| XXII. Corales y animales similares a los corales | Página de título | XXIV. El Devónico y el predominio de los peces |
[ p. 289 ]
En este capítulo estudiaremos los peces, sus especies, su ascendencia y cómo sus descendientes, más versátiles, abandonaron sus hábitats acuáticos ancestrales para instalarse en la tierra. Es un estudio fascinante, ya que a toda la humanidad le interesa el gran valor económico de los peces o las horas felices que su pesca proporciona al pescador, pero tiene un interés especial para el filósofo que ve en los peces un paso en el camino de la evolución que lleva desde los invertebrados vermiformes hasta la cima de la independencia orgánica en el hombre racional.
Los peces fueron la primera clase de vertebrados con un éxito notable. Gracias a su columna vertebral y sistema nervioso central, y a su mayor capacidad de alerta, movilidad y fecundidad, tienen una clara superioridad sobre sus especies inferiores, los invertebrados. Están tan adaptados al agua como las aves al aire. Por regla general, viven exclusivamente en mares y océanos o en aguas dulces terrestres. Sin embargo, muchos peces marinos pasan parte de su vida en agua dulce; algunos se alimentan y se reproducen en hábitats tanto dulces como marinos; otros se alimentan en el mar y se reproducen en los ríos, o en casos excepcionales, como las anguilas, pasan de los ríos al mar para reproducirse. De los peces marinos que remontan los ríos para reproducirse, algunos han permanecido allí por elección propia, otros se han quedado aislados debido al surgimiento de barreras terrestres y han evolucionado [ p. 290 ] en razas o especies distintas. Gracias a la captura de los ríos y a la dispersión de los huevos en las patas embarradas de las aves zancudas, los peces de agua dulce han alcanzado su actual amplia distribución sobre los continentes.
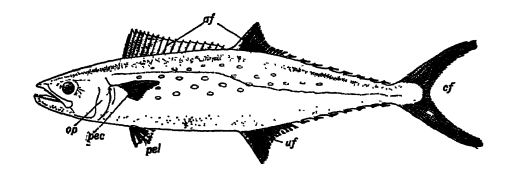
Características distintivas. — Los peces son vertebrados sin cuello. Carecen de orejas y, por lo tanto, carecen de tímpano. Su temperatura corporal se aproxima a la del medio en el que viven, por lo que se les considera de sangre fría. Debido a su hábitat acuático, todos los peces tienen branquias y aletas (véase la fig., pág. 289). Las branquias se encuentran a cada lado de la cabeza y consisten en filamentos delicados y huecos por donde circula la sangre. El agua se absorbe por la boca y se expulsa a través de estos órganos respiratorios, donde la sangre de los tubos extrae el oxígeno libre disuelto en ella. La boca suele estar provista de dientes. Generalmente presentan una vejiga natatoria que puede servir como órgano hidrostático o flotador, aunque en ciertas formas se transforma en un saco celular, que actúa como pulmón y ayuda a los pulmones en la respiración.
La piel es suave y desnuda, o, más comúnmente, protegida por el desarrollo de espinas, mosaicos de dentículos, escamas superpuestas o placas óseas conocidas como escudos. El principal órgano de locomoción es la poderosa cola. Esta es asistida por las aletas pares, de las cuales el conjunto delantero, conocido como las extremidades pectorales, se encuentra justo detrás de las aletas, y el par trasero, o extremidades pélvicas, se coloca más atrás (ver Fig., p. 289). Las aletas pares de los peces corresponden a las extremidades anteriores y posteriores de los vertebrados superiores. Sin embargo, su esqueleto no puede compararse fácilmente con la estructura de las extremidades de otros vertebrados. Cómo estas aletas se convirtieron en extremidades se discute más adelante en este capítulo. Los peces también suelen poseer un número variable de aletas medianas impares que se utilizan principalmente como órganos de equilibrio; Las del dorso se llaman aletas dorsales, las del lado inferior, entre el ano y la cola, se conocen como aletas anales, y la cola representa la aleta caudal (véase la fig., pág. 289). Todas las aletas están sostenidas por barras o radios esqueléticos, llamados radios de aleta.
El esqueleto puede ser cartilaginoso, aunque suele ser más o menos óseo. El esqueleto principal está formado por la columna vertebral. Los huesos de la cabeza son de dos tipos: dérmicos o cutáneos, como las escamas, y los huesos de la caja encefálica.
Tipos de colas de peces. — La cola presenta tres tipos generales. (1) La columna vertebral continúa con articulaciones que disminuyen constantemente casi hasta el extremo del animal, y allí está rodeada por una aleta caudal vertical, triangular y unilobulada. Este es el tipo más primitivo, muy común entre los peces del Devónico, y se observa en las etapas [ p. 292 ] de cría de [ p. 291 ] la mayoría de los peces vivos; se conoce como cola difícil o simétrica (véase Lám., pág. 295, Fig. 4). (2) En otros, la columna vertebral se curva hacia arriba y termina con articulaciones que disminuyen en el lóbulo superior y más grande. Por lo tanto, este tipo de cola rara vez es simétrica, y se denomina heterocerca debido a la desigualdad de los lóbulos (véase Lám., pág. 291, Figs. 2, 4; Lám., pág. 295, Figs. 1, 2). Es común en los tiburones actuales y en la mayoría de los peces del Paleozoico. (3) Los peces óseos modernos también tienen una cola simétrica, pero si bien la columna vertebral se curva hacia arriba, termina de forma bastante abrupta, y desde las articulaciones terminales se desarrollan los radios de las aletas formando una cola simétrica. Este tipo de apéndice caudal se denomina homocereal (véase la figura a continuación).
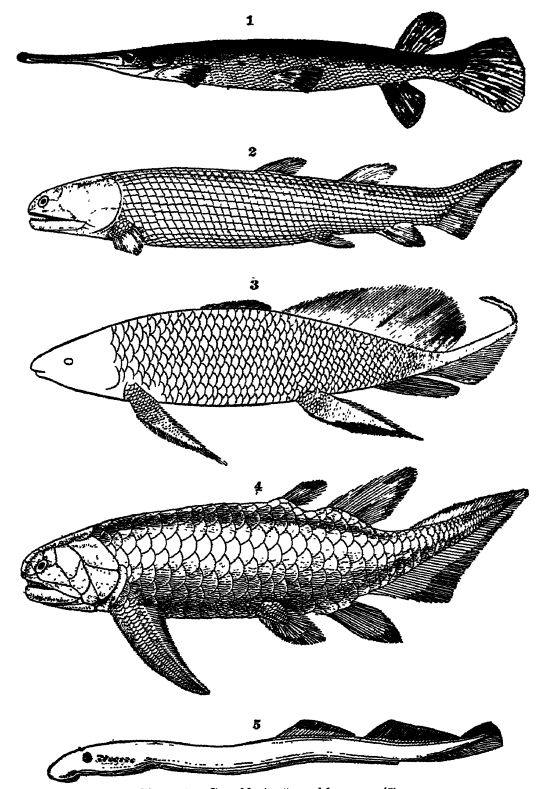
Fig. 1, living American gar-pike (Lepidosietis osseiLs), x 2, Devonian ganoid from Scotland (Osteolepis macrolepidotu8)i x J; 3, Upper Devonian ganoid from Scaumenac Bay, Canada (Scaumenacia curia), x 4, Devonian ganoid with Q’cloid scales, from Scotiand (ffolopiychius flemingi), X §; 5, Innng marine lamprey (Pefromyzon marinus), x i, note the seven openings back of the eye, exits from the gills; also edge of circular mouth, and absence of paired fins.
Figs. 2 and 4 after Woodward, British Museum Guide Book; Fig. 3 after Hussakof.
Alimento de los Peces. — Entre los peces vivos, algunos se alimentan exclusivamente de plantas, otros de plantas y animales por igual, otros exclusivamente de animales, y otros del lodo donde crecen plantas y animales diminutos. Sin embargo, la mayoría de los peces se alimentan de otros peces, sin importar la especie, devorando a sus crías con la misma facilidad que las de cualquier otra especie. Los dientes con puntas romas suelen servir para triturar conchas; los dientes con bordes dentados a menudo indican que se alimentan de plantas; mientras que los incisivos fuertes pueden indicar la preferencia por las aletas y los crustáceos como alimento. En general, cuanto más afilados sean los dientes y más grande la boca, más claramente se distingue una especie de pez. Los peces de boca pequeña tienden a ser herbívoros o de lodo.
Crecimiento. — Los peces crecen mientras viven y su tamaño depende de su ascendencia más la cantidad de alimento que puedan conseguir: la duración de su vida es variable; algunos viven muchos años, otros viven poco y algunos parecen ser anuales.
Ascendencia de los Peces. — A pesar de que la mayoría de los peces tienen esqueletos óseos y de que se conocen animales similares a los peces desde el Champlainiano Medio, sus ancestros fósiles más primitivos aún no han sido descubiertos por los paleontólogos. Los zoólogos tampoco pueden determinar su ascendencia entre la vasta horda de formas vivas y sus embriones. Lo único que los paleontólogos pueden afirmar es que los vertebrados fosales más antiguos son, sin duda, peces, y aparentemente de los tipos elasmobranquios y ostracodermos. En estas circunstancias, [ p. 293 ] dejaremos este asunto para futuros descubrimientos y pasaremos directamente a la descripción de los tipos de peces.
Clasificación de los peces. — Para la presente clasificación, los peces pueden dividirse de la siguiente manera:
- Clase Piscis o verdaderos peces.
- Subclase Elasmobranchii o peces cartílago
- Orden Acanthodei o tiburones espinosos
- Orden Selachii o verdaderos tiburones y rayas
- Subclase Ostracodermi o tiburón extinto aberrante
- Subclase Ganoidei o peces sellados con esmalte
- Subclase Teleósteos o peces óseos
- Subclase Dipnoi o verdaderos peces pulmonados
- Subclase Arthrodira o peces acorazados
- Subclase Elasmobranchii o peces cartílago
¶ Subclase Elasmobranquios o Peces Cartílago
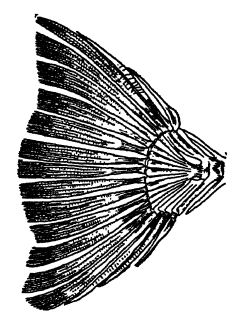
El término elasmobranquios significa «con branquias en forma de placa», y el grupo incluye a los antiguos y modernos tiburones, peces sierra, gatos marinos (quimeras), rayas y mantas. Con pocas excepciones, estas formas vivas habitan aguas marinas (véase lámina, pág. 295, fig. 3). Son los peces más primitivos y, muy temprano en el Paleozoico, dieron origen a los tipos superiores.
En la mayoría de los elasmobranquios, la boca se encuentra en la parte inferior de la cabeza, pero en algunos se encuentra en el extremo delantero. El esqueleto interno es de cartílago o cartilaginoso, con mayor o menor calcificación. La cola suele ser heterocerca. La piel a menudo parece estar desnuda, pero siempre hay dentículos dérmicos (pequeños trozos) con la punta de esmalte duro y brillante, con un núcleo de dentina (fosfato y carbonato de calcio con flúor) y una base ósea (fosfato de calcio) hundida en la parte más profunda de la piel. Este tipo de escama se llama placoide y tiene la misma estructura que los dientes, que originalmente también se desarrollaron a partir de la piel. Cuando los dentículos son pequeños y están muy juntos, constituyen el llamado shagreen (que significa un mosaico rugoso).
Las cinco hendiduras branquiales a ambos lados de la parte posterior de la cabeza, en los peces cartilaginosos actuales, están abiertas o cubiertas por una membrana cutánea, nunca por un opérculo óseo como en la mayoría de las demás especies. Las aletas medias suelen estar provistas de espinas largas, afiladas, homicidas u óseas, sobre todo en las dorsales. Estas espinas y los dientes de tiburón son los que suelen conservarse como fósiles; sin embargo, por desgracia, aportan poca información sobre las características generales de los animales.
El cerebro está encerrado en una sólida caja cartilaginosa o cráneo sin suturas ni articulaciones, y en esta característica los elasmobranquios [ p. 294 ] se diferencian de casi todos los demás peces (con la rara excepción de los ganoideos), en los que los huesos del cráneo se distinguen fácilmente. Carecen de vejiga natatoria.
Los tiburones acantodios o espinosos son pequeños y los más primitivos de los verdaderos peces j. Aparecen a finales del Silúrico, son particularmente comunes en la arenisca roja antigua de Escocia y rara vez superan los 30 cm de longitud (véase la lámina, pág. 295, fig. 1). Se diferencian de los verdaderos tiburones y de la mayoría de los demás tiburones paleozoicos por su piel laminar, en la que se incrustan pequeñas placas óseas aplanadas y ajustadas, compuestas de dentina y esmalte, y por el hecho adicional de que las robustas espinas no se limitan a las aletas medias, sino que también se desarrollan en conexión con las aletas pares. Los dientes son pequeños y bastante puntiagudos, como los de los tiburones modernos. Los tiburones acantodios son, directa o indirectamente, los progenitores de los tiburones posteriores y de los ganoides superiores.
Los tiburones comedores de concha estaban especialmente bien representados en el Paleozoico tardío, por dos familias conocidas como cestraciontos y cocliodontos. El primer grupo está ahora mejor representado por el tiburón de Port Jackson, que vive frente a la costa de Australia del Sur (véase Pl., p. 295, Fig. 3), y se caracteriza por tener dos espinas gruesas en la aleta dorsal y, en las partes más profundas de la boca, muchos dientes romos y similares a pavimento adaptados para triturar mariscos y crustáceos (por lo tanto, también conocido como el «triturador de ostras»). En los tiburones cocliodontos, los dientes individuales están fusionados en dos grandes placas cuya superficie superior tiene crestas curvadas en espiral que sugieren las espirales de las conchas de los caracoles, la característica que dio origen al término familiar (véase Fig. A, p. 342). Son estas partes duras las que se ven tan a menudo en el Paleozoico tardío y mucho menos comúnmente en el Mesozoico; Los cocliodontos, sin embargo, desaparecieron con el Carbonífero.
Los tiburones modernos son los verdaderos selaquíos (término griego que significa tiburones). Este orden incluye a los grandes y activos tiburones fusiformes, los peces sierra y las rayas y mantas aplanadas y perezosas que viven en el fondo marino, tanto de la época actual como de las eras Cenozoica y Mesozoica. Este tipo de tiburón, con dientes afilados y cortantes, ha sido abundante. En el Mioceno de Carolina del Sur se encontraron dientes de este tipo, el más grande de los cuales mide casi 15 cm (Carcharodon), lo que indica que los tiburones pueden alcanzar una longitud de 18 metros. El gran tiburón azul actual alcanza los 12 metros de longitud.
Presencia geológica. — En América, los restos más antiguos de tiburón son pequeñas espinas de aleta, aparentemente de especies acantódicas (Lám., pág. 295, Fig. 1), halladas a finales del Silúrico (Cayugan y no Clinton, como se afirmó originalmente) de Pensilvania. Con el Devónico medio, las espinas, dientes y dentículos de piel de tiburones pequeños y grandes se vuelven comunes. En el Misisipiense, las formas que se alimentaban de concha estaban muy diversificadas (véase pág. 341), pero perdieron su predominio en el Pensilvánico y prácticamente desaparecieron con el Paleozoico. El tipo de diente cortante, tan común en los tiburones actuales, es raro en el Paleozoico, se observa con más frecuencia en el Mesozoico temprano, pero no es común hasta el Cretácico.
¶ Subclase Ostracodermi o tiburones aberrantes
Los ostracodermos poseen una armadura pesada y posiblemente sean un grupo de tiburones cartilaginosos altamente aberrantes. Esta subclase incluye los peces más antiguos conocidos, comenzando en el Champlamiano Medio y desapareciendo a finales del Devónico. Representan una línea de evolución de los peces [ p. 296 ] en la dirección equivocada, pues no han dejado descendientes. De hecho, son tan diferentes de otros peces auténticos que aún es imposible indicar su parentesco con otros vertebrados. Los ostracodermos eran pequeños, ninguno de más de 18 cm, probablemente se alimentaban en el fondo y tenían hábitos perezosos, aparentemente viviendo principalmente en las zonas lagunares de los mares y en los ríos. La cabeza y la parte anterior del cuerpo eran grandes, anchas y deprimidas, y estaban protegidas por pequeñas placas o escamas placoideas, que a veces se fusionaban con placas óseas más o menos gruesas, mientras que la región posterior estaba desnuda o más o menos cubierta por placas o escamas similares. La cola era heterocerca y no tenía esqueleto interno (véanse las figuras arriba y pág. 297).

Fig. 1, Lower Devonian acanthodian shark, from Scotland (Climatius X i: 2. Upper Devonian shark, from Ohio (Cladoselache fyleri), x J; 3, livin Port Jackson cestraciont shark, from Austraha (Cestracion philippi) x 1/10; livin lung-fish of Australia (Neocerotodus fosters) X A; 5, Devonian lung-fish,

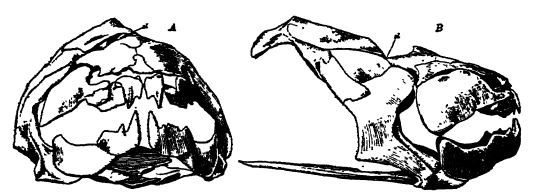
En estructura y forma, existía una considerable variedad entre los peces de piel ósea. Las especies más antiguas, de constitución muy diversa, carecían de un par de palas laterales o extremidades natatorias (Pteraspis, Cephalaspis, Drepanaspis), y los ojos estaban muy separados o muy juntos (véase la fig., pág. 297). Restringidas al Devónico Superior se encuentran las formas con palas anteriores acorazadas, una armadura definida sobre la región anterior y ojos muy juntos (Bothriolepis, Pterichthys, considerados en su día escarabajos gigantes, véase la fig., arriba). [ p. 297 ] Sin embargo, es posible que estos últimos géneros —los «peces alados» de Hugh Miller, quien comentó que su primera impresión fue haber encontrado en ellos «el nexo de unión entre las tortugas y los peces»— no pertenezcan a esta subclase. Los extracodermos están ampliamente distribuidos por Europa y Norteamérica.
¶ Subclase Ganoidei o Peces de escamas esmaltadas
El término Ganoidei, que significa apariencia brillante, se refiere a la superficie lustrosa de la mayoría de estos peces, como, por ejemplo, en los lucios actuales (véase la Lám., pág. 291, Fig. 1). Este brillo se debe a las escamas rómbicas más o menos gruesas que presentan los lucios o a las escamas cicloides de las formas devónicas, que están cubiertas de un esmalte brillante del mismo tipo que la capa externa de los dientes. Las escamas ganoideas se diferencian de las placoides de los elasmobranquios en que solo tienen dos capas: un esmalte externo y una capa ósea más gruesa debajo. Sin embargo, en algunos ganoideos existen escamas cicloides elásticas auténticas, tan comunes en los peces comestibles, es decir, delgadas placas circulares de fosfato de calcio, pero sin esmalte (véase la Lám., pág. [ p. 298 ] 291, Fig. 4). Los ganoideos son esencialmente peces de agua dulce, aunque algunos son marinos y solo remontan los ríos para desovar. En el pasado geológico, los miembros marinos parecen haber sido considerablemente más comunes que en la actualidad. Los ejemplos vivos más conocidos son los esturiones y los lucios, pero el origen de esta subclase se remonta al menos al Devónico Inferior, pues constituyen la mitad de la ictiofauna conocida en la arenisca roja antigua (véase lámina, pág. 291, figs. 2-4) y continuaron siendo abundantes hasta principios del Cretácico. En su organización, forman un vínculo entre los tiburones acantodios y los peces óseos.

El cráneo está bien protegido por huesos dérmicos o completamente osificado, y si bien el esqueleto interno es más o menos óseo, existen algunas formas en las que es completamente cartilaginoso. En esta subclase aparece por primera vez la boca perfeccionada de pez, en la que la mandíbula inferior se apoya en los dientes de la mandíbula superior perfeccionada, como en los vertebrados superiores. Siempre existe una vejiga natatoria conectada con el esófago; en los lucios, esta ayuda a la respiración, pero nunca es celular como en los peces pulmonados. El intestino presenta una válvula espiral, y la cola es heterocercal o dificercal.
Ganoideos con aletas lobuladas. — Según las características de sus aletas, los ganoides se dividen en dos órdenes: (1) los ganoides con aletas lobuladas o con flecos (Crossopterygii), y (2) los ganoides con aletas pilar (Stylopterygii). En este último tipo, las extremidades no presentan los lóbulos gruesos, musculosos y cubiertos de escamas del otro orden, sino que se parecen más a los de los peces comestibles. Del primer orden solo existen actualmente dos géneros, ambos habitando las aguas dulces de África. Aunque su vejiga natatoria también funciona como pulmón, estos peces no pueden vivir fuera del agua más de tres horas. Fueron los peces comunes del Devónico Inferior, siendo las formas más conocidas los géneros Holoptychius, Eusthenopteron y Osieolepis (Lám., p. 291, Figs. 2-4). En estos ganoides, las capas óseas de los dientes cónicos suelen presentar pliegues profundos y complejos, o ser laberínticas, lo que resulta particularmente interesante porque el mismo tipo de dientes se encuentra en todos los anfibios del Paleozoico. Esta similitud en la estructura dental es otra pista que muestra el probable origen de los anfibios (orden Stegocephalia) en estos ganoides de agua dulce del Devónico con aletas finas.
¶ Subclase Dipnoi o verdaderos peces pulmonados
Los dipnoi (del griego aning, que significa «doble respiración») son los «peces pulmonados», llamados así porque los tres géneros actuales poseen una vejiga natatoria que se abre en la boca, simple o doble, con numerosos espacios celulares. Esta es una prolongación de la región anterior del tracto digestivo y funciona como pulmón, complementando, y en épocas de sequía, suplantando, las funciones de las branquias como órganos respiratorios. Desde un punto de vista evolutivo, los peces pulmonados son muy importantes, ya que la vejiga natatoria es comparable [ p. 299 ] al pulmón de los vertebrados superiores, ya que devuelve la sangre aireada directamente al corazón, mientras que en la mayoría de los demás peces la sangre se transporta desde la vejiga natatoria a través de la circulación general antes de llegar al corazón. Los sacos nasales se encuentran en el exterior del hocico y se abren en la boca, una condición que también se encuentra en anfibios y vertebrados superiores, pero muy raramente en peces. Es en los peces pulmonados, pues, donde vemos las posibilidades para el desarrollo de los vertebrados superiores, aunque esto no significa necesariamente que fueran los progenitores de estos últimos. En muchos sentidos, presentan una estructura intermedia entre los peces inferiores y las salamandras anfibias, pero no se cree que sean el nexo de unión directo entre los peces y los anfibios. Es probable que los ganoides con aletas de flecos (crosopterigios) fueran los progenitores de los anfibios.
De los peces pulmonados actuales, sabemos que ciertas formas (Lepidosiren) habitan arroyos fangosos y pantanos, donde se alimentan de plantas. Otras se alimentan de gusanos, insectos, mariscos, crustáceos y ranas. Son animales perezosos que ocasionalmente suben a la superficie para respirar; en épocas de sequía, los Lepidosiren se encierran en un capullo de limo y arcilla, y respiran a través de una abertura hecha en el lodo. Durante la temporada de lluvias, los peces pulmonados vivos comen vorazmente, almacenando grandes cantidades de grasa entre los músculos, la cual les permite sobrevivir durante la temporada seca, de modo que pueden vivir sin alimento, a veces hasta ocho meses, hasta que el agua regresa.
En las formas actuales, también presentan dos o cuatro pares de branquias cubiertas por un opérculo óseo móvil. Generalmente, tienen una apariencia anguiliforme, con las aletas medias dorsal y ventral generalmente continuas con la aleta caudal. En las formas del Devónico, la cola suele ser heterocercal (Lám., pág. 291, Fig. 2), pero en las formas recientes es difícercal (Lám., pág. 295, Fig. 4). Las aletas pares son largas, ampliamente lobuladas, musculosas, escamosas y con flecos en las especies del Devónico y en los Ceratodus actuales, pero en otras formas modernas son muy delgadas, puntiagudas y sin flecos. El cuerpo está cubierto de escamas cicloides superpuestas.
Los dipnos son peces cartilaginosos que presentan cierta osificación, pero carecen de vértebras diferenciadas. El cráneo está compuesto de cartílago recubierto por un número variable de huesos membranosos, que son más gruesos en los géneros paleozoicos. Estos huesos superficiales se desarrollan en la membrana que recubre el cartílago y no se forman en el propio cartílago. Los dientes son escasos, generalmente tres pares, de los cuales dos juegos son grandes placas de trituración.
[ p. 300 ]
Los dipnos están estrechamente relacionados con los ganoides de aletas con flecos. Estos dos tipos de peces son los descendientes más importantes de los tiburones primitivos y evolucionaron hacia los vertebrados superiores. Los dipnos más antiguos, con unas quince especies, se encuentran en el Devónico, tanto en Europa como en América.
¶ Subclase Arthrodira o Peces Acorazados
Los peces más llamativos de los mares Devónicos fueron los Arthrodira, con su gran armadura; casi el 40 % de todos los peces del Devónico pertenecen a esta subclase. El término Arthrodira significa «cuello articulado» y se le atribuye porque la armadura de la cabeza se desliza ligeramente sobre la del cuerpo, lo que permite que la cabeza se mueva hacia arriba y hacia abajo, una característica poco común entre los peces y que se desarrolla mejor en esta subclase. Los Arthrodira tienen una armadura más o menos gruesa y, en este sentido, nos recuerdan a los peces pulmonados y a los primeros anfibios. Sin embargo, su relación sistemática aún se desconoce, pero aparentemente son los más cercanos a los peces pulmonados y a los ganoides de aletas con flecos. Debido a esta supuesta relación, A. Smith Woodward los denomina «peces pulmonados de cabeza grande». Por supuesto, no se sabe nada sobre los pulmones ni los hábitos respiratorios de los Arthrodira.
Los artrodiros más antiguos se encuentran en los depósitos de agua dulce del Devónico temprano de Alemania, mientras que los del Devónico medio y superior de Norteamérica eran marinos. Fueron los peces más grandes y feroces de este período, pero se extinguieron a principios del Misisipiano, época en la que los tiburones conchaleños se alzaron con el dominio de los mares.
Los artrodiros carecían de dientes verdaderos; las estructuras que funcionaban como tales eran grandes tijeras o placas trituradoras. La cola era heterocerca y parece haber tenido un par de dedos [ p. 301 ] pélvicos. El esqueleto interno era escaso, ya que el eje vertebral no era óseo, aunque los arcos neural y ventral estaban algo osificados.
Los Arthrodira dominaron con facilidad la vida acuática de los mares devónicos; una especie americana, Dinickthys, el «pez terrible», alcanzaba una longitud de más de 6 metros (Fig., pág. 300). Se conocen más de cuarenta especies en Norteamérica, y solo una localidad de Alemania (Wildungen) ha presentado más de cincuenta especies.
¶ Subclase Teleósteos o Peces óseos
Los teleósteos (que significa hueso verdadero) son los peces modernos y representan la organización más alta entre estos animales, pues su evolución ha tendido a completar la adaptación al agua (véase la fig., pág. 289). Constituyen aproximadamente el 99 % de todos los peces vivos. La mayoría de los peces comerciales y deportivos pertenecen a esta subclase y abundan en aguas dulces, mares y océanos. Su origen se encuentra en los Ganoidei, aparentemente a finales del Paleozoico, pero no fueron comunes hasta el Cretácico Inferior, alcanzando su máximo desarrollo en el Cenozoico y en la actualidad.
Los peces óseos casi siempre están cubiertos de escamas delgadas, elásticas y cicloides, nunca de escamas placoideas y, muy raramente, de escamas ganoides. En algunos, la piel es desnuda y viscosa. La aleta caudal suele ser homocerca y las extremidades pares nunca son lobuladas. Las branquias siempre están cubiertas por un opérculo móvil. La vejiga natatoria o de aire alargada, cuando está presente, está llena de gas y se encuentra dorsalmente a la cavidad corporal; no es un pulmón, sino un órgano hidrostático. El esqueleto interno está completamente osificado, las vértebras son huecas en ambos extremos (anficoelas), existe un arco neural óseo que encierra la médula espinal y cada vértebra tiene también un par de costillas que encierran las vísceras. Muchas de estas características también se observan, aunque con menos perfección, en los Ganoidei, por lo que algunos naturalistas consideran que ambas subclases representan un solo grupo.
¶ Origen de las aletas y extremidades
Desarrollo de las aletas. — En los animales marinos superiores, la locomoción intencional comienza con la estimulación lateral de un cuerpo flexible, al que se añadieron aletas para facilitar la progresión a través del agua. Las aletas son de dos tipos: los pliegues impares y las aletas pares. Los pliegues impares, originalmente flexibles, surgieron de la piel y luego se volvieron rígidos, dando al pez una superficie más resistente. Estos pueden ser más o menos continuos desde la cabeza a lo largo de la línea media de la espalda (aleta dorsal), alrededor de la cola (aleta caudal) y hacia adelante [ p. 302 ] a lo largo de la parte inferior hasta la abertura nasal (aleta anal). Es habitual en la mayoría de los peces ver estas aletas impares discontinuas, es decir, localizadas.
Las aletas pares, al igual que las impares, probablemente se originaron, según Lull, en pliegues cutáneos que crecían a ambos lados del cuerpo, y que en su origen servían como quillas que proporcionaban un mejor equilibrio al estrecho cuerpo del pez o frenaban su movimiento hacia adelante. Se aprecian claramente en los tiburones acantodios del Silúrico y el Devónico (Lám., p. 295, Fig. 1), y en este caso se mantenían extendidas por robustas espinas óseas, al igual que las dorsales impares, sostenidas verticalmente en los tiburones modernos. En otras formas, estas espinas óseas laterales fueron sustituidas por varillas internas de refuerzo. Osborn afirma que, en el curso de la evolución, estas varillas se concentran para formar el eje central de una aleta completamente articulada, y con un mayor desarrollo se transforman en los cartílagos y huesos de las cinturas pélvicas (véase la Fig., arriba).
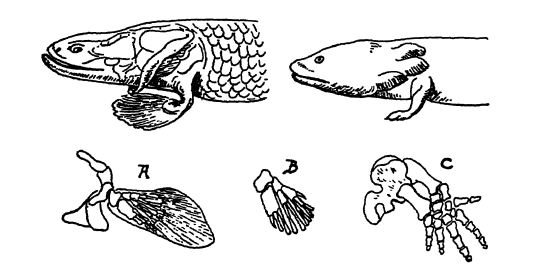
En los peces, las aletas pares con sus cinturas son los rudimentos a partir de los cuales se desarrollaron las piernas y los pies mediante el desplazamiento forzado de los ganoides con aletas de fleco en su búsqueda de pozos de agua en regiones desérticas. Se cree que en ellos se desarrolló una estructura profética de piernas y pies, aunque la extremidad transicional axial, ya sea en forma fósil o en embriones de vertebrados con extremidades vivas, es completamente desconocida.
La huella más antigua conocida, Thinopus (Fig., p. 331), del Devónico Superior, podría dar una pista de cómo fue esta evolución. Parece que solo hay dos dedos completamente formados, probablemente el primero y el segundo, separados [ p. 303 ] por un dedo que se adentraba profundamente en la planta del pie. El fósil indica además la presencia de un tercer dedo nidimentario y posiblemente incluso un cuarto. Si la estructura de esta huella es fiable, entonces el pie terrestre comenzó con dos dedos en lugar de cinco, como se suele creer.
¶ Origen de la doble respiración en los peces
Órganos Respiratorios de los Animales Marinos. — Un estudio de animales marinos típicos revela numerosos y diversos métodos para la extracción de oxígeno libre del agua. Sin embargo, sea cual sea el tipo de órgano respiratorio, siempre es del mismo tipo fundamental: uno o más órganos localizados con canales delicados y membranas muy delgadas por donde circula la sangre, poniendo los glóbulos rojos en contacto casi directo con el agua circundante. La hemoglobina de estos glóbulos rojos tiene una marcada afinidad por el oxígeno, que extrae del agua y que las células flotantes transportan a través de la circulación sanguínea a todas las partes del cuerpo del animal. En ningún lugar del mar ningún animal posee un órgano adicional para la extracción de oxígeno directamente del aire, excepto en aquellas poblaciones que se sabe que tuvieron antepasados terrestres.
Evolución de los Pulmones. — En la mayoría de los peces, por encima de los tiburones, existe un órgano conocido como vejiga natatoria o aérea, que estructuralmente es una excrecencia en forma de saco, simple o doble, del tubo digestivo. La función principal de esta estructura degenerada y modificada secundariamente en la mayoría de los peces vivos es la de un órgano hidrostático, para proporcionar flotabilidad a los animales, y está controlada por músculos que permiten a su portador mantenerse a cualquier nivel deseado en el agua. La vejiga natatoria es homóloga de los pulmones de los vertebrados terrestres y es utilizada por los llamados peces pulmonados, y en menor medida por otros peces reliquia (Amia, Lepidostevs, Pdypterus) como órgano respiratorio complementario a las branquias.
La amplia gama de modificaciones de esta estructura demuestra su origen extremadamente antiguo, y su estado incipiente posiblemente se evidencie en un par de protuberancias similares a bolsas en la faringe, o cavidad de la garganta, de los tiburones. El estancamiento del agua y la pérdida de oxígeno libre harían que los peces subieran a la superficie para absorber aire, y dichas bolsas, si estuvieran provistas de vasos sanguíneos, servirían de forma muy rudimentaria para ayudar a la aireación de la sangre. Se cree que una mayor importancia a estas estructuras estimularía su desarrollo hasta alcanzar la condición observada en los peces pulmonados modernos.
Origen de los Pulmones en Animales que Viven en Cuerpos de Agua Temporales. — Los animales que se originan y viven en cuerpos [ p. 304 ] de agua permanentes, y especialmente en el océano, no necesitan respirar aire, por lo que se sostiene que el estímulo para tal alteración podría haber surgido donde el agua les faltaba periódicamente. No se conoce ningún pez que respire aire permanentemente en la región entre mareas bajas y altas, aunque varias especies viven aquí durante un tiempo en busca de alimento. Por otro lado, en ninguno de los deltas marinos actuales se encuentran las condiciones climáticas necesarias para obligar a los peces a desarrollarse como peces pulmonados, ni se encuentran en las zonas costeras ni en los cauces de los ríos permanentes. En estos lugares no existe una zona de transición que obligue a los peces acuáticos a adaptarse a la tierra firme.
Así como hoy en día existen regiones áridas, parece que existieron climas terrestres similares durante gran parte del tiempo geológico. En estos climas, las masas de agua aparecen y desaparecen según la temporada de lluvias y sequías, por lo que los animales recurren a diversos métodos para sobrevivir durante la sequía. Durante la temporada árida, la lucha por la supervivencia es intensa, no solo por la aglomeración anormal de los individuos en espacios cada vez más oscuros y la reducción de la cantidad de alimento disponible, sino aún más por el carácter cada vez más salino y amargo del agua. Se cree que, bajo el estímulo de estos cambios, los peces con respiración branquial se adaptaron inicialmente a excavar en la arena. Así, protegidos en charcas de agua y lodo, la humedad pudo pasar por las branquias durante un tiempo, pero en tales entornos la vida era muy precaria y, en la lucha, la mayoría de los individuos fueron destruidos. Después de innumerables fracasos en sus esfuerzos por tragar aire hacia la faringe, esfuerzos que duraron durante un largo tiempo geológico, los ganoides y los peces pulmonados se desarrollaron y perfeccionaron gradualmente, siendo su primera aparición a principios del Devónico.
Mentalidad Superior entre los Vertebrados Terrestres. — La respiración aérea, una vez establecida entre los peces, ha resultado en que los vertebrados terrestres alcancen la máxima perfección mecánica y mental, una evolución necesaria para la adaptación a una amplia gama de condiciones ambientales en comparación con la relativa uniformidad del mar. La mentalidad más elevada en el mar se ha derivado repetidamente de los continentes, ya que muchas poblaciones de vertebrados terrestres se han adaptado al mar debido a la facilidad con la que pueden allí depredar a los menos alertas e inteligentes. Sin embargo, de estas poblaciones no surge una mentalidad superior. Representan una adaptación en la dirección equivocada, es decir, hacia una vida más fácil, pues la mentalidad más moderna se ha desarrollado solo en la tierra, donde [ p. 305 ] la lucha por la existencia es mayor debido a la constante necesidad de adaptación a un entorno cambiante. La supremacía orgánica se alcanza solo mediante la vigilancia constante.
¶ Lectura colateral
Joseph Barrell, Influencia de los climas silúrico-devónicos en la abundancia de vertebrados aerorespiradores. Boletín de la Sociedad Geológica de América, vol. 17, 1916, págs. 387-436.
T. C. Chamberlin, Sobre el hábitat de los primeros vertebrados, Journal of Gcolo©% Vol. 8, 1900, págs. 40(M12.
Bashford Dean, Peces vivos y fósiles. Nueva York y Londres (Macmillan), 1895.
C. R. Eastman, Peces Devónicos de las Formaciones Neoyóricas. Museo Estatal de Nueva York, Memorias 10, 1907.
R. S. Lull, Evolución orgánica. Nueva York (Macmillan), 1917.
H. F. Osborn, El origen y la evolución de la vida, Nueva York (Scribner), 1917.
| XXII. Corales y animales similares a los corales | Página de título | XXIV. El Devónico y el predominio de los peces |