somewhat deformed, though never closely folded. The series is faulted extensively, and in a somewhat complicated manner.
Thickness. On account of the faulting, the thickness of the Newark series is difficult of determination. In the Richmond area of Virginia, it is estimated at something more than 3,000 feet; in New England, at 7,000 to 10,000 feet; and in New Jersey even more. Correlation. The stratigraphic relations of the Newark series in the United States would not determine its age. It lies unconformably on rock which is mainly pre-Cambrian, and is overlain unconformably by Comanchean (Lower Cretaceous) beds. About the Bay of Fundy, however, the rocks lie unconformably on the early Permian. The physical relations of the Newark series therefore show that it is post-early-Permian, and pre-Comanchean. Between the Permian and the Comanchean there are two periods, the Triassic and the Jurassic. In referring the series to the former, the chief reliance is on the fossils, and on the same basis it is believed to represent only the later part of the period.[5]

[ p. 684 ]
In the West
The deposits of the western interior.[6] The interior area of set li mentation, chiefly between the 100th and 113th meridians, had its southern limit, so far as now known, near the southern boundary of the United States, while at the north it extended into Canada. This area is believed to have been cut off from the Gulf by land in eastern Texas, or if it had connection with the sea, it was probably slight. Into this interior area of sedimentation, detritus was borne from the surrounding lands. Some of the deposits were probably laid down by streams, some in fresh lakes, and some in bodies of salt water, as in the Permian period. Fresh-water fossils (unios) are found locally, as in New Mexico. The structure of some of the sandstone is such as to suggest an eolian origin.
The deposits of the period are in large measure concealed by hit er beds, but they are exposed at various points where the strata have been warped, and the overlying beds removed by erosion. The most easterly outcrops are in Texas, Oklahoma, and South Dakota. The system may underlie later formations between these localities and the Rocky Mountains. Red beds which are thought to be Triassic outcrop interruptedly along the eastern base of the Rocky Mountains from British America to New Mexico. These beds are thin, and contain more or less gypsum and sometimes salt. Ocasionally they contain fossil leaves.
Farther west, Red beds have representation among the surface rocks. Some of these beds are perhaps Triassic; but in much of the western interior, the undifferentiated Triassic and Permian rest conformably on the Carboniferous (Pennsylvanian) , though occasionally they overlap it and rest upon pre-Cambrian formations. In southwestern Colorado and eastern Utah, the Trias rests nnconformably on older, deformed, unf ossiferous red beds (presumably Permian) and on strata of Pennsylvanian age.[7]
Thickness. In the eastern part of this western area, t he Tri system is thin, sometimes no more than 100 feet. To the west it [ p. 685 ] thickens, reaching to 2,000 to 2,500 feet in the Uinta Mountains, beyond which it again becomes thinner.
On the Pacific Slope[8]
The Triassic system has its greatest development in America along the Pacific coast. In the latitude of Nevada, the Pacific seems to have extended eastward over the site of the Sierras to longitude 117° (approximately). The shore line of the Pacific farther north has not been definitely located. It was probably irregular, and, in general, several degrees farther east than now, well up into British Columbia. Between the latitudes of 55° and 60°, the sea is believed to have crossed the present Cordilleran [9] belt, though this northern bay east of the Rockies was probably nob connected freely with the areas of sedimentation in the western interior. Marine beds of the Lower Trias are found as far east as eastern Idaho, and the sea probably reached this region by way of Utah, Nevada, and Southern California. Later formations of the system are not found east of western Nevada and eastern Oregon.[10] The Triassic system is found as far north as Alaska.

[ p. 686 ]
The published measurements assign the system the great thickness of 17,000 feet (maximum) in the West Humboldt range of Nevada, where it rests on pre-Cambrian terranes. To have supplied such a volume of sediment, the land to the east must have been high, or repeatedly renewed, to counterbalance the waste, unless the great thickness is due to oblique deposition.
The succession of faunas in the Trias of the Pacific coast indicates that considerable changes in the physical geography of the northern Pacific were in progress during the period. These will be referred to in connection with the faunas of the system.
Climatic Conditions
The wide distribution of gypsum and salt in the Triassic system, not only of America but of Europe, is good evidence of wide-spread aridity. The prevalent redness of the system, in other continents as well as our own, is also commonly regarded as an indication of aridity. Some of the peculiarities of the Newark conglomerate also might find their explanation in aridity. In an arid climate, expansion and contraction due to changes of temperature are so great as to be effective in disrupting rock if it is not covered by soil. Under such circumstances, much coarse debris originates, largely of rock which is undecomposed. Violent storms (cloud-bursts), which often characterize arid climates, might account for the transportation of debris from the place of its origin to the place of its deposition. For the formation of abundant debris in this way, steep slopes are needful, for gentle slopes and flats soon get a covering of mantle rock which prevents the disruption of the rock beneath. If this was the origin of the coarse materials of the conglomerate, their rounding and wear would have to be attributed to the waves of the body of water in which deposition took place.
[ p. 687 ]
Close of the Trias
Considerable geographic changes marked the close of the Triassic period in eastern North America, bringing the areas which had been the sites of deposition to higher levels, faulting the rocks, and affecting them by igneous intrusions. These changes were comparable in extent and importance to the changes which separate the various systems of the Paleozoic, but they were not of continental extent.
In the western part of the United States, the separation of the Triassic period from the Jurassic was not pronounced, and the sedimentary history of much of the western half of the continent seems to have run an uninterrupted course from the beginning of the Permian to the later part of the Jurassic. The case may have been somewhat different north of the United States, for in British Columbia and in the adjacent islands, Triassic and older formations were upturned, deeply eroded, and again submerged before the beginning of the Cretaceous. The great igneous formations associated with the Trias of the northwest appear to have been made during the Triassic period, rather than at its close.
¶ Foreign Triassic
Europe
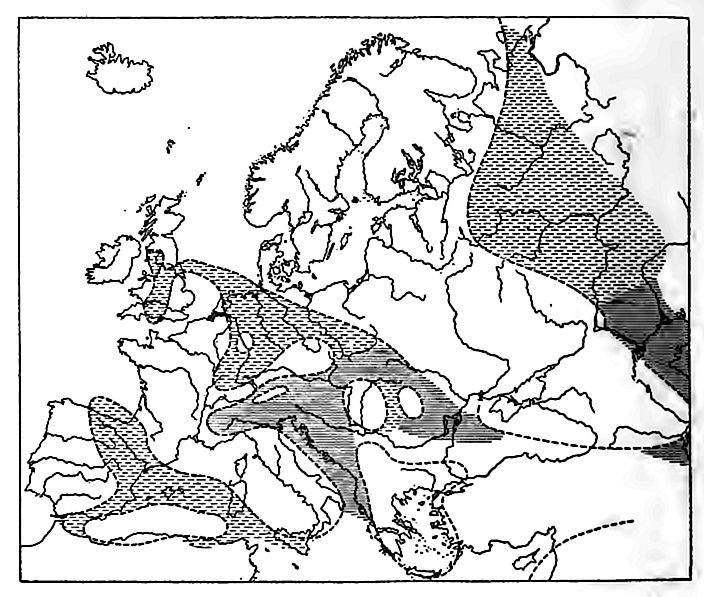
In Europe, the Trias is exposed in many widely separated places, the largest being in northwestern Russia; but the system is better known in the western part of the continent. In England, it is unconformable on the Permian, but on the continent, generally conformable.
The system has a marine and a non-marine phase. The nonmarine (or Triassic) phase prevails throughout the northern part of the continent, while the marine (or Alpine) phase is found farther south. The former resembles the Permian of Europe, and the Permian and Triassic of the United States.
In general, the Upper Trias is more wide-spread than the Lower, especially in the southern part of the continent, and is marine over [ p. 688 ] a wider area. The principal subdivisions recognized in Britain and Germany are the following:
| Britain | Germany |
|---|---|
| Rhaetic | Keuper |
| Upper Trias | Muschelkalk |
| Lower Trias | Bunter |
Northern Europe. The Bunter formation of Germany was deposited chiefly in lakes, inland seas, and on land, as shown by the fossils, the beds of salt and gypsum, and the dune structure of some of the sandstone. The name has reference to the high colors of the formation, red predominating. Toward the top of the formation, thin beds of marine origin are intercalated locally with the others, showing that changes in the relation of land and water were in progress, and that the sea gained on the land toward the close of the epoch.
[ p. 689 ]
The Muschelkalk is a limestone formation and records the further encroachment of the sea. Its fauna has been thought to indicate that the water in which it lived was not the open ocean, but a body of water comparable to the Black sea or the Baltic [11].
The Keuper formation resembles the Bunter, and, like it, is marine in its upper portion, and is followed by the marine beds of the Jurassic period. The Keuper contains a little coal (not workable), a common accompaniment of shallow-water and marsh formations.
In England the system is often known as the New Red Sandstone, though formerly the Permian was also included under this term. It differs from the Trias of Germany chiefly in that the marine member of the German system is absent in England. Both salt and gypsum occur in workable quantities in some parts of England.
In southern Sweden, the Trias contains coal. The system here was probably once continuous with that of Germany, and may still be, for borings show that it underlies parts of the North German lowland. The Triassic beds of most of Russia are similar to those of western Europe.
The non-marine formations of red color so characteristic of the Triassic system both in North America and Europe afford another striking inter-continental analogy, and doubtless point to a common cause, or to similar wide-spread conditions.
Southern Europe. The Alpine or marine phase of the Triassic has its best development in the eastern and southern Alps, and is made up of thick beds of limestone, often dolomitic, alternating with thinner beds of clastic rock. The limestone and dolomite are much more resistant than the associated shales, and as a result, erosion has developed a distinctive topography (Karst topography) at several points in the southern Alps — a topography so striking that the localities where it is seen have become the objective point of travel, both for geologists, and for lovers of wild and picturesque [ p. 690 ] scenery. In these regions the dolomite (limestone) stands up in bare, bold-faced walls, peaks, and towers, surrounded and separated by valleys and passes clothed with abundant vegetation. The decay of the projecting limestone leaves little soil behind, and the little which is formed is promptly carried away by wind and rain.
The Trias of the Italian Alps is the source of the Carrara marble. The Trias of the western Alps is largely non-marine, and in some parts of Switzerland the Upper Trias contains coal and igneous rocks.
Other Continents
Asia. The marine phase of the system, similar to that of southern Europe, is continued eastward to southern Asia. It is also found in the high latitudes of Asia, including numerous islands north of the mainland. The Trias is generally conformable on the Permian and beneath the Jurassic.
South America. No marine deposits of Triassic age are known east of the Andes, but coal-bearing Trias occurs in Argentina and Chile, and marine beds at various points in the Andes. Thus it is clear that the site of parts of this great system of mountains was beneath the sea in the Triassic period.
The Triassic system is represented also in South Africa, Australia, New Zealand, and New Caledonia.
¶ The Life of the Triassic Period
The remarkable physical conditions that dominated the land and impoverished its life in the Permian period still held sway during the earlier part of the Triassic. In their general biological aspects, as in their physical conditions, the two periods were much alike. Toward the close of the Triassic there was a pronounced change. The land became lower and the sea encroached upon it, bringing about appropriate changes in life. Nearly all that is known of North American Triassic life belongs to the later portion of the period.
[ p. 691 ]
The Plant Life
The record of the vegetation is very imperfect, and it was probably scant in reality, for broad saline basins and arid tracts are inhospitable to plants. The Triassic was distinctly an age of gymnosperms. Ferns and fern-like plants still held an important place, and the Equisetales were more important than now; but the dominance of these types was past. The great lycopods, too, were almost gone, though sigillarias were among their lingering representatives. Among gymnosperms, the cordaites had declined, but conifers of the types that came in during the Permian, and kindred new ones, were prominent. The cycadean group occupied the place of central interest. The Bennettitales, formerly called cycads, abounded, and from them the true cycads sprang later. The ginkgos (Ginkgoales) diverged from the ancestral cordiates at about this time.
The calamites had given place to true equiseta, which were represented by forms that were gigantic in comparison with modern types. In the far east and in the southern hemisphere, the Glossopteris and its allies constituted a marked feature of a flora whose general aspect was much like that of the preceding Permian in the same regions.
The Triassic floras of Europe and America, so far as known, were much alike, and both bore a scrawny pauperitic aspect that reflected the hostile conditions against which they struggled, conditions for which the stunted conifers of to-day stand as representatives.
In the closing stages of the period, a much ampler flora seems to record some amelioration of the inhospitable conditions. The larger part of the known American fossils belong to this stage. The Richmond coal-beds of the Newark series, probably the product of marsh vegetation, contain great numbers of equiseta and ferns, but almost no conifers and few cycadeans. A few plant fossils have been found in Mexico, Arizona, and California.
The Land Animals
The physical conditions of the Permian and Triassic periods were so similar that adaptation to the conditions of the first would [ p. 692 ] seem to have been a fitting preparation for life in the second. Yet, in spite of this fact, there was a great break in the succession of land life, so far as the known record shows. What became of the Permian vertebrate faunas of North America is unknown, for between the horizons yielding Permian fossils of land animals, and those yielding corresponding Upper Triassic fossils, there are great thicknesses of red sandstone barren of fossils of all sorts, so far as now known, and the Upper Triassic (Keuper) fauna does not appear to have descended from the Permian. In Africa there appears to have been a much less serious break between the land life of the Permian and that of the Trias. In other continents few fossils of land life of the early types have been found, and the record of the Middle Trias is but little less meager. Of the life of the Upper Trias the record is much fuller. The period was one of great transitions, in which many types were initiated, while only a few were carried to their maximum development.
There is abundant proof of the intermingling of European and American land faunas late in the Triassic period, for at this time there were, in North America, representatives of groups that had lived in Europe since the early Permian, but which had never before appeared in our continent, so far as now known.
The amphibians had lost the foremost place they held in the Permian, though still numerous (but not in North America in the early part of the period). Before its close, however, they entered upon a rapid decline, from which they never recovered. Ancestors of the whole tribe of terrestrial vertebrates, they soon became its most insignificant representatives.
The strange ancestral reptiles evolved rapidly. The branch with the mammalian strain (p. 672,Fig. 470) seems to have been left far behind by the more distinctively reptilian branch, which developed greatly in the closing stages of the period when the dryness was ameliorated and vegetation began again to flourish. Refore the close of the period, every important group of the class had its representatives. The crocodilians, the flying saurians, and the scaled reptiles (lizards, snakes, etc.) came in near the close of the period, as some of the older types were disappearing.
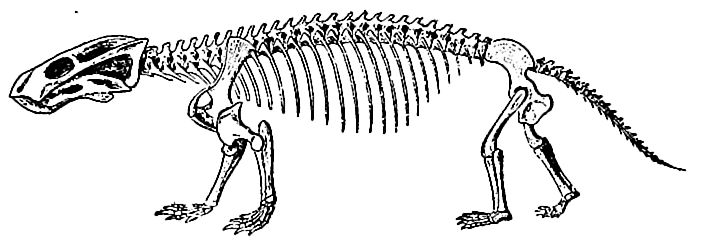
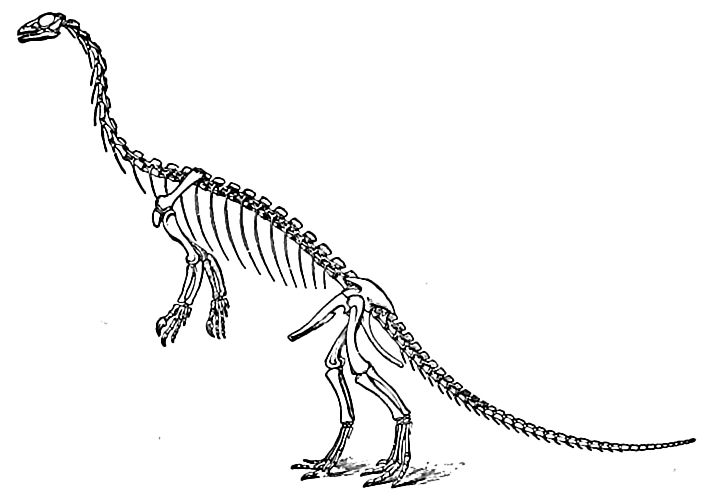
A foremost feature of the life was the advent and rapid evolution [ p. 693 ] of the dinosaurs (terrible saurians), the reigning reptilian dynasty of the era. They were at first of generalized types, but later became more specialized, and diverged widely. While some were small and delicate in structure, others were gigantic and ungainly. Carnivorous forms only (Theropoda) are known in the Trias, and most of them were not especially large. Their general form is indicated by the partially restored skeleton shown in Fi^. 471. The strong development of the hinder parts, the relative weakness of the fore limbs, and the kangaroo-like attitude, are the most obvious features. The bones of these upright-walking forms were hollow, and certain other structural features resemble those of birds. The reduction of the functional toes of the hind feet to four, with one of them much shorter than the others, caused their three-toed tracks to be mistaken for those of birds, until recently. Even in the Triassic period, the dinosaurs ranged widely, living in the Rocky Mountains, along the Atlantic coast from Carolina to Prince Edward Island, in western Europe, India, and South Africa.
[ p. 694 ]
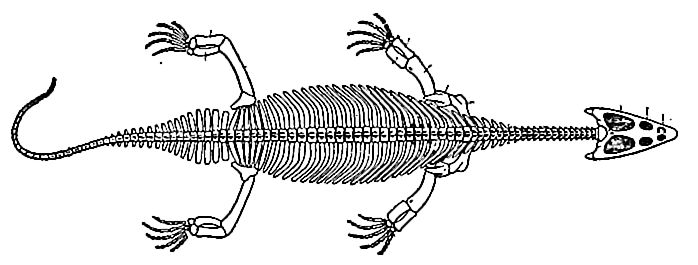
Before the close of the period both branches of the reptilian tribe sent delegations to sea, the one represented by the ichthyosaurs (Fig. 489) and the other by the plesiosaurs. It is not difficult to find good reasons for this sea-ward movement. Besides the tendency of every masterful race to invade all accessible realms, the notable extension of the sea before the close of the period invited the adaptation of land animals to an aqueous habitat, for the shallow waters with their prolific life, creeping in upon the land, set tempting morsels before the voracious reptiles, while the reduction of the land areas and their terrestrial feeding-grounds, intensified by their own multiplication, encouraged a movement to the sea.
The advent of mammals. Of peculiar interest is the appearance of early forms of non-placental mammals. They were small, and so primitive in type that it is not altogether certain that they were mammals; but they are commonly regarded as such, with kinship to the marsupials. Their appearance while reptiles were yet dominant suggests that mammals diverged from the primitive stock much earlier. In view of the mammalian dominance of later times, it is note-worthy that the non-placentals developed but slowly and feebly during the Mesozoic era. It is an open question whether placental mammals are the descendants of the Mesozoic non-placentals, or whether they had an independent origin.
[ p. 695 ]
The Marine Life
The reduction of marine life of the shallow-water type during the Permian (p. 673) was continued into the Triassic period, and since its remains are in sediments now buried, such record as it made is mainly concealed. This was especially true of North America. To trace the shallow-water marine life of the Permian into that of the next period, it is necessary to bring together evidence from different continents. The question of supreme interest is the way in which the epicontinental sea life, crowded to a minimum habitat between the land and the deep sea, maintained its continuity, transformed its species, and later re-peopled the shallow waters when they again became more extensive in the closing stages of the Trias and later.
When the sea readvanced on the North American continent, it was chiefly from the Pacific, but there were also incursions up the MacKenzie Valley and from the Gulf of Mexico. It is not clear that the sea completely withdrew from the present land area on the Pacific coast after the Permian; but the fossils so far recovered from the Trias of this region do not give clear evidence of continuous submergence of an area such as to allow the development of a definite provincial fauna.
The transition tracts. It was otherwise on the Eurasian continent. While the sea withdrew from the northwestern part of [ p. 696 ] Europe during the Permian period, it lingered about the Mediterranean, in Russia, Turkestan, and northwestern India, and probably on the continental platform in or near Siberia. The Mediterranean, the Himalayan, and the Siberian regions are the best known tracts into which the shallow-water marine life of the late Paleozoic retreated and underwent transformation into the early provincial faunas of the Mesozoic. It is quite certain that there was at least one other area where important faunal reorganization took place at about this time, for a notable fauna suddenly appeared in the Middle Triassic, which does not seem to have originated in any of these three districts.
In each of these three areas an important remnant of Paleozoic sea life seems to have undergone a radical and perhaps rather rapid evolution, such as might be anticipated from the crowding of the great faunas of earlier times into such limited areas. From these areas the new faunas spread when the sea again extended itself upon the land.
The transition faunas. The most complete record of the transition from Paleozoic to Mesozoic marine life is found in India. Beds containing fossils characteristic of the Permian are overlain conformably by beds containing forms characteristic of the Mesozoic. In the Permian beds there are forms foreshadowing the Mesozoic types, and in the beds above there are Permian types that lived on and mingled with Mesozoic forms. The transition fauna of the Mediterranean region appears to have been less rich than that of India. Concerning the early stages of the Siberian fauna, little is known; but its peculiarities, as revealed in a later stage of the early Trias, leave little doubt of its independence of origin.
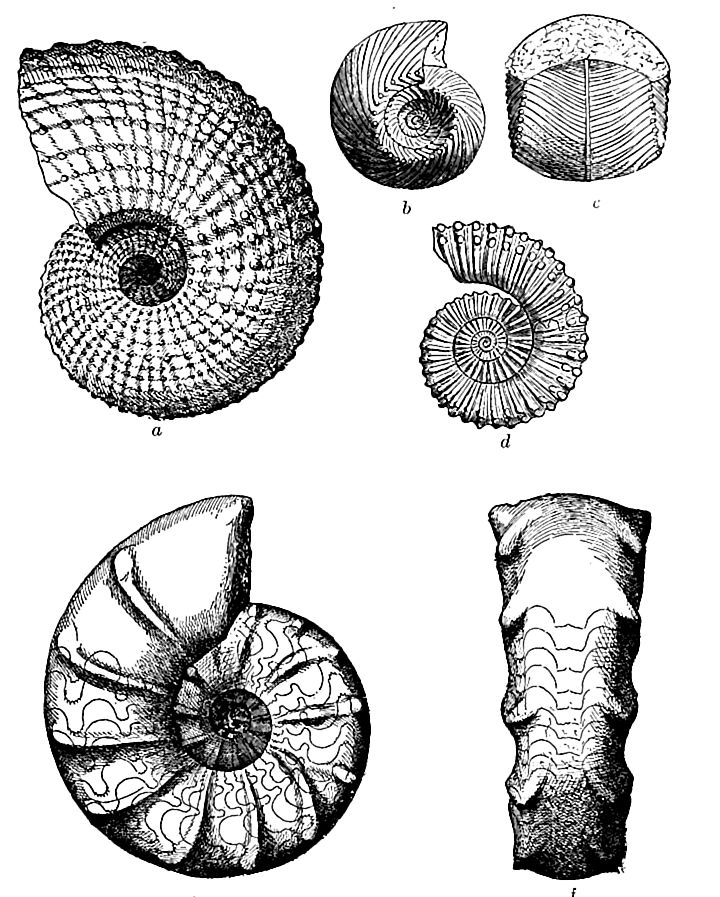
General nature of the faunal changes. In nearly all Paleozoic faunas, brachiopods were a leading element of the marine faunas, while trilobites, crinoids, and corals, each in turn, gave distinctive character to the successive faunas. In the Mesozoic era, the ammonites took the first place, followed by the pelecypods and the gastropods. The ammonites (Fig. 473) are peculiarly fitted for distinguishing successive horizons, not only because they were free forms, measurably independent of bottom conditions, but because they were steadily and rapidly advancing in organization, [ p. 697 ] and because their shells were so constituted as to record their progress.
[ p. 698 ]
The earliest fauna was markedly restricted. This may be more apparent than real on account of the imperfection of the accessible record; but it was doubtless real in some measure, and due to the physical limitations already sketched. At the same time, there was an increase in differentiation. The conditions which repressed the life, reducing the number of individuals, species, and genera, forced them to diverge more and more, in order to accommodate themselves to the conditions inhospitable to life. This is shown best in the development of the land and fresh-water life, but to some extent also in the marine life.
The geographic suggestions of the faunas. A great group of ammonites, embracing more than 200 species, formed the leading features of the early Indian fauna. The alliance of the Indian forms with those of North America is so close as to indicate that before the close of the early Trias, migratory connections had been established between India and western America.
Somewhat later in the early Trias there appeared in the Siberian region (Olenek River) a fauna having some of the same genera as the Indian. Closely related species are found in Idaho. If there was connection between the Indian and Siberian regions, it would be possible for Indian species to reach America either by way of Siberia and the Arctic coast, or by the Pacific sea-shelf, and slight changes, involving submergence or emergence in the region of Bering Strait might change the combination of the faunas.
The Indian and Siberian provinces seem to have been distinct from the Mediterranean province throughout the earlier Triassicj but in California a few fossils have been found which are characteristic of the earlier Triassic of southern Europe.
The early Triassic faunas of central Europe were very variable, a part being developed apparently in fresh water, a part in isolated seas, and a part perhaps in dependencies of the ocean. The marine life was scanty, and its origin and relations uncertain; but it seems to have been largely independent of the Mediterranean basin.
By the middle of the Triassic period the faunas had begun to intermingle, and to lose their provincial characteristics. The Mediterranean [ p. 699 ] fauna gained access to the Indian basin and to our western coast, and counter-migrations were of course made possible. At about the same time, the Siberian fauna had access to western United. States.
During the later stages of the period, a rich marine fauna flourished in California. Many of its species were identical with those of the Mediterranean and Himalayan regions, or closely allied to them. It is therefore inferred that these provinces were in free communication, so far as marine life was concerned, with the west American coast. The Upper Trias of British Columbia, on the other hand, contains a different fauna, including a type that belongs to the Siberian group. The British Columbian fauna is perhaps to be regarded as the descendant of the Idaho fauna of the early Trias, with additions from Siberian sources, while the California fauna is perhaps a derivative from the Mediterranean and Himalayan provinces by some different route. Present knowledge, however, is not sufficient to show the precise nature of the migrations between Europe, Asia, and America during Triassic times.
The most conspicuous feature of the Triassic faunas was the reascendancy of the cephalopods in the form of the ammonites, which had a marvellous development during the period, reaching a thousand species. Their evolution was the more notable because the structural changes were conspicuous, and showed plainly the advance of each stage over the preceding. While early types still persisted, the closely coiled, intricately-sutured forms predominated. The first known cephalopods of the cuttlefish type appeared at this time. The deployment of the cephalopods was therefore more varied and comprehensive than ever before, though they did not reach their culmination till the next period. Old forms, orthoceratites and goniatites, made their last appearance in this period. The remarkable commingling of old and new types makes this one of the most instructive assemblages in the history of the cephalopods.
A similar commingling of transitional forms was presented by the i_. The progress of the bivalves was scarcely less real, though they do not show the transition from ancient to modern so conspicuously. Their numbers were large, and most of their genera [ p. 700 ] modern, some being identical with those now living. With the modern types there were about half as many that still bore a Paleozoic aspect.
The dominant brachiopod types of the late Paleozoic were distinguished by extended hinge lines (spirifers, orthids, etc.), while the narrower beaked or rostrate forms were in a respectable minority. In the Triassic period, the rostrate forms (Rhynchonella, Terebratula, and allied genera) became predominant, and have remained so ever since. (Compare Figs. 474 and 419.)

Although echinoderms are not abundant in the Triassic fauna, the period marks the transfer of leadership from the crinoids to the sea-urchins, and a structural change in the latter. Beginning with the Triassic, the echinoids had twenty rows of plates in belts of two rows each, whereas the Paleozoic forms had more. At first they retained the previous pentamerus symmetry, but later this gave place to a bilateral symmetry. Starfishes and brittle-stars were present, but not abundant.
[ p. 701 ]
While corals were rare in most places, they were rather abundant in favored localities. Some of them resembled the Paleozoic forms in being simple and cup-shaped, but the compound species took on the modern (hexacoralla) form, and the compound Paleozoic (tetracoralla) type disappeared. These later compound corals do not seem to have descended from the compound Paleozoic forms, but from some simple type.
While the general aspect of the Triassic marine faunas was revolutionary, it is important to note, in view of beliefs once current, that it was transitional, and not an abrupt substitution of a new fauna for an old one. Paleozoic types lived side by side with later forms, though usually represented by new genera. This overlapping and commingling of old and new clearly indicates the gradation of the earlier into the later. The transition was extraordinary in the apparent rapidity of its progress, and in the extent to which it affected all classes. The fact that most of the new types were already present in the earliest Triassic, indicates that the transition was chiefly in the Permian. The fundamental cause was, with little doubt, the readjustment of the earth’s surface to internal stresses, and the physiographic and climatic changes consequent upon this readjustment.
Marine reptiles seem to have thriven in the western part of our country, especially in the middle and later Trias. The numerous ichthyosaurs found in the later Triassic beds, and their peculiarities, suggest that this may have been a center of dispersion of these reptiles. With the ichthyosaurs were other reptiles (thalattosaurs) unknown elsewhere.[12]
¶ Notes
For an account of the Newark series sec Russell, Bull. 85, U. B, GeoL Surv., 1892. Full bibliography to date of publication. ↩︎
The Connecticut valley and New York- Virgin i.i areas are beel known. and the descriptions of the formations here given apply especially to them. ↩︎
For summary of the Trias of Connecticut, sec Davis, 7th Ann. Rept., U. S. Geol. Surv., or a fuller and later account in the 1 8th Ann. Rept., Pt. II Sec also the Holyoke folio, l". S. Ceo!. Surv., and for the Pomperaug area, Hobbs, 21st Ann. Rept., U. S. Geol. Surv., Pt. III. ↩︎
For summary of the Newark of New York and New Jersey, see Kummel, Rept. of the State Geologist of New Jersey, 1896, and Jour. Geol., Vol. VII. ↩︎
Rice and Gregory would make the Newark the approximate equivalent of the Keuper of Europe. Bull. 6, State Geol. and Nat. Hist. Surv. of Conn. ↩︎
There is some doubt about the age of most of the beds formerly referred to this system. The tendency of later study has been to refer more and more of them to the Permian. See references under Permian, and Hill. Physical Geography of the Texas Region, folio U. S. Geol. Surv. ↩︎
Cross: See footnote p. 602. ↩︎
King, Geol. Surv. of the 40th Parallel, Vol. I, gives an account of the Triassic as far west as the Sierras in this latitude. See also many of the California folios of the U. S. Geol. Surv., and the Roseburg, Ore., folio. ↩︎
Dawson, Science, March 15, 1901. ↩︎
Stanton, Jour. Geol., Vol. XVII. ↩︎
Kayser, op. cit., p. 286. ↩︎
Merriam, The Thalatlosauria, Cal. Acad, of Sci.; Triassic Ichthyosauria: Memoirs of the Univ. of Cal., 1908. ↩︎