| XXXVI. Los dragones de la época medieval | Página de título | XXXVIII. El Cretácico Inferior y la primera aparición de las plantas con flores (angiospermas) |
[ p. 528 ]
¶ Amonitas
Características generales. — Los mares mesozoicos se caracterizaron por la abundancia de cefalópodos con concha conocidos como ammónidos (véase lámina, pág. 477, figs. 4-16). Estas hermosas conchas enroscadas, parientes de los nautilidos, presentaban una gran variedad de tamaño, forma, ornamentación y características de sus septos (las placas transversales que dividen la cavidad de la concha en cámaras), y los paleontólogos han descrito más de seis mil formas. Probablemente, el tamaño promedio oscilaría entre 7,5 y 10 cm, aunque pueden alcanzar diámetros de hasta 2,4 metros (Pacjidiscus seppenradensis del Cretácico Superior de Alemania), y si el tubo enroscado de estos grandes se enderezara, la longitud sería de entre 6 y 10,6 metros. Por lo tanto, los ammónidos superaron a los nautilidos paleozoicos tanto en tamaño como en diferenciación específica. Parecen haber sido animales más activos y mejores nadadores y flotadores que los nautilidos, por lo que se arrastraban menos por el fondo marino. Si [ p. 529 ] eran buenos nadadores, también debieron tener un hiponoma funcional (véase Fig., p. 226) como los nautilidos. La creencia de que nadaban bien se deduce de la naturaleza de sus conchas muy delgadas, la amplia distribución de algunas especies y la poca profundidad o estrechez de los conos enrollados (véase Fig., p. 528). Esta afirmación se aplica prácticamente a todos los tubos que se enrollan en un plano como el muelle de un reloj, pero donde las conchas son espirales, más o menos desenrolladas o rectas, son más gruesas y se cree que los animales se arrastraban en lugar de nadar (véase Lám., p. 575, Fig. 16). Ya no se considera que sean degenerados, como hasta ahora, sino que representan nuevas adaptaciones, aunque reversionarias, en la búsqueda de alimento en los fondos marinos. Es posible que algunos incluso hayan estado anclados en el fondo marino (Nipponitas). Antes de continuar, el estudiante debería refrescar su memoria leyendo lo que se ha dicho sobre los nautilidos en la página 225.

Historia del término. — La palabra ammonite se remonta a 1732, cuando los fósiles se consideraban minerales, como lo indica la terminación ite. Las conchas recibieron este nombre debido a su aparente parecido con los cuernos de carnero, representados como uno de los atributos de la deidad egipcia Amón. En la India, estos fósiles son un artículo de comercio común y se utilizan en todo el país en ciertos ritos religiosos.
Comparación con los nautilidos. — En muchos sentidos las conchas de los ammónidos son como las del nautilus perlado, pero difieren radicalmente de ellos en varios aspectos. Son casi siempre más ornamentadas, más estrechas o menos profundas, y a menudo presentan una quilla distintiva a lo largo del centro de los verticilos externos. Además, la boca de las conchas suele presentar extensiones lobuladas a los lados (lapetas), y en general, presentan una quilla media que puede extenderse hasta una punta afilada, el rostro (véase la fig., pág. 528). Por otro lado, el sifúnculo en los nautilidos se encuentra cerca del centro de los septos, pero en los ammónidos siempre se sitúa cerca o en contacto con el cono en su lado externo o ventral (también llamado vientre). En los nautilidos, la boca de la concha nunca está cerrada por un opérculo, pero en los goniátidos y ammónidos probablemente estaba cerrada en la mayoría de las formas por una cubierta cuando el animal estaba en reposo. Esta cubierta, cuando es delgada y está hecha de quitina, se conoce como anáptico, pero cuando es gruesa y está compuesta de carbonato de calcio en dos piezas articuladas se llama aptico. La cámara corporal es de longitud variable, a veces tan larga como una revolución y media de la concha.
Naturaleza de los septos. — Sin embargo, todas estas diferencias, si bien importantes, no son tan valiosas para la clasificación como lo es la naturaleza de [ p. 530 ] de los septos (véase pág. 226). En los nautilidos, estas particiones en la concha son simples y más o menos cóncavas; en los ammónidos, son simples solo en la parte central, y cada septo se vuelve cada vez más estriado u ondulado hacia su unión con la concha exterior (lám., pág. 477, figs. 10 y 14). Cuando la concha exterior está ausente, como suele ocurrir en estos fósiles, la línea de sutura, o borde de los septos, siempre se ve como una línea ondulada con un patrón distintivo, y este patrón también se vuelve más complejo con la edad (véase fig., pág. 366). Es esta compleja línea de sutura, con sus intrincados lóbulos (dirigidos hacia adentro) y sus sillas de montar (curvadas hacia la boca de la concha), lo que hace que estos fósiles sean tan valiosos para descifrar la cronología del Mesozoico, ya que indica una evolución progresiva determinada a partir de la secuencia de los estratos, controlada por el crecimiento u ontogenia del individuo. El valor temporal de las suturas fue observado por primera vez por Yon Buch en 1830.
A menudo se pregunta: ¿Por qué estos animales desarrollaron un tipo de tabique tan complejo? Entre los nautilidos rara vez vemos algo parecido al tabique lobulado, y en este caso, la cámara vital no solo presenta una profundidad variable, sino también una anchura considerable. En otras palabras, la cara ventral del cono es redondeada y ancha, y las conchas suelen ser considerablemente más profundas que las de los amónidos. Por lo tanto, los músculos laterales que sujetan la concha al animal no solo tienen una mayor superficie de apoyo en los nautilidos, sino que, lo que es más importante, el cono es más o menos redondo y, por lo tanto, se apoya de forma más uniforme sobre todas las partes del animal. Mientras que en los amónidos, las conchas son lenticulares y, por lo tanto, se desvían con mayor facilidad hacia un lado al moverse en el agua. Probablemente, la estrechez y la naturaleza comprimida de las conchas llevaron a la lobulación de los tabiques, debido a la necesidad de aumentar la fuerza de sujeción del animal sobre la concha. En cualquier caso, los septos lobulados solo se observan en las conchas lenticulares de los cefalópodos (nautilidos, goniátidos, amónidos).
Evolución. — Los nautilidos de concha profunda de finales del Silúrico dieron origen a goniátidos pequeños y de concha estrecha con septos escasamente lobulados. En la mayoría de los goniátidos del Devónico (Lám., pág. 322, Figs. 1-6), la línea de sutura de los lóbulos y las sillas de montar termina de forma aguda, pero en el Misisipiano la mayoría de las especies no solo presentan más lóbulos y sillas de montar, sino que estos son casi todos redondeados. Así, observamos un cambio gradual en los cefalópodos de concha estrecha, comenzando como verdaderos nautilidos y pasando a diversas estirpes de goniátidos de concha más o menos estrecha que finalmente dieron origen, en el Carbonífero, a numerosas líneas evolutivas entre los ammónidos primitivos, resultando en el establecimiento de estos últimos en las aguas más cálidas del Pérmico del Mediterráneo tetiano (Fig., pág. 431). La mayor variación entre ellos tuvo lugar a finales del Pérmico y principios del Triásico. Al final del Triásico, casi se extinguieron, como se explica [ p. 531 ] en el capítulo sobre el Triásico. En el Jurásico, se produjo otra rápida evolución a partir de un género (Phylloceras), alcanzando el cenit de su desarrollo en ese momento. El declive de los ammónidos comenzó en el Cretácico Inferior, y durante el Cretácico Superior, las especies mostraron poco de su antigua virilidad. Su extinción completa se produjo al final del Cretácico, durante la época crítica de la Revolución Laramide.
¶ Belemnis, Calamares o Peces Tinta
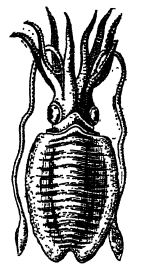
Descripción general. — En el Mesozoico también presenciamos el surgimiento de los belemnítidos, ancestros de las sepias (también conocidas como jibias o calamares, Fig., lado opuesto). La palabra belemnite proviene del nombre griego para dardo, y los fósiles fueron considerados en su momento como los rayos de Thor, el dios del trueno (Fig. A, p. 532). Hugh Miller relata que la gente del campo consideraba estos fósiles «de gran eficacia para curar al ganado hechizado». Los belemnítidos eran cefalópodos muy activos y altamente carnívoros, que se alimentaban de peces, cangrejos y moluscos.
Poseían ojos grandes y completamente desarrollados, carecían de conchas externas y, debido a que solo poseían dos plumas branquiales internas, se les ha dado el nombre de Dibranchiata. Poseían diez brazos, posiblemente ocho cortos y dos largos protráctiles, como los calamares actuales (Fig., lado opuesto). En el Mesozoico, estos brazos solían estar provistos en su interior de ganchos quitinosos curvados para sujetarse y, más raramente, de ventosas. Posteriormente, las ventosas se convirtieron en el tipo dominante. Al principio, los belémnidos tenían un esqueleto interno pesado, pero durante el Mesozoico, de la estirpe de belémnidos surgieron los calamares, animales que perduraron a medida que el tiempo geológico perdía cada vez más de su esqueleto ancestral, aunque aún quedan vestigios de este en todas las formas vivas. En los calamares, el cerebro está altamente especializado y representan la máxima expresión de alerta entre los invertebrados. Por esta razón, se les ha llamado los «piratas de las profundidades».
Saco de tinta. — Todos los dibranquiados poseen un saco de tinta interno que contiene sepia, un fluido marrón negruzco que se mezcla fácilmente con el agua de mar; los animales lo arrojan al frente cuando están en peligro [ p. 532 ] y luego escapan hacia atrás, alejándose de la pantalla defensiva de agua coloreada. La sepia de los artistas es un pigmento hecho de esta sustancia. Humboldt, en su Cosmos (1844), afirma que esta tinta está tan bien conservada en los especímenes jurásicos que aún conserva el color con el que se puede dibujar la imagen de los animales. Al igual que los demás cefalópodos, los dibranquiados poseen un sifón a través del cual se expulsa el agua hacia adelante, impulsándolos hacia atrás. También pueden nadar con la ayuda de sus aletas laterales.
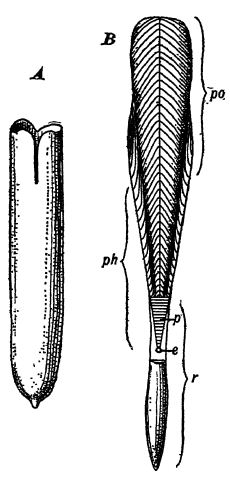
Naturaleza del esqueleto. — Los belémnidos se originaron a principios del Triásico (Lám., pág. 477, Fig. 3) a partir de los ortocéridos del Paleozoico (Lám., pág. 236, Fig. 19), en el sentido de que los animales, por así decirlo, se abrieron paso cada vez más fuera de las conchas externas y las envolvieron con sus mantos de tal manera que se convirtieron en esqueletos internos, un pedigrí interno que indica su linaje (véase la Fig., opuesta). Entre los fósiles, la parte que se conserva generalmente se llama guarda, un cuerpo calcáreo sólido, con forma más o menos de cigarro, puntiagudo en un extremo y una profunda fosa circular en el otro. Estas guardas a veces miden hasta 2 pies de largo y 4 pulgadas de grosor. En la fosa de las guardas y extendiéndose más allá de ellas se encuentra el ‘phragmacone’, un vestigio de su ancestro ortocérido, con cámaras y sifunculado. Esta se extiende hacia adelante en la cara dorsal hasta formar un trozo más o menos largo, ya sea de cal o de quitina, conocido como proöstracum (véase la Fig. B, arriba). Los belémlidos son muy característicos del Mesozoico, constituyen excelentes manadas estratigráficas y se conocen unas cuatrocientas especies, en unos sesenta géneros.
[ p. 533 ]
Los auténticos escuadrones del Mesozoico conservaban únicamente el proóstraco y un remanente completamente modificado del fragmacón, que juntos formaban una secreción espesa y ancha, pero reticulada y, por lo tanto, ligera, de carbonato de calcio, conocida como jibión. Sobre el proóstraco y proyectándose hacia la cabeza se encontraba la pluma, llamada así por su aspecto similar a una pluma de ave, hecha de quitina. En algunos calamares o jibiones actuales solo se conserva la pluma. En el Jurásico Superior de Solenhofen se encontraron jibiones de hasta 60 cm de largo, lo que indica animales con una longitud corporal probable de 1,8 a 2,4 metros; en los mares actuales habitan los gigantes más grandes, cuyos cuerpos miden 5,5 metros de largo, con dos brazos protráctiles que alcanzan una longitud de 9 metros cuando están completamente extendidos.
¶ Lectura colateral
P. Bartsch, Piratas de las profundidades — Historias del calamar y el pulpo. Informe anual del Instituto Smithsoniano, 1916, 1917, págs. 347-375.
C. Diener, Lebensweise und Verbreitxmg der Ammoniten. Neues Jahrbuch fiir Mineralogie, etc., 1912, Vol 2, págs. 67-89.
C. O. Dunbar, Fases de la adaptación de los cefalópodos. En «Adaptación orgánica», próxima publicación de Yale University Press.
R. Ruedemann, Observaciones sobre el modo de vida de los cefalópodos primitivos. Boletín de la Sociedad Geológica de América, Vol. 32, 1921, págs. 315-320.
E. Boese, Los ammonoides pérmicos-carboníferos de las montañas Glass, oeste de Texas, y su importancia estratigráfica - Universidad de Texas, Boletín 1762, 1917.
S. S. Buckman, Tipo Ammonites, Vols. 1-4 (otros en preparación). Londres (Wesley), 1909-1923.
A. Hyatt y J. P. Sinna, Los géneros de cefalópodos del Triásico de Norteamérica. Servicio Geológico de Estados Unidos, Documento Profesional 40, 1905.
J. B. Reeside, Jr., Algunos amonites jurásicos americanos de los géneros Quenstedticeras, Cardioceras y Amoeboceras, familia Cardioceratidm. Ibid., Professional Paper 118, 1919.
J. P. Smith, Las faunas de invertebrados marinos del Triásico Medio de Norteamérica. Ibíd., Revista Profesional 83, 1914.
| XXXVI. Los dragones de la época medieval | Página de título | XXXVIII. El Cretácico Inferior y la primera aparición de las plantas con flores (angiospermas) |