| XXXIX. Cretácico Superior y el Nacimiento de las Montañas Rocosas | Página de título | XLI. El amanecer de lo reciente en el Cenozoico |
[ p. 582 ]
En todas las tierras actuales y en la superficie de los mares existen más de 15.000 especies de aves, ninguna de las cuales posee dientes. Incluso en el embrión aún no eclosionado, los dientes son discernibles en pocas formas, y solo como meros rudimentos. En el Cenozoico se han descrito unas 650 especies de aves agrupadas en casi 400 géneros, y aquí, como en el mundo actual, todas las conocidas carecen de dientes. Sin embargo, las aves fósiles son extremadamente escasas hasta el Mioceno y depósitos posteriores. Los cráneos de aves son muy raros a finales del Pleistoceno, pero en el Eoceno americano temprano (Río Verde) se conoce un excelente esqueleto cuyo cráneo también carece de dientes. Todo esto sirve para enfatizar la característica principal que distingue a las aves medievales: la posesión de dientes.
Se conocen huevos fósiles de once tipos diferentes: uno del Cretácico y los demás del Cenozoico tardío. Se conservan plumas en nueve especies, y probablemente las impresiones más finas se encuentran en los depósitos de Solenhofen del Jurásico Superior. En el Mesozoico, se conocen esqueletos de buena calidad de aproximadamente media docena de especies, todas de características reptilianas y provistas de dientes.
Origen de las Aves. — Actualmente se acepta ampliamente que las aves, en su constitución general, y especialmente en su cerebro, esqueleto, órganos reproductivos y modo de desarrollo, son «reptiles glorificados». La antigua idea de Huxley de que las aves surgieron de los dinosaurios ha dado paso a la teoría de que, a finales del Pérmico o principios del Triásico, un pequeño reptil parecido a un lagarto, de hábitos parcialmente bípedos y que vivía en el suelo, se adaptó a vivir en arbustos y árboles, probablemente por motivos de seguridad. Se cree que el hábitat arbóreo impulsó a estos reptiles a aprender a volar. Se cree que estos reptiles ancestrales remotos y primitivos (Diapsida) vivieron probablemente en el Pensilvánico, y posteriormente dieron origen no solo a las aves, sino también a los dinosaurios, los pterodáctilos y otros grupos de reptiles. Los progenitores tenían cerebros pequeños, hábitos comparativamente lentos y una temperatura corporal muy variable. [ p. 583 ] Durante los difíciles climas áridos del Pérmico y el Triásico, se desarrollaron reptiles carnívoros de sangre caliente más resistentes y activos. Algunos de ellos, al correr, probablemente se alzaban sobre sus patas traseras, al igual que ciertos lagartos actuales. Estos lagartos pro-aviares, corredores y bípedos más activos, probablemente tenían todo el cuerpo cubierto de escamas superpuestas y, saltando de rama en rama o de árbol en árbol, aprendieron no solo a lanzarse en paracaídas, sino también a batir sus extremidades delanteras en la aviación. En estos esfuerzos, las escamas se transformaron en frondas largas y complejas y finalmente en plumas que mantenían el calor corporal; las extremidades delanteras se convirtieron en alas; el esqueleto se volvió más ligero (hueco y neumático); y, además, el cuerpo se llenó de un extenso sistema de celdas de aire para ayudar a mantener al ave a flote durante el vuelo. Las aves se distinguen además por tener plumas, una característica dérmica que es tan característica de ellas como el pelo lo es de los mamíferos.

Aves Jurásicas. — Las aves aparecen como fósiles por primera vez en el Jurásico Superior y representan uno de los avances más notables que la vida de este período puede mostrar. Hasta el momento, solo se ha encontrado una especie de ave jurásica, procedente de la división superior, cerca de Solenhofen, Alemania. Esta ave, del tamaño aproximado de una paloma grande, se llama Archaeopteryx (del griego, zorro, ala antigua). También se le conoce como el ave de cola de lagarto. Presenta numerosos puntos de semejanza con los reptiles y muchas características que solo se repiten en los embriones de las aves modernas (véanse las figuras, págs. 583 y 584).
Las peculiaridades que llaman la atención a primera vista se encuentran en la cabeza y la cola; no tenía pico, pero las mandíbulas presentaban una hilera de pequeños dientes, mientras que la cola era muy larga y tenía forma de hoja de palmera datilera, compuesta de vértebras separadas (unas 20) y con un par de plumas de ave unidas a cada articulación. Esta construcción muestra de inmediato que en el Archosopteryx la cola no se parecía en nada a la de las aves modernas, es decir, tenía forma de abanico, con las vértebras anquilosadas y las grandes plumas plegadas unas sobre otras. El ala se construyó siguiendo el mismo plan que la de un ave moderna, pero era decididamente más primitiva. Los cuatro dedos eran libres (en las aves recientes, dos de los tres dedos están fusionados); tenían el mismo número de articulaciones que en los lagartos y todos estaban provistos de garras. El plumaje era completamente aviar, pero se caracterizaba por la presencia de plumas de ave en las patas y, al parecer, también por la ausencia de plumas de contorno en la cabeza, el cuello y gran parte del cuerpo, dejando estas partes desnudas. Esta extraordinaria criatura era, por lo tanto, un ave real, pero conservaba muchos rasgos de su ascendencia reptiliana.
[ p. 584 ]
En el noreste de Sudamérica habita uno de los vínculos más notables con el pasado: el ave conocida como hoatzin (Opisthocomus). Es la única ave viva conocida con cinco dedos en las alas. Beebe afirma que el ave adulta tiene voz, movimiento, dedos y hábitos reptilianos. Su voz, ronca y croante, es similar a la de una rana. De cría, «se arrastraban a cuatro patas, trepaban con los dedos de las manos y los pies, se zambullían de cabeza y nadaban con la misma destreza que el antiguo Hesperornis».

[ p. 585 ]
Aves del Cretácico. — En los estratos del Cretácico Superior temprano de Kansas se encuentran ocasionalmente esqueletos de excelente calidad de grandes aves acuáticas reptilianas de una especie que Marsh llamó el «ave real del oeste» (Hesperornis regalis, Figs., págs. 559 y 579). Fue en este género donde Marsh observó por primera vez dientes en las aves, y este descubrimiento lo llevó a predecir que cuando se encontrara la cabeza del Archceopteryx, también se vería que tenía dientes. El esqueleto del Hesperornis mide 1.8 metros de largo, aunque no superaba los 1.2 metros de altura; sus alas eran vestigiales e inútiles en el aire o el agua, pero sus grandes patas eran palmeadas y nadaban en el mar, por curioso que parezca, con movimientos laterales hacia afuera en lugar de hacia abajo. Pasaron la mayor parte de su vida en el mar, y en tierra firme eran más indefensos que los colimbos, encorvándose como las focas. Asociadas a estas, se encuentran muy raramente otras aves mucho más pequeñas, con alas poderosas (Ichihyornis, Fig., p. 559), que se parecían mucho a las gaviotas y charranes modernos. Ambas aves, al igual que las del Jurásico, tenían dientes pequeños y curvos y se alimentaban de carne.
Existe una gran diferencia entre las aves verdaderamente reptilianas del Jurásico Superior y las que se conocieron posteriormente, en el Cretácico tardío. Lucas afirma que estas diferencias son mayores que entre las del Cretácico y las actuales. Las más antiguas tenían largas colas de reptil con las plumas en pares a cada lado, mientras que las del Cretácico parecen haber tenido colas en abanico, como en las aves actuales. Las distintivas alas de tres dedos del Archaeopteryx están unidas en las aves del Cretácico para el soporte de las plumas, pero los cráneos de todas ellas. Las formas mesozoicas siguen siendo reptilianas, ya que los huesos no están completamente unidos como en las formas actuales.
Origen del vuelo. — Un vuelo pesado y engorroso estaba claramente presente en el Jurásico Superior (Archaeopteryx) y un vuelo excelente en aves pequeñas como Ichthyornis en el Cretácico Superior. A finales del Cretácico, las aves dentadas de gran tamaño (Hesperornis), aunque carentes de vuelo, eran grandes buceadoras en los mares de Kansas en busca de peces. Hacia el final del Mesozoico, se cree que todas las aves comenzaron a perder sus dientes y aquellas con un vuelo bien desarrollado se desplegaron en los tipos modernos desdentados, mientras que aquellas con un vuelo deficiente (pero no Hesperornis) dieron origen a las aves corredoras sin vuelo y sin dientes como los avestruces y los moas aún más grandes del hemisferio sur.
Se han propuesto dos teorías para explicar el origen del vuelo en las etapas posteriores a la fase arbórea de la evolución de las aves: la teoría de las dos alas o Archaeopteryx, y la teoría de las cuatro alas o Tetrapteryx [ p. 586 ] theory. Estas teorías se ilustran en la figura inferior. Ambas hipótesis asignan dos fases al origen del vuelo en las aves: primero, una fase primaria de vida terrestre, durante la cual se desarrollaron las características peculiares de las extremidades traseras y los pies, con sus fuertes analogías con los pies bípedos de los dinosaurios; segundo, una fase puramente arbórea que conduce al vuelo bien desarrollado.
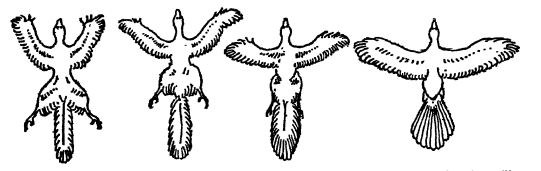

Pérdida del vuelo. — Los partidarios de ambas teorías creen que, tras la fase arborícola o voladora de las aves, se produjo, entre las formas terrestres, más grandes y, por lo tanto, más pesadas (generalmente denominadas struthious, término que hace referencia a los avestruces), una fase terrestre secundaria en la que se perdió la capacidad de volar y se reanudó la locomoción a carrera. De las aves voladoras descendieron las especies acuáticas, algunas de las cuales también perdieron las alas, como en HesperorniSy, que solo conserva un vestigio, o en los pingüinos actuales, en los que el ala se transformó en un remos. Los buceadores de los mares del Cretácico representan uno de los muchos casos en los que las formas adaptadas a la tierra se volvieron completamente acuáticas, y su transformación se desarrolló como resultado del nuevo hábitat, al que recurrieron [ p. 587 ] debido a su mayor cantidad de alimento más fácil de obtener.
Casi todos los continentes, en algún momento del Cenozoico, albergaron grandes aves terrestres similares a los avestruces. Las más altas y pesadas fueron los moas de Nueva Zelanda, exterminados por los maoríes hace cinco o seis siglos. Existían unas veinte especies, la mayor de las cuales, el Dinornis maximus, medía 3 metros de altura, 60 centímetros más que el avestruz más grande. Otra especie estrechamente relacionada, pero de menor tamaño, era el Aepyornis de Madagascar, un ave que ponía los huevos más grandes conocidos, de 23 x 33 centímetros. Fue el hallazgo de estos huevos por los primeros navegantes lo que dio lugar a las enormes exageraciones que conmueven al lector con asombro y terror en los relatos del Roc que Simbad el marino cuenta en Las mil y una noches.
Durante el Eoceno temprano, vivió en Wyoming una gigantesca ave corredora con solo vestigios de alas, conocida como Diatryma. Un ejemplar de esta especie, exhibido en el Museo Americano de Historia Natural, mide casi 2 metros de altura y muestra un cuello corto pero macizo, coronado por una cabeza tan grande como la de un caballo. Sin embargo, la ave terrestre más poderosa del Cenozoico fue el Phororhacos, hallado en la formación Pampas de Argentina, de 2,1 a 2,4 metros de altura, con un cráneo de 58 centímetros de largo, pesado y marcadamente picudo, aparentemente la más temible de las aves rapaces. No estaba emparentada con los avestruces, sino con las garzas actuales (véase la fig., pág. 686).
¶ Lectura colateral
F. A. Lucas, Animales del pasado. Museo Americano de Historia Natural, Manual, n.º 4, 1922.
F. A. Lucas, Los comienzos del vuelo, American Museum Journal, vol. 16, 1916, págs. 1-11.
W. D. Matthew y W. Granger, El esqueleto de Diairyma, un ave gigantesca del Eoceno Inferior de Wyoming. Boletín del Museo Americano de Historia Natural, vol. 37, 1917, págs. 307-326.
H. F. Osborn, El origen y la evolución de la vida. Nueva York (Scribner), 1917.
| XXXIX. Cretácico Superior y el Nacimiento de las Montañas Rocosas | Página de título | XLI. El amanecer de lo reciente en el Cenozoico |