| XXI. El Silúrico y los primeros animales que respiraban aire | Página de título | XXIII. El auge de los peces y la profecía del dominio de los vertebrados |
[ p. 282 ]
Descripción general. — En los mares y océanos actuales poco profundos, y más especialmente en los de aguas cálidas, existe una gran variedad de animales radialmente simétricos, con o sin esqueletos externos de materiales homófobos o calcáreos. Estos animales con forma de flor son los corales y las anémonas, conocidos técnicamente como Caelenterata. Este nombre implica que tienen forma de saco, con una sola cavidad interna que sirve tanto de cavidad corporal (celoma, característico de todos los animales superiores) como de saco digestivo (enterón). Esta simple estructura corporal, comparable a un saco de doble pared, tiene el extremo bucal deshilachado en tentáculos, mientras que el otro extremo está adherido a objetos extraños en el fondo marino.
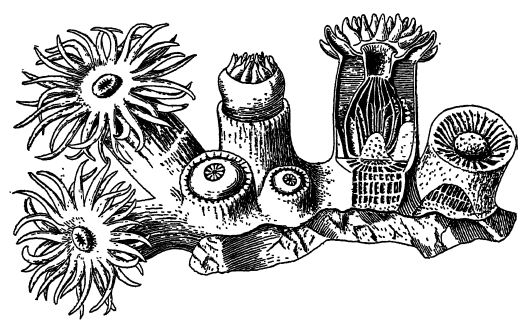
Los celentéreos son, por regla general, animales muy simples en los que aparecen los inicios de órganos definidos. En su variedad, parecen [ p. 283 ] casi agotar las posibilidades de la simetría radial, y muchos son muy similares a las flores. Por ello, también se les ha llamado zoófitos (animales vegetales), y sus colonias de yemas ofrecen interesantes ejemplos de cooperación y división del trabajo.
Algunos, como las anémonas, no secretan un esqueleto, pero muchos otros tipos sí lo hacen, y a todos ellos se les llama comúnmente corales pétreos, pero no todos los esqueletos calcáreos parecidos a los corales son de ninguna manera obra de verdaderos corales.

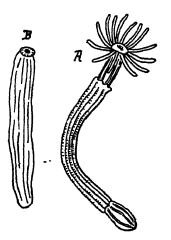
Los celentéreos pican mortalmente a los animales más grandes de los que se alimentan. Esta picadura se produce mediante una multitud de diminutos filamentos, cada uno de los cuales sale disparado de una célula enterrada en la piel y penetra en cualquier cuerpo blando que entre en contacto con los animales (Fig., lado opuesto).
El alimento pasa a la boca, que se digiere en una cavidad del saco de estructura más o menos sencilla, y por la misma abertura se expulsa cualquier resto indigerible. Se alimentan, por regla general, de animales más pequeños, pero cuando los individuos, o pólipos, como se les llama, son diminutos, su alimento consiste en animales o plantas microscópicos.
¶ Antozoos
Existen muchos tipos de celentéreos, pero en este capítulo nos centraremos únicamente en los más complejos: los antozoos (que significan animales flor-flor) formadores de rocas. Estos son los corales pétreos, muchos de los cuales forman arrecifes y rocas. Los animales que construyen los arrecifes pétreos suelen ser muy pequeños (menos de 6 mm), aunque algunas especies alcanzan* un diámetro de 2,5 cm o más. Los esqueletos coloniales pueden alcanzar hasta 4,5 metros de ancho y 4,5 metros de alto.
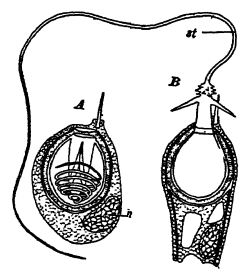
La cavidad interna presenta un esófago digestivo tubular rudimentario que se abre hacia arriba en una boca alargada y, por debajo, en la cavidad digestiva verdadera o codenterón (véase la fig., [ p. 284 ] pág. 282). La pared de este último presenta varios pliegues longitudinales o crecimientos internos de la pared corporal que se proyectan con mayor o menor profundidad en la cavidad. Al cortar transversalmente uno de estos pólipos, los pliegues o particiones se observan como radios, por lo que también se les conoce como animales rayados (fig., pág. 282). Cuando existe un esqueleto de soporte duro o parte pétrea, conocido como corallum, como suele ser el caso, este suele estar compuesto de carbonato de calcio (véase la parte I, pág. 182), casi siempre es secretado por la capa externa del cuerpo blando y tiene su forma exacta. Por lo tanto, los esqueletos de los corales tienen una estructura radiada o particionada, y es solo este esqueleto calcáreo el que se conserva en los fósiles (véase la fig., abajo). Los pólipos viven solos y luego tienen forma cilíndrica o cónica (los corales copa), o más comúnmente forman colonias mediante la gemación de individuos muy adyacentes.

El esqueleto del coral. — Los corales copa, que son las bases calcáreas de pólipos individuales, son muy frecuentes en las rocas paleozoicas. Se multiplican por reproducción sexual y no por gemación, como ocurre en las formas coloniales. Los corales copa suelen ser cónicos y más o menos curvados, con copas de profundidad variable divididas en numerosos compartimentos poco profundos por tabiques radiales llamados septos (véase la figura anterior). Estos septos se originan en la pared exterior y tienen una longitud variable, con una disposición regular; solo una parte de ellos, los septos primarios por lo general, se unen en el centro, donde a menudo forman una [ p. 285 ] columna simple o retorcida, la columela. Los corales coloniales tienen aproximadamente la misma estructura general que los pólipos individuales, pero cuando están muy juntos, los coralitos adquieren una forma prismática (véase la figura, pág. 284). También pueden tener un contorno circular y los espacios entre los individuos se rellenan con una estructura laxa.
Los tipos de estructura de coral que se observan con mayor frecuencia como fósiles son los siguientes:
Tabulata o corales tabulados (tabula significa mesa y hace referencia a las numerosas particiones transversales que se encuentran en los tubos). Estos corales paleozoicos primitivos pero especializados siempre crecen en forma colonial y nunca como individuos únicos, por lo que entre los Tabulata no hay corales de copa. Como regla general, los pólipos individuales son pequeños o incluso diminutos transversalmente, pero longitudinalmente forman tubos más o menos largos que están abundantemente divididos por tabulsas y perforados por poros a medida que los animales crecen hacia arriba (véase la figura anterior). Los pólipos pueden tener un crecimiento laxo cuando los coralitos individuales son redondos u ovalados en contorno, pero con mayor frecuencia los tubos están estrechamente adheridos entre sí y los coralitos son uniformemente prismáticos. No hay septos radiales y, cuando parecen estar presentes, se ven como una serie de espinas cortas dispuestas en filas longitudinales (septos incipientes, por así decirlo), o las espinas pueden estar dispuestas irregularmente. Por lo tanto, los Tabulata son corales paleozoicos sin septos verdaderos, por lo que se distinguen fácilmente de sus asociados, los Tetracoralla. Las paredes de los coralitos son delgadas y están perforadas por grandes poros, los poros murales (véase la Fig. B, arriba), que representan fallas en la gemación debido a la escasa disponibilidad de espacio para que la gran mayoría de los posibles brotes se conviertan en individuos.
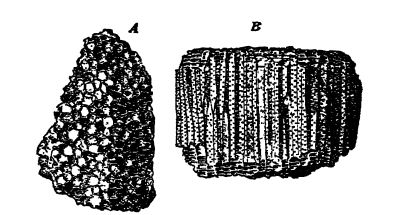
Los corales tabulados aparecieron en el Champlainiano Medio y alcanzaron su máximo desarrollo en el Silúrico y el Devónico. El coral cadena [ p. 286 ] (Halysites, Lám., pág. 270, Fig. 1) fue característico del Silúrico y el Champlainiano, mientras que los corales panal (Favosites, Lám., pág. 285) fueron importantes formadores de arrecifes en el Silúrico y el Devónico, donde algunas colonias alcanzaron un diámetro de 1,2 metros. El coral organpipe (Syringopora, Lám., pág. 320, Fig. 8) fue común durante todo el Silúrico y la mayor parte del Paleozoico tardío.
Tetracorales (corales con los septos dispuestos en cuatro cuadrantes o bimdles). — A lo largo del Paleozoico desde el Champlainiano Medio, y más especialmente a partir del Silúrico, los tetracorales son fósiles comunes (véanse las figuras, pág. 284, y 4-7, pág. 320). Se presentan como coralitos individuales, los corales copa, o en colonias compuestas. Los corales copa fueron muy comunes en el Paleozoico Medio y fueron más numerosos, más variados y de mayor tamaño (hasta 60 cm de largo) que los de épocas posteriores. Los tetracorales compuestos también fueron abundantes y, junto con los corales copa, tuvieron una amplia distribución, siendo especialmente comunes como formadores de arrecifes en el Silmiano y el Devónico. Fueron raros en el Pensilvánico y desaparecieron con el Pérmico.
En los tetracorales de copa, la formación de septos suele apreciarse fácilmente, y con su cubierta exterior lisa o ligeramente rugosa (epiteca) que se extiende desde la base hasta el borde de la copa, se distinguen fácilmente de otras especies. Los tetracorales compuestos no suelen distinguirse tan fácilmente de los hexacorales (corales de seis radios), debido a que el tetramerismo o división en cuatro partes está muy o totalmente oculto. Sin embargo, por regla general, se observa que los pólipos son más grandes, mientras que los septos radiales son más numerosos o, al menos, más delgados, rectos y simples. Algunos de estos tetracorales compuestos alcanzan el tamaño de colonias modernas y se han observado hasta 3,6 metros de diámetro.
El zoantario más primitivo en los mares actuales es Edwardia, y se sabe que las Hexacoralla actuales atraviesan una etapa similar a Edwardia en su ontogenia. Edwardia es un animal de cuerpo blando, es decir, carente de esqueleto, que vive en aguas más frías. En el Cámbrico Medio, cerca de Field, Columbia Británica, Walcott encontró un fósil muy similar a Edwardia, también sin esqueleto calcáreo. Lo denominó Mackenzia costalis, considerándolo, sin embargo, como una holoturia. Raymond sugiere que Mackenzia podría estar estrechamente relacionada [ p. 287 ] con Edwardia, y que formas similares podrían haber dado origen en el Champlainiano a los Tetracoralla, que secretan cal, y posteriormente, en el Triásico, a los corales pétreos o Hexacoralla. (Véase la Fig., pág. 256).
Los hexacorales fueron las especies comunes después del Paleozoico y son los principales constructores de arrecifes de los océanos actuales. El nombre hace referencia a que los pólipos jóvenes comienzan con seis septos primarios y todos los ciclos subsiguientes de particiones se introducen regularmente entre los precarios. Por lo tanto, existe una disposición radial regular de los septos y nunca se agrupan en cuadrantes como en los tetracorales. Los pólipos individuales tampoco tienen una forma tan marcada de copa o embudo. En las formas compuestas, los animales tienden a ser pequeños, a menudo muy pequeños, como en los corales cuerno de ciervo, mientras que los coralitos no suelen tener paredes tan marcadas entre sí como los corales más antiguos. Las paredes también suelen parecer gruesas, y en muchas formas los individuos están comprimidos y parecen fusionarse irregularmente, lo que sugiere las circunvoluciones del cerebro de un mamífero; de ahí el nombre de corales cerebro, que a veces se les aplica. Véase Parte I, pág. 183.
Hábitats de los corales vivos. — Los hexacorales viven en todos los océanos desde el nivel del mar hasta los 11.000 pies, pero son particularmente comunes en aguas tropicales claras y en profundidades de menos de 240 pies. En las aguas más profundas, oscuras y frías predominan los pólipos simples, mientras que en los arrecifes donde la luz solar es fuerte los corales compuestos son los más prolíficos. Hay cinco veces más tipos de hexacorales en aguas de no más de 150 pies de profundidad que entre esta profundidad y los 240 pies, la profundidad máxima a la que se extiende cualquier especie de arrecife. Entre los 600 y los 2400 pies hay otra zona de coral, generalmente de pequeños pólipos simples o de delicadas formas compuestas ramificadas. Más allá de esta última profundidad, los corales son pocos en número y variedad y los que existen son pequeños y extremadamente frágiles. Frente a la costa este de Norteamérica, entre Cabo Hatteras y Terranova, existen catorce especies de corales, varias de las cuales se extienden por debajo de los 1825 metros (Verrill). Se sabe que los corales hemisféricos alcanzan los 4,5 metros de diámetro, mientras que se ha informado de colonias de corales cuerno de ciervo que alcanzan alturas de 4,5 metros.
Respecto a la temperatura, los corales estrictamente constructores de arrecifes prosperan sólo entre 68° y 78° F, pero continúan viviendo incluso a 85° F. La zona de coral más profunda tiene una temperatura mucho más baja, entre 40° y 50° F, y las especies de aguas profundas pueden vivir en aguas tan frías como 32° F. En otras palabras, los corales pueden vivir a cualquier profundidad del agua del mar, pero los corales constructores de arrecifes sólo viven en aguas cálidas y poco profundas.
[ p. 288 ]
Probablemente existan más arrecifes de coral hoy que en la mayoría de los momentos de la historia de la Tierra, aunque ahora todos se limitan a los océanos más cálidos. Se estima que aproximadamente una vigésima parte de los mares de plataforma (500.000 millas cuadradas) está actualmente cubierta de arrecifes de coral. Este exceso parece deberse a la reciente elevación del nivel oceánico como consecuencia del aumento de agua resultante del derretimiento de los casquetes polares. Gracias a la elevación del nivel oceánico, las zonas inferiores de los arrecifes de coral se convierten en hábitats más favorables y ampliados. Para obtener información sobre la naturaleza de los arrecifes de coral y sus lugares de crecimiento, véanse las páginas 182-190 de la Parte I. Para arrecifes de coral fósiles, véanse los capítulos Silúrico y Devónico.
¶ Lectura colateral
J. D. Dana, Corales e islas de coral. 1872.
W. Saville-Kent, La Gran Barrera de Coral de Australia. 1893.
T. W. Vaughan, Algunos corales de aguas poco profundas de la isla Murray (Australia), las islas Coeos-Keeling y la isla Fanning. Artículos del Departamento de Biología Marina, Institución Carnegie de Washington, vol. 9, 1918, págs. 51-210. (Véase para ver excelentes fotografías de corales vivos y la lista de otros artículos sobre corales del mismo autor).
Corales paleozoicos:
W. J. Davis, Corales fósiles de Kentucky. Servicio Geológico de Kentucky, 1876. James Hall, Fósiles y corales del Devónico. Servicio Geológico de Nueva York, 1876. También en Paleontología de Nueva York, vol. 6, 1887.
P. E. Raymond, Historia de los corales y los océanos sin cal. American Journal of Science, 5.ª serie, Volumen 2, 1921, págs. 343-347.
Cabl Romingeb, Corales fósiles. Servicio Geológico de Michigan, Vol. 3, 1876.
| XXI. El Silúrico y los primeros animales que respiraban aire | Página de título | XXIII. El auge de los peces y la profecía del dominio de los vertebrados |