Public domain
[p. 702]
¶ Formations and Physical History
The eastern part of the continent. Jurassic formations have not been identified certainly in the eastern half of the continent, but there are beds outcropping along the western margin of the Coastal Plain in Maryland that may belong to this system. In constitution, structure, and all physical relations their alliance is with the Corn anchean (Lower Cretaceous) of the same region, and since their fossils are indecisive, this seems, at present, their best correlation.
Erosion seems to have been the leading geologic process in the eastern part of the continent during the period. Its effectiveness may be judged by the fact that both the uplifted and deformed Triassic system and the Appalachian mountain region farther west were essentially base-leveled before the Comanchean period was far advanced. The sediments worn away from these areas were deposited somewhere, presumably east of the present coast. Erosion seems to have been in progress also in the eastern interior over most or all the area which emerged during the closing stages of the Paleozoic.
The western interior. In contrast with the eastern half of the continent, deposition was in progress, probably, in some parts of the western interior, though the possible early Jurassic beds of this region have not been clearly differentiated from the Trias. There is perhaps room for doubt whether the early and middle parts of the system have much representation in this region.
Late in the period, an arm of the sea extended itself over a large tract in the western interior (Fig. 475), covering much of Wyoming,[1] [p. 703]Montana,[2] Utah, and Colorado, and parts of several other states[3] (Fig. 476). This is shown by the presence in these states of sedimentary beds containing marine fossils of early Upper Jurassic age. The beds are chiefly exposed in the mountains (Wasatch, Uinta, Black Hills, etc.) where the erosion which followed the uplift and deformation of the strata has discovered their edges.[4]
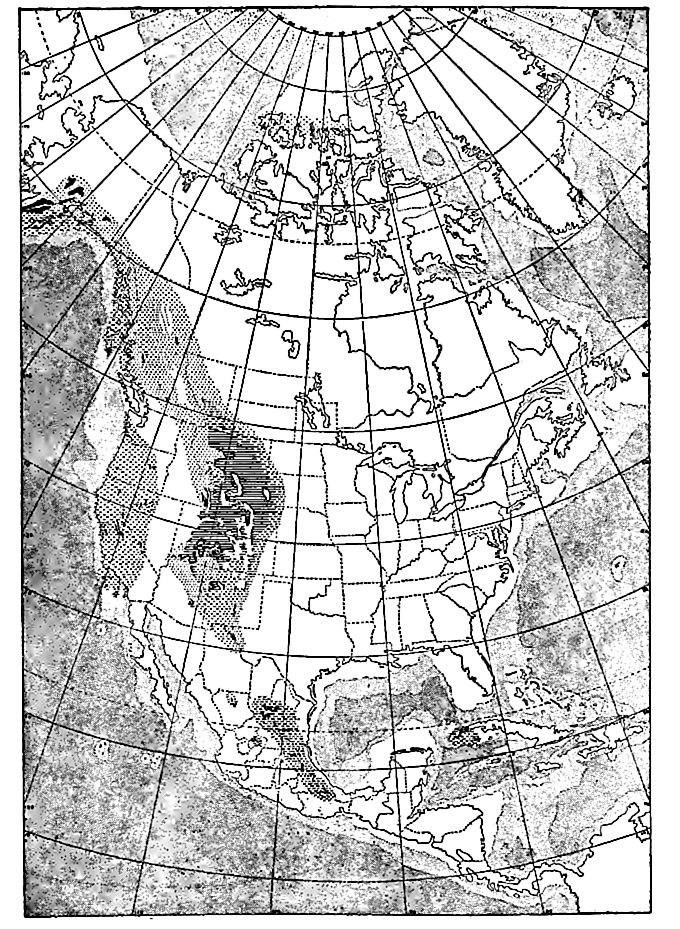
[p. 704]
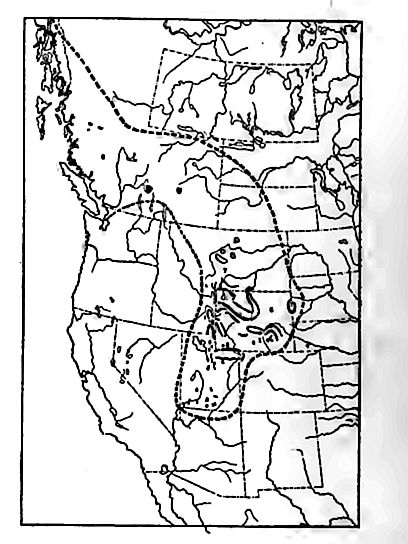
The avenue through which the sen reached the western Ulterior has not been determined, but the fossils of the interior are so unlike those of the Californian coast as to lead to the inference thai the waters of the interior did not come in from the west. The identity [p. 705] of many species from the Upper Jurassic beds of the Queen Charlotte Islands and British Columbia with those of the western interior, imply shallow-water connection of these areas. Whether this connection was through British Columbia direct, or by way of Alaska and down the east side of the Rocky Mountains, is not known. The presence of fresh-water beds (the Morrision [or Como] beds of Colorado, Montana, and Wyoming), sometimes regarded as of late Jurassic age in some parts of the western interior, would, were their age established, show that the sea-water withdrew before the end of the period.

Marine Jurassic limestone has been reported recently from western Texas,[5] and though the exposures are limited, their connections are probably southward with the Jurassic of Mexico, where the system is somewhat wide-spread.
The Pacific coast. Marine deposition was in progress on the Pacific coast[6] (California and Oregon), though much of the system [p. 706] here is concealed beneath younger formations, so that its original extent is not known. In the latitude of Nevada and Utah, the earlier formations of the system extended, east to longitude 117°.
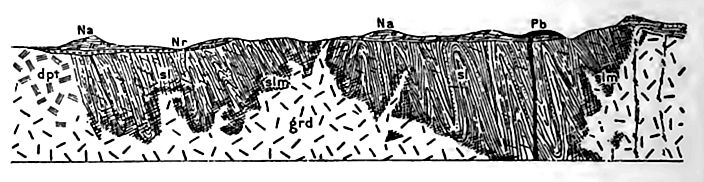
The Lower Jurassic beds generally rest on the Trias conformably, where both are present, but the younger beds overlap the older systems at some points, and fall short of it at others.
In the later pari of the period, the sea appears no1 to have extended east of the Sierras in the latitude of California, but in northern [p. 707] British Columbia, where the Lower and Middle Jurassic beds have little representation, the sea extended farther east than during the earlier part of the period. The deposits embrace all the usual sorts of sedimentary rocks, as well as considerable beds of fragmental igneous rock. Jurassic formations are also known at somewhat widely separated points in Alaska.[7] On the shores of Cook Inlet, 10,000 feet of Middle and Upper Jurassic are reported.
Thickness. The total thickness of the system in California does not exceed 2,000 feet (in part tuff). Farther east, in western Nevada,[8] nearer the land whence sediment was derived, it attains a thickness twice or thrice as great, being made up of limestone below, and slates above. In the western interior, its thickness is relatively slight.
Surface distribution and position of beds. The Jurassic beds do not now appear at the surface over large areas, being concealed in many places by younger beds. In some areas they retain their original position, while in others they have been tilted, or even folded or metamorphosed. This is especially the case in the Sierra Mountains and in some other ranges near the western coast.
¶ Close of the Jurassic
Orogenic movements. At the close of the Jurassic period, there were considerable disturbances in the western part of North America. Great thicknesses of Triassic and Jurassic strata began to be folded into the Sierras,[9] and the Cascade and Klamath[10] Mountains farther north perhaps began their growth. It is not to be understood that these mountains attained great height at this time, or that they have not had later periods of growth. In the Klamath Mountains, for example, there are deformed beds of late Tertiary age. After this closing-Jurassic period of orogenic movement, the coast was [p. 708] somewhat farther west than now in northern California and southern Oregon.
It is probable that the Coast Range of California began its history at the close of this period, for deformed Jurassic beds (Golden Gate series) underlie the Lower Cretaceous unconformably in the axis of the range;[11] but the movements which gave the Coast Range its present form (modified by erosion), took place much later. Various other ranges of the west are thought to have begun their history as mountains at about the same time.
Toward the close of the period, much, if not all, of the great Upper Jurassic gulf of the northwestern part of the continent disappeared. All in all, the deformations at this time were greater than those which mark the close of most periods.
¶ Foreign Jurassic
Europe. Jurassic strata are exposed in many and widely separated parts of Europe, though for the most part in small areas only. As in the case of older systems, the present distribution of the system at the surface is no measure of its real extent.
It has been thought that the Jurassic of England is probably continuous with that of France beneath the English Channel, and thence by way of southeastern France, with those parts of the system which appear about the Mediterranean, and by way of Belgium, the Netherlands, and the German lowlands, with those parts which appear in Poland and Russia. In southern Russia, too, the Jurassic beds are probably wide-spread beneath younger formations. The lower part of the system is less extensive than the Middle, and the Middle less wide-spread than the Upper. Progressive submergence was, indeed, one of the features of the period. In this respect, the North American and European continents are in harmony, btit marine formations are much more extern let 1 in Europe.
In Europe, the subdivision of the system has been carried bo a high degree of refinement. but the many zones are grouped into a few principal divisions:
[p. 709]
| England (4,000-5,000 feet) | Germany (2,000-3,000 feet) |
| Upper (Portland) Oölite | Upper (White) Jura or Malm |
| Middle (Oxford) Oölite | Middle (Brown) Jura or Dogger |
| Lower (Bath) Oölite | |
| Lias | Lower (Black) Jura or Lias |
Among the more distinctive features of the system in Europe are the following:
(1) A considerable content of coal in some places, notably Hungary. (2) The abundance of oolitic limestone, both in England and on the continent. (3) The presence of lithographic stone (Solenhofen limestone of southern Germany) . This stone is so fine and so even-grained, and at the same time so workable and so strong, that it has come into use the world over for lithographic purposes. The stone is also remarkable for the perfection of its fossils, including such delicate parts as the gauzy wings of insects. (4) The considerable development of non-marine beds in the lower part of the system, and again at its very top, especially outside of southern Europe.
Close of the Jurassic in Europe. The close of the Jurassic appears to have been marked by a somewhat wide-spread emergence of land. In central Europe, the emergence appears to have begun before the close of the Jurassic, for the latest beds (Purbeck) of the system in England are unconformable on beds lower in the system. Similar changes are known to have occurred in late Jurassic time in some other regions. On the other hand, the Upper Jurassic and the Lower Cretaceous beds are in places so closely associated as to show that no change of continental dimensions brought the Jurassic period to a close. Great deformative movements seem to have affected no part of Europe at the close of the period.
Extra-European Jurassic
The Upper Jurassic is wide-spread in Arctic lands. This distribution points to a great Arctic sea in the later part of the period, with two considerable dependencies to the south, — the one in Russia, the other, as we have seen, in western America. The Lower Jura is wanting in these latitudes, so far as known, and the Middle Jura is present but rarely.
[p. 710]
The lower Jura occurs in southwestern Asia and Japan. The Middle Jura, largely clastic and of terrestrial origin, is wide-spread in Northern Asia, and marine in Middle Jura is known in northern India. The Upper Jura is much more extended, especially in the north. The system is known in New Zealand, Borneo, and Australia, and is well developed in Mexico, Peru, the Bolivian Andes, Chile, and Argentina.
Coal. Coal of considerable value is somewhat widely distributed in the Jurassic system. Besides that in the Lias of Hungary, coal occurs in the Caucasian region, Persia, Turkestan, southern Siberia, China, Japan, and Farther India, in many of the islands southeast of Asia, and in Australia and New Zealand. In the last-named island, the coal-bearing formations are interbedded with marine strata, suggesting considerable oscillations of level. In most of these countries, the coal is Liassic. Outside of North America, it is probably that no other system except the Pennsylvanian contains so much coal.
Climate
The testimony of fossils gathered in various parts of the world is to the effect that the climate of the Jurassic period was genial. In Europe, corals lived 3,000 miles north of their present limit, and saurians and ammonites flourished within the Arctic circle. Nevertheless, climatic zones were probably defined. Corals are unknown in the deposits of the great Arctic belt of Upper Jura, and the detailed study of the Jurassic faunas has led to the belief that one climatic zone is recorded in the Jurassic beds of the Arctic belt, a second in the deposits of central Europe, and a third in the southern province of Europe and the lands farther south. There can be no doubt of the differences in the faunas of these different provinces, but it is not certain that the differences were due wholly or even mainly, to climatic influences.
¶ The Life of the Period
As the Jurassic was a period of sea extension, the marine life again assumes a place of leading importance. At the same time the land life, though suffering somewhat by the limitation of its [p. 711] habitat, was favored by the genial climate. The frequent shif tings of land- and sea-areas, without involving great relief or severe climatic states, favored biological changes.
The Marine Life
It will be recalled that an expansional stage of epicontinental sea life set in toward the close of the Trias. This continued into the Jurassic, reaching a climax late in the period, when the sea attained the limit of its transgression over the land. The faunal progress is less well revealed in North America than in Europe and Asia, and the following general sketch of the life is based, in part, on the fossils of those continents.
The great features of the marine life of the period lay in (1) the continued dominance of ammonites among the invertebrates, (2) the rise of the belemnites, (3) the abundance and modernization of pelecypods, (4) the rejuvenation of corals and crinoids, (5) the marked development of sea-urchins, (6) the introduction of crabs and modern types of crustaceans, (7) the prevalence of foraminifera, radiolarians, and sponges, (8) the change in the aspect of the fishes, and (9) the great sea-serpents, descended from the land-reptiles of the Trias.
(1) The ammonites were still the masters among invertebrates, and were represented by many beautiful forms (Fig 481). They deployed along ascending lines in some cases, and retrograde lines in others. Erratic and degenerate developments showed themselves by uncoiling and strange coiling, presaging a stage of “sporting” and retrogression in the next period, followed by extinction. Despite these adverse foreshadowings, the ammonities were yet in the heyday of their luxuriance and beauty.
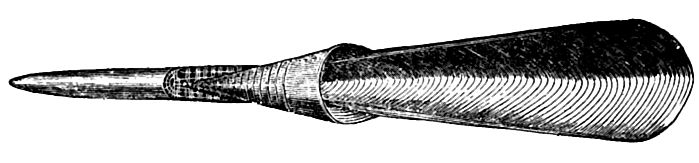
(2) Ammonites and their predecessors (ceratites, goniatites, and orthoceratites) were tetrabranchs, with external shells; but dibranchs, with internal shells, had appeared in the Trias, and rose rapidly to prominence in the form of belemnites, usually represented in the fossil state by their internal shell or “pen” (Fig. 482). In the course of the period the belemnites almost came to rival the ammonites, and were almost as characteristic of the successive [p. 712] stages of deposition. The first known cuttle-fishes (sepeoids), also appeared at this time.
(3) Pelecypods flourished duriagthe period (Fig. 483), and took on a markedly modern aspect, the oyster family taking the fead. Gastropods were abundant in some quarters, but singularly absent in others. Existing genera were represented.

[p. 713]
(4) Suggestive of shallow clear seas was the reappearance of corals and crinoids in abundance in the later part of the period. The modern (Hexacoralla) type of corals was in the ascendant (a, 6, c,Fig. 484) , and formed abundant reefs, particularly in the European seas of the Middle Oolitic stage. Crinoids also rose again to prominence, though their diversity was not great. They departed from Paleozoic forms in various ways. Most of them lived in shallowwater, as most of the Paleozoic types had; but there is evidence that deep-water species had begun to appear, leading toward the prevalent but not exclusive habit of the present.

[p. 714]

[p. 715]
(5) The slow evolution of the sea-urchins in the Paleozoic era was succeeded in the late Trias by the beginning of a rapid evolution, which reached its climax in the early Tertiary.
(6) The Paleozoic crustaceans, the trilobites in the sea, and the eurypterids in the land waters, had been succeeded by the decapods which rose to a moderate and prolonged ascendancy. The prawns and lobsters (Macrura, long-tailed decapods) were the earlier division, and the most numerous in this period; but the first of the known crabs (Brachyura, short-tailed decapods) appeared before the period was past. The macrurans seem to have frequented embayments and protected locations near the land, or perhaps within it, where terrestrial, fresh-water, and marine species are preserved in the same sediments. It is not improbable that the macrurans, then as now, had representatives in terrestrial waters, as well as in the sea.

[p. 716]
(7) Sponges and foraminifera were prolific and arc well preserved.
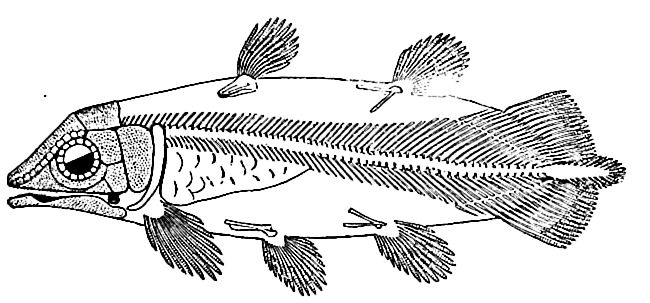
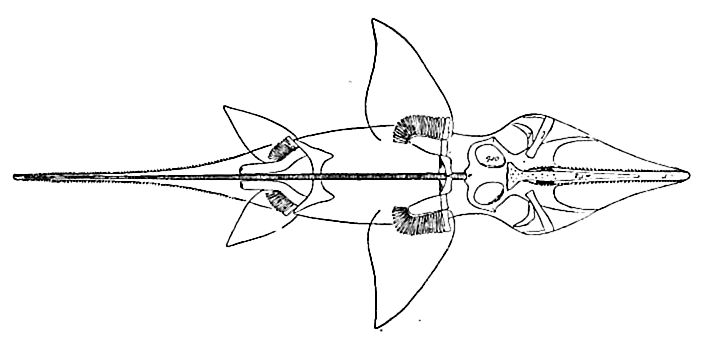
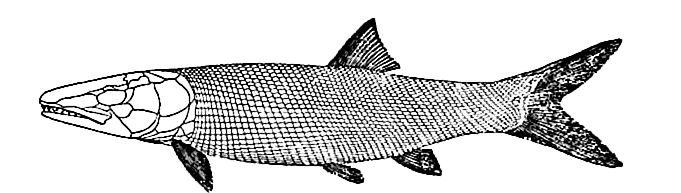
(8) A marked change in the aspect of the fishes had set in during the Trias, and was carried farther in this period. The crossopterygians (Fig. 486) and dipnoans were reduced, the selachians continued undiminished; the skates and rays began their modern career; the Chimceridae, the existing family of sea-cats or spookfishes, made its appearance, so far as fossils show, and developed notably (Fig. 487); the forebears of the living garpikes and sturgeons took precedence in numbers; the forerunners of the modern Amia (Fig. 488) were important, and the initial forms of the bony fishes (teleosts), the dominant existing type, made their appearance. The aspect of the class was markedly more modern than at the close of the Paleozoic.
(9) It was noted under the Trias that certain land-reptiles went down to sea, and introduced a new phase of vertebrate mastery over the deep. Though doubtless suffering from the new dynasty, it appears that the fishes continued in notable abundance and variety. It will be seen later that they outlived the invading race, and resumed, in large measure, their former dominance. Some of the reptiles which had taken to the sea had become extinct, while others made their first appearance late in the period. The ichthyosaurs (fish-like saurians) reached their highest development in this period, and seem to have swum every sea. Their adaptation to aquatic life is shown in the complete transformation of their limbs into paddles (Fig. 489), in the reduction of the outline of the body to ichthic lines and proportions, in the sharp down-bending of the vertebrae at the end of the tail for the support of a caudal fin, in the long snout set with teeth adapted to seize and hold slipping prey, but not to masticate it, in the protection of the eye by bony plates, and, interestingly enough, in the development of a viviparous habit that freed them from the necessity of returning to land to deposit their eggs, after the manner of sea-going turtles and crocodiles. That their food consist ed in part of invert eJ urates US evident from the fossil contents of the stomachs, the remains of 200 beleinnites having been found in a single one. There were small as well as large forms of ichthyosaurs some exceeding 30 feet in length.
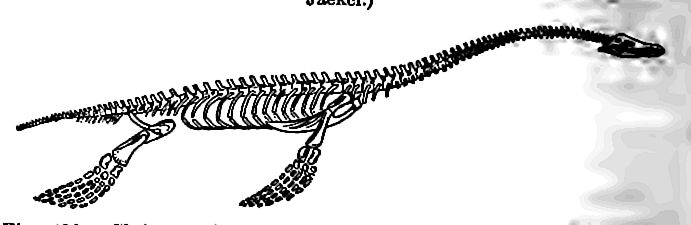
Descended from a different stock, the plesiosaurs adapted them selves to sea life in another way (Fig. 490). The body took on a form like that of a turtle, while the neck was elongate, giving rise to the epigrammatic description “the body of a turtle strung on a snake.” Locomotion seems to have been chiefly dependent on the paddles, though a fin-like adaptation of the tail existed in some. The elongation of the neck was variable, the vertebrae of the neck ranging from 13 to 76. The neck appears not to have been so flexible as familiar illustrations have represented it, nor were the jaws separable and extensible as in the case of snakes. This implies either that they lived on small prey, or tore their food to pieces before swallowing. They were doubtless formidable foes of the smaller sea animals, but probably not of the larger. Like the ichthyosaurs, they were without scales. They ranged from 8 to 40 or more feet in length.
[p. 717]
[p. 718]


[p. 719]
Marine crocodilians made their appearance in the later part of the period. They had undergone a remarkable adaptation to the sea (Fig. 491). They were fish-like in appearance, their skins were bare, and their tails terminated in a large fin like that of the ichthyosaurs. The fore limbs were short and paddle-like. The hind limbs were modified but slightly from the land type, perhaps due to the recurring necessity of visiting the shores for depositing and hatching their eggs.
Marine turtles, so characteristic of the Cretaceous, had not yet appeared.
The Land Life
Vegetation. The land vegetation of the Jurassic was little more than a continuation and expansion of that of the late Triassic, with slow progress toward living types. Cycadeans, conifers, ferns, and equiseta were the leading plants, slightly more modernized than their Triassic ancestors, but not changed radically.[12] The cycadeans (Bennettitales and Cycadales) were perhaps the most distinctive forms, though the conifers showed the more notable modernization. They embraced yews, cypresses, arborvitas, and pines, all of which had a somewhat modern aspect, though all the species are extinct. The ginkgos also played a somewhat important role.
An interesting feature of the European record is the rather frequent occurrence of land plants in marine beds, which implies that many trunks, twigs, leaves, and fruits, were floated out to sea, and that the landward edges of the marine deposits have escaped [p. 720] destruction. In the same beds are the remains of many land insects, not a few of them being wood-eating beetles.
In the closing stages of the period, the area of land was extent led. This should in itself have been favorable to an expansional development of plants; but extensions of land are so liable to be attended by adverse climatic and topographic changes, that no safe inference can be drawn except from the actual record, which, in this case, is rather scanty.
Animals. Of the North American land faunas of the Jurassic, except perhaps its latest stages, little is known. But with the close of the period there appeared in the Morrison beds (p. 705) a rich but not varied land fauna, composed chiefly of dinosaurs. These animals attained remarkable size and diversity, and were easily lords of the reptile horde. Some were large, and some were small, and the group, as a whole, developed great diversity in many directions. There were not only carnivorous types, which had appeared in the Triassic, but numerous herbivorous forms. Among them all there was not a single type which was distinctively North American. It is therefore concluded that there was freedom of migration between the eastern and western continents at this time.
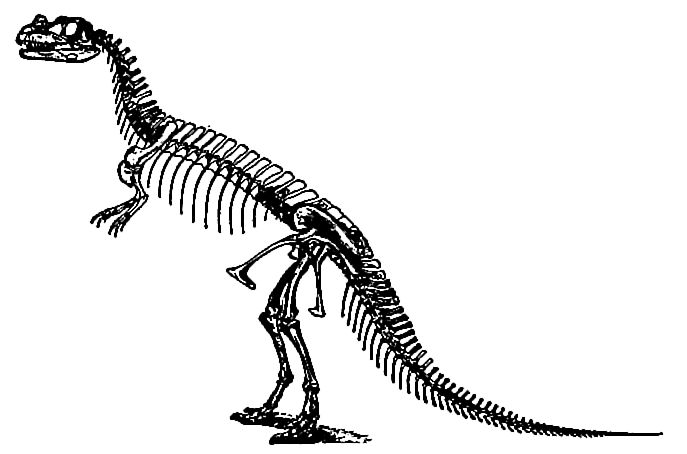
Of the carnivores, one of the most typical was Ceratosaurus nasicornis (Fig. 492) . The fore limbs seem to have been used chiefly for seizing and holding prey, rarely for walking. The animal’s pose was facilitated by hollow bones. The head was relatively large, an unusual character for a race among which small heads and brains were the fashion of the time. Not all the carnivorous dinosaurs were large. There were small leaping forms (like Compxognoihus) not larger than a rabbit.
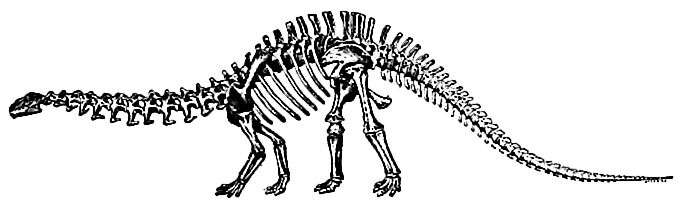
The herbivorous dinosaurs are known first in this system, but their development was so extraordinary that they soon outranked the carnivorous forms in both size and diversity. The sauropoda were generally massive, with sub-equal limbs and the quadruped habit. Among them, Brontosaurus (Apatosaurus) att ained the extraordinary length of 60 feet and possibly more, taking rank as one of the largest of known land animals (Fig. 493) . This enormous creature was characterized by weakness rather than strength, for its general organization was unwieldy, its head small, and its [p. 721] brain had less diameter than its spinal cord. “The task of providing food for so large a body must have been a severe tax on so small a head.” The largest of all known dinosaurs was Brachiosaurus, of which the femur measured more than two meters in length (80½ inches)[13]. There were several other genera of similar nature, and of bulk only inferior to these monsters.
The typical ornithopod (bird-footed) dinosaurs were bipedal in habit, like the carnivores. On the hind limbs there were usually [p. 722] only three functional toes, so that they left a bird-like track; the fore limbs, however, had five digits. Camptosaurus, one of the largest of this group, measured about 30 feet in length, and 18 in height in the walking posture.
The stegosaurs were quadrupedal in habit, and had solid bones. They were curiously armored, and formed a very remarkable group that frequented England and Western America. They were not so large as some of the preceding, but found compensation in protective plates, spines, and similar modes of defense. The Stegosaurus of Colorado and Wyoming (Morrison beds) was one of the most unique (Fig. 494). Its diminutive head and 1 train imply a sluggish, stupid beast, depending for protection on its bulk and armor.

Turtles, which had lived elsewhere since the Middle Trias, made their first appearance in North America in the Morrison beds, and the crocodilians of the period, though still retaining the primitive [p. 723] type of biconcave vertebrae, became differentiated into several branches. Primitive lizards were doubtless abundant, but because of their terrestrial habits and small size, they have little representationamong the fossils, and none have been found in our continent. It has already been noted that crowding on the land may have led some land reptiles to take to the sea. The same influence may have forced others to take to the air, thereby escaping the monsters of the swamps, jungles, and forests. Whatever the cause, a unique feature of the period was the development of pterosaurs, or flying reptiles. Appearing at the very close of the Trias in a few yet imperfectly known forms, they presented themselves at the opening of the Jurassic period as fully developed flying animals (Dimorphodon), and later formed a diversified group embracing long-tailed (as Rhamphorhynchus, Fig. 495) and short-tailed forms (as Pterodactylus, Fig. 496). With little doubt they sprang from some agile, hollowboned saurian, more or less remotely akin to the slender, leaping dinosaurs. Between the ponderous brontosaurs (Fig. 493) and the pterosaurs (Fig. 495), the Jurassic saurians present the strangest contrasts.

The Jurassic pterosaurs were small, but their successors attained a wing-spread of nearly a score of feet. They were curiously composite in structure and adaptation. Their bones were hollow, their fore limbs modified for flight, their heads bird-like, and their jaws set with teeth, though toothless forms appeared later. They were [p. 724] provided with membranes stretched, bat-like, from the fore limbs to the body and hind limbs, and serving as organs of flight (Fig. 495). The fifth, or as some paleontologists believe, the fourth, digit was greatly extended, and served as the chief support for the wingmembrane. The sternum was greatly developed, implying true powers of flight, a conclusion supported by the occurrence of their remains in marine sediments free from other land fossils. Some of them had singular elongate rod-like tails, with a rudder-like expansion at the end.
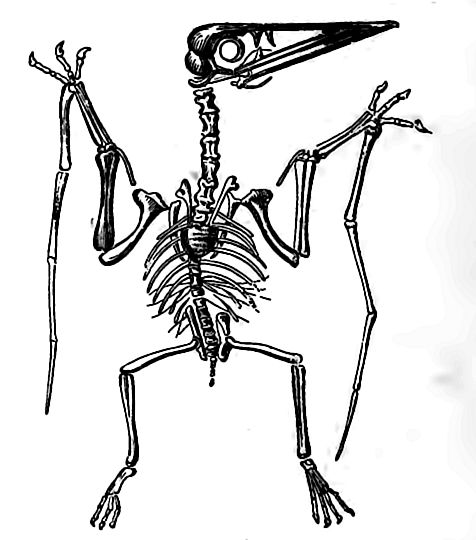
The pterodactyls (Fig. 496) had short tails, and were usually small and slender. Fully differentiated as first found, the pterosaurs underwent no radical change of structure during their career, and the steps of their remarkable evolution are U)- the most part unknown. Representations of flying reptiles among the .Jurassic fossils of North America are extremely rare.
[p. 725]
A less bizarre, but really greater evolution, was the contemporaneous differentiation of true birds. The remote ancestors of the pterosaurs and the birds may have been closely allied, but there is no evidence that the birds are descended from pterosaurs. The two types are examples of analogous and parallel evolution, not of relationship.
[p. 726]
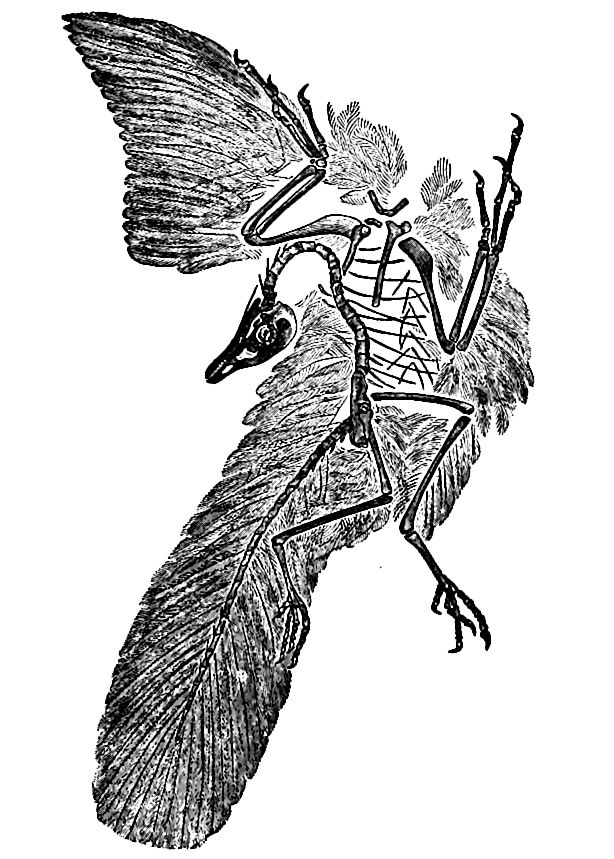
The oldest known bird, Archceopteryx macrura (Fig. 497), shows an advanced state of evolution, and at the same time clear traces of reptilian ancestry. From this ancestry it retained a long, vertebrated tail, reptile-like claws, fore limbs, teeth set in sockets, biconcave vertebrae, and separate pelvic bones. On the other hand, its head and brain were bird-like, its anterior limbs adapted to flying in bird- (not pterosaurian) fashion, its posterior limbs modified for bird-like walking, and most distinctive of all, it was clothed with feathers. The development of the feathers, while yet the body retained so many reptilian features, is most notable. But for their fortunate preservation, it is uncertain whether the creature would have been classed as a bird or reptile. The known specimen was somewhat below the size of a crow.
The marvelous deployment of aquatic and terrestrial reptiles and of birds makes the scanty record of the mammals all the more singular. Only a few jaw bones of the size of those of mice and rats have been found. These low types are referred, without complete certainty, to marsupials. They appear to have been insectivorous.
The insects of the Jurassic appear to have included members of nearly all the fossilizable groups that were not dependent on angiospermous plants.
Map work. Folios published by the U. S. Geological Survey, good for the study of the Jurassic and Triassic systems, are the following: California, Downieville, Jackson, Pyramid Peak, Redding, San Luis; Colorado, Anthracite-Crested Butte, Rico, Telluride, Tenmile, Walsenburg; Massachusetts-Connecticut, Holyoke; Montana, Fort Benton; New Jersey, Passaic; New York-New Jersey, New York City; Oregon, Roseburg; South Dakota- Nebraska, Oelrichs; Virginia-West Virginia- Maryland, Harpers Ferry; Wyoming, Alladin, Cloud Peak-Fort McKinney, Devils Tower.
These folios may well be grouped geographically for study. Thus those which deal with the Newark series of the east form one unit, those of the Rocky Mountains another, those of the Pacific coast another.
In the folios, the Jurassic and Triassic are commonly grouped together under the name Jura-Trias, but the text of the folios often separates the Jurassic from the Triassic, even where the map does not.
¶ Notes
Logan, Jour. Geol., Vol. VIII, p. 241; Knight, Bull. 45, Wyo. Exp. Station, and Bull. Geol. Soc. of Am., Vol. XI, pp. 377-388, and the Wyoming folios of the U. S. Geol. Surv. ↩︎
See Montana folios, U. S. Geol. Surv. ↩︎
For South Dakota see Darton, 21st Ann. Rept., U.S.Geol Surv., Pt. IV. and the folios, U. S. Geol. Surv. ↩︎
In addition to the above folios, U. S. Geol. Surv., see those of Colorado. ↩︎
Cragin, Jour. Geol., Vol. V; and Hill, Am. Jour. Sci., Vol. II, 1897, p. 449, and Physical Geography of Texas, Topographic Atlas, U. S. Geol. Surv. ↩︎
For the Jurassic of the Pacific coast, see Hyatt, Bull. Geol. Soc. of Am., Vols. Ill and V, both articles chiefly paleontological; Meek, Paleontology of California, Vol. I, and the California folios of the U. S. Geol. Surv. ↩︎
See Alaskan Reports, U. S. Geol. Surv. ↩︎
King, Survey of the 40th Parallel, Vol. I. ↩︎
Whitney, Geology of California, Vol. I, and Am. Jour. Sci., Vol. XXXVIII, 1864; and Fairbanks, Am. Geol., Vol. IX, 1892, Vol. XI, 1893. ↩︎
Diller, Bull. Geol. Soc. Am., Vol. IV, p. 224, and 14th Ann. Rept., U. S. Geol. Surv. ↩︎
Fairhanks. Jour. Oeol., Vol. Ill, pp. il."> K50, and Smith, Bull, OeoL Soc. of Am., Vol. V, pp. 257-258. ↩︎
For a comprehensive paper on the Jurassic plants of the United States, with descriptions and illustrations by Lester F. Ward, see 20th Ann. Rept., U. S. Geol. Surv., 1898-99, pp. 334-430. ↩︎
Riggs, Amer. Jour. Sci., 1903. ↩︎