| XLII. La evolución de los mamíferos y el auge de la mentalidad en el Cenozoico | Página de título | XLIV. La evolución de los elefantes |
[ p. 624 ]
¶ Los caballos
Al demostrar la veracidad de la evolución, los caballos, sobre todo entre todos los organismos, son la mejor ilustración del desarrollo de esta doctrina mediante la selección natural y la adaptación al medio. Son los «animales de exhibición» de la evolución, ya que su historia, que se remonta a millones de años, es bien conocida. De esta manera, la famosa Colección de la Universidad de Yale, reunida por el profesor Marsh, contribuyó en gran medida a establecer la veracidad del darwinismo (Fig. del estudio, pág. 627).
Huxley afirmó hace muchos años que el caballo debió de derivar de algún cuadrúpedo con cinco dedos completos en cada pata, y cuyos huesos del antebrazo y de la pierna estaban completos y separados. Además, si el caballo ha evolucionado de esta manera, y se han conservado los restos de las diferentes etapas de su evolución, deberían presentarnos una serie de formas en las que el número de dedos se reduce gradualmente, los huesos del antebrazo y la pierna adquieren la condición equina, y la forma y disposición de los dientes se aproximan sucesivamente a las de los caballos actuales. Desde la época de Huxley, se han encontrado casi todos los eslabones perdidos en la evolución de los caballos, y en ningún otro lugar esta historia está tan completa como en las formaciones cenozoicas de las Grandes Llanuras de Estados Unidos.
El caballo es el animal doméstico más útil y hermoso del hombre, y ha sido uno de los factores más importantes de su civilización. En sus inicios, el hombre se alimentaba de caballos, y en Solutre, en el departamento de Saône-et-Loire, Francia, se encuentra una pila de huesos de caballo que se estima representa a 80.000 individuos. Posteriormente, el caballo se convirtió en el principal medio de transporte del hombre y su bestia de carga en la agricultura y la guerra. El caballo también se encuentra entre las máquinas orgánicas de correr más perfectas y veloces, como el hombre demuestra con el caballo de carreras. Como migrantes en todos [ p. 625 ] los continentes y adaptándose a diversos entornos, desde climas tórridos hasta árticos, los caballos solo han tenido dos iguales: los elefantes y el hombre. En su estado salvaje, los caballos están ahora restringidos a las llanuras abiertas y áridas de Asia central y África, ya que los mustangs y los caballos salvajes de América del Norte y del Sur descienden de caballos domésticos que han estado en libertad desde la época de los exploradores españoles.
Caracteres distintivos. — La familia de los caballos (Equidse, de Equus caballus, el caballo viviente) incluye los caballos, cebras y asnos actuales. Pertenecen a los mamíferos ungulados de dedos impares (Perissodact 3 da), en los que el eje del pie se encuentra en el tercer dígito. Los caballos se caracterizan por sus pies muy largos y delgados, cada uno compuesto por un solo dedo funcional, el tercer dígito. El casco es el equivalente a la uña o garra del tercer dedo o dedo del pie en otros animales. Por lo tanto, los caballos caminan sobre la punta del dedo, de hecho, sobre la uña del tercer dedo, siendo la muñeca lo que los jinetes llaman la «rodilla» y el talón el «corvejón». Como el tercer dedo de cada extremidad sostiene a todo el caballo, es necesariamente mucho más grande que en los animales en los que el peso se distribuye entre varios dígitos. Sin embargo, a cada lado del dedo funcional, es decir, el «hueso de la caña», se encuentra un hueso delgado conocido como «hueso de la tablilla». Estos son los vestigios del segundo y cuarto dedo de los cinco originales en los ancestros de los caballos (Fig., pág. 627). Por curioso que parezca, los caballos domésticos a veces nacen con dos o tres dedos supernumerarios en una o más patas, y se dice que César cabalgó uno de ellos en batalla. Toda la estructura del caballo está preeminentemente adaptada a la carrera rápida, y cada parte del esqueleto se ha modificado y especializado para tal fin.
Los dientes de los caballos son tan peculiares como sus pezuñas de un solo dedo (Fig., arriba). Las muelas son prismas largos y cuadrados que crecen [ p. 626 ] desde las encías tan rápido como se desgastan en las coronas. Este desgaste se acelera debido a la sílice en las hierbas que comen y por la arena del suelo que se ingiere durante la alimentación. La superficie de molienda exhibe un patrón peculiar y complejo de bordes de esmalte duro, entre los cuales hay espacios más blandos compuestos de dentina y de un material llamado cemento, muy similar en calidad a la dentina, pero más blando y formado de una manera diferente. La dentina se forma en las superficies internas del esmalte mientras el diente aún está dentro de la mandíbula; el cemento se deposita en las superficies externas del esmalte después de que el diente ha atravesado la mandíbula y antes de que aparezca por encima de la encía.

Evolución. — La familia de los caballos se remonta a casi principios del Cenozoico, sin una sola interrupción importante. Cuando los pequeños «caballos del amanecer» de cuatro dedos (Eohippus), no más grandes que un perro pequeño, aparecieron en el oeste de Norteamérica a principios del Eoceno (Wasatch), la tierra se encontraba mucho más cerca del nivel del mar que ahora y el clima, aunque al principio con inviernos, pronto se volvió cálido y uniforme durante todo el año. Durante mucho tiempo, las estaciones fueron muy parecidas y el clima fue tropical y húmedo, lo suficientemente [ p. 627 ] [ p. 628 ] como para dar lugar a extensas áreas de bosque, al menos en las Cordilleras (montañas Laramide). Sin embargo, en las Grandes Llanuras, el clima era más seco y aquí se extendían grandes llanuras abiertas cubiertas de hierba. Con el Mioceno, todo el oeste de Norteamérica comenzó a elevarse, alcanzando su punto máximo en el Pleistoceno. Junto con estos cambios topográficos, el clima se volvió más frío, más seco y, finalmente, gélido. A todos estos cambios en el medio ambiente, los caballos se adaptaron o migraron a hábitats más favorables, y al hacerlo cambiaron de las formas más pequeñas, con muchos dedos, a las más grandes, con menos dedos, más rápidas y más inteligentes.
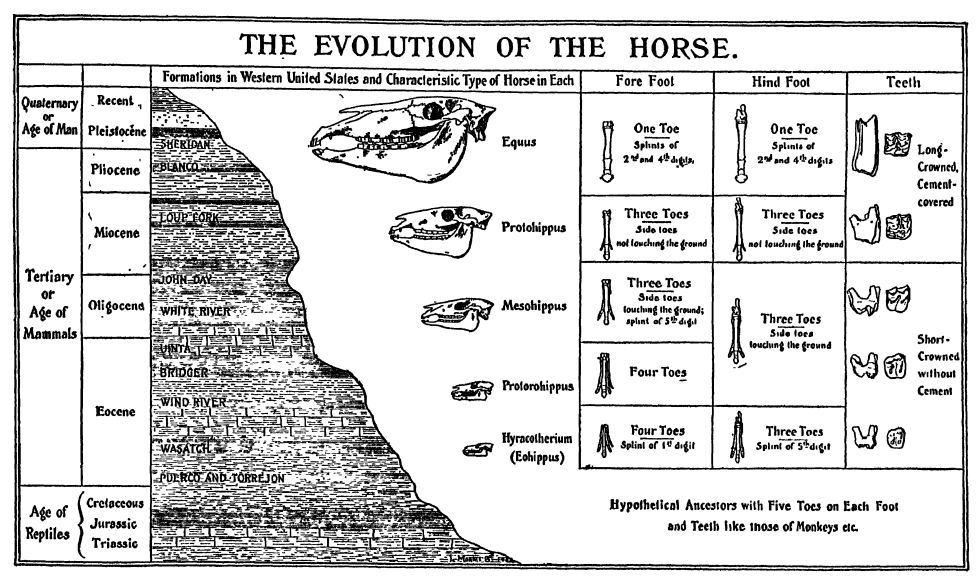
Al principio, los caballos de múltiples dedos pastaban en los bosques, donde eran presa fácil de los carnívoros de la época. Sin embargo, con la disminución de los bosques y la aparición de llanuras herbosas más secas, se extendieron en busca de protección a las llanuras abiertas, donde desarrollaron una velocidad cada vez mayor. Con el alargamiento de la parte inferior de las extremidades y el desarrollo del hábito de correr rápidamente sobre las puntas de los pies, se produjo la pérdida gradual, por desuso, de los dedos adicionales, y un cambio igualmente notable en la dentadura, de corona corta para ramonear a corona larga (para pastar). De esta manera, los caballos se convirtieron en uno de los animales más especializados, adaptados a un entorno particular.
Se desconoce el origen de la familia de los caballos. Los «caballos del amanecer» aparecen aproximadamente al mismo tiempo y en el mismo estado de evolución, en Europa occidental y Norteamérica. En Europa se extinguieron rápidamente (Eoceno), pero Norteamérica fue su centro de desarrollo durante el Cenozoico. Curiosamente, aunque los caballos estuvieron presentes durante todo el Pleistoceno, tanto en Norteamérica como en Sudamérica, todos se extinguieron en algún momento antes de la llegada del hombre rojo. Nuestros caballos salvajes actuales son asilvestrados, es decir, tuvieron ancestros domesticados, y los de Asia, África y Europa son descendientes de los caballos del Mioceno temprano que se extendieron desde Norteamérica hasta Siberia a través de Alaska. A finales del Mioceno, los caballos norteamericanos se extendieron a Sudamérica.
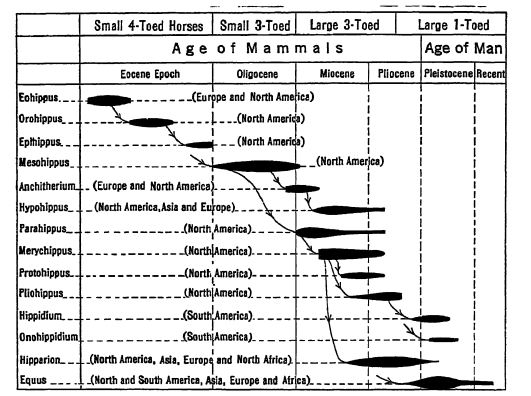
Mateo reconoce doce etapas en la evolución de la familia de los caballos, presentes en otras tantas formaciones geológicas diferentes y sucesivas. Además de la línea principal de descendencia que dio origen a los caballos, asnos y cebras modernos, hubo varias ramas colaterales que no dejaron descendencia.
Scott resume el largo y maravilloso desarrollo de la familia de los caballos de la siguiente manera: (1) En tamaño hubo un aumento algo fluctuante, que condujo por lentas gradaciones desde los diminutos caballos del Eoceno Inferior, aproximadamente del tamaño de un fox-terrier, hasta los grandes animales [ p. 629 ] del Pleistoceno, de aproximadamente 14 manos de altura. (2) Los molares, originalmente bajos, cuspideantes y con raíces, cambiaron gradualmente del tipo ramoneo a los dientes de pastoreo muy largos, prismáticos, complejos, y las mandíbulas inferiores crecieron en profundidad para acomodar esta elongación. (3) La cara se volvió relativamente más larga y los ojos se desplazaron progresivamente más hacia atrás. (4) El cuello corto se alargó mucho y las vértebras individuales se modificaron para dar flexibilidad sin pérdida de fuerza. (5) Las extremidades se alargaron mucho más, los huesos del antebrazo y la parte inferior de la pierna se fusionaron, el del lado interno (radio y tibia) se agrandó para soportar todo el peso, y el externo (cúbito y peroné) se atrofió más o menos. (6) Los pies se alargaron mucho y el dedo medio o tercero de cada uno se agrandó gradualmente hasta soportar todo el peso, modificando al mismo tiempo la forma del casco para que fuera el único soporte del cuerpo. Los demás dedos se redujeron gradualmente y se volvieron infuncionales, aunque a menudo se conservaban como férulas. Primero se perdió el primer dedo, luego el quinto, luego el segundo y el cuarto se redujeron a espolones y finalmente a férulas. Así, los caballos pentadactilos del Eoceno Inferior se transformaron en las especies monodactilas del Plioceno y el Pleistoceno.
Cerebro y Mentalidad. — El cerebro de los caballos actuales es grande y complejo, lo que implica una gran inteligencia, pero no es igual al del elefante. La docilidad del caballo y su capacidad de aprendizaje son notables. Por otro lado, es emocional, y su psicología está en gran medida ligada a su modo de defensa habitual —la huida—, ya que el primer impulso de un caballo doméstico al ver algo incomprensible es huir. En estado salvaje, este mismo impulso es de gran ayuda como medio de supervivencia. (Lull.)
Antepasado de los caballos. — Cuando el gran paleontólogo inglés, Richard Owen, describió al pequeño «caballo del amanecer» de cinco dedos de Europa (Hyracotherium), un tipo estrechamente relacionado con el «caballo del amanecer» americano (Eohippus), desconocía en absoluto que poseía el linaje ancestral de los caballos, tan diferente era de la forma moderna. Con el posterior hallazgo en Norteamérica de etapas de desarrollo entre este y otros caballos, se hizo evidente que el Hyracotherium era una forma generalizada que descendía de un linaje que dio origen no solo a los caballos, sino también a los tapires y rinocerontes. En esto se ve cómo un linaje ancestral que se despliega en diferentes entornos conduce eventualmente a animales que se ven muy diferentes entre sí y, sin embargo, tienen dentro de sus cuerpos estructuras que muestran sus relaciones. «La conclusión es inevitable: el caballo, el rinoceronte y el tapir, tres razas [ p. 630 ] ampliamente diferentes hoy en día, se derivan a través de cambios progresivos de un tipo ancestral común» (Matthew).
Sucesión geológica. — El miembro más antiguo conocido de la familia EquidjB fue el «caballo del alba», que pululaba en los bosques y claros del Eoceno Inferior (Wasatch). Eran pequeñas y gráciles criaturas, no más grandes que un perro, o de unos 28 cm de altura a la cruz. Tenían en las patas delanteras cuatro dedos funcionales y el vestigio de otro, mientras que las patas traseras tenían tres dedos y dos diminutas férulas, vestigios del primero y el quinto dedo. Esta evidencia indica claramente que el Eohippus se originó en ancestros con pies pentadáctilos. Los molares eran cuspideados y cortos, propios de la alimentación por ramoneo; el cuello era muy corto, el cuerpo largo, con el dorso arqueado, las extremidades y los pies cortos, y las extremidades traseras mucho más largas que las delanteras (Fig., p. 626).
Los caballos del Oligoceno tuvieron un desarrollo intermedio entre los del Eoceno y el Mioceno. En el Oligoceno Inferior (Río Blanco), los más grandes eran del tamaño de una oveja (Mesokippus), y si bien sus dientes eran de corona baja y de tipo ramoneador, estaban evolucionando hacia los de un animal de pastoreo. Las patas delanteras tenían tres dedos funcionales, y aunque el central era el más grande, los dos dedos laterales tocaban el suelo; las patas traseras también tenían tres dedos, pero no presentaban férulas (Fig., p. 627).
En el Mioceno Medio, los caballos de tres dedos seguían siendo mayoritariamente ramoneadores, aunque algunos de los más progresivos se transformaban rápidamente hacia los herbívoros. Es aquí donde encontramos las formas intermedias (Merychippus) [ p. 631 ] y observamos la división de la familia de los caballos en varios filos que se desarrollaron independientemente. El Plioceno Inferior presentó tipos tanto progresivos como conservadores de estos animales. En el Plioceno Medio, entre las manadas de caballos que vagaban por las llanuras, existían tres filos de caballos de pastoreo de tres dedos, pero los dedos laterales ya no tocaban el suelo; si bien eran «espolones», en ocasiones aún tenían cierta utilidad de apoyo. En el Plioceno Superior se encontraron los últimos caballos de pastoreo y ramoneo de tres dedos, y aquí también vivió el primer miembro de un solo dedo del género Equus. En la primera mitad del Pleistoceno existían al menos diez especies del mismo género, entre ellas caballos más grandes que cualquier otro caballo vivo actual. (Estudio Fig., pág. 630.)
[ p. 632 ]
¶ Los camellos
La familia de los camélidos incluye ahora las dos especies de camellos de las zonas desérticas de Asia central y los uamas y guanacos de las zonas más altas y frías de Sudamérica. Estos extraños animales tuvieron su origen y evolución esencial en el oeste de Norteamérica, y su historia corre paralela a la de los caballos. Representan adaptaciones a climas templados y llanuras arenosas abiertas. Los camellos pertenecen a la división de los mamíferos ungulados de dedos pares, los artiodáctilos. En estos casos, el eje del pie se sitúa entre el tercer y el cuarto dedo, y no, como en los caballos, en el tercero.
Los camellos aparecieron por primera vez en el Eoceno Superior con un tamaño similar al de un gato doméstico, y posteriormente se extendieron a lo largo del Cenozoico. En el Mioceno Inferior, los camellos, al igual que los caballos, comenzaron a diversificarse, y los camellos grandes aún abundaban en Norteamérica durante la primera mitad del Pleistoceno, pero poco después se extinguieron. Las llamas se extendieron hacia el sur a través de los trópicos hasta Sudamérica en el Plioceno, y al mismo tiempo, la población de camellos se expandió hacia el norte y finalmente cruzó el puente terrestre entre Alaska y Siberia hacia Asia y, de allí, hacia África.
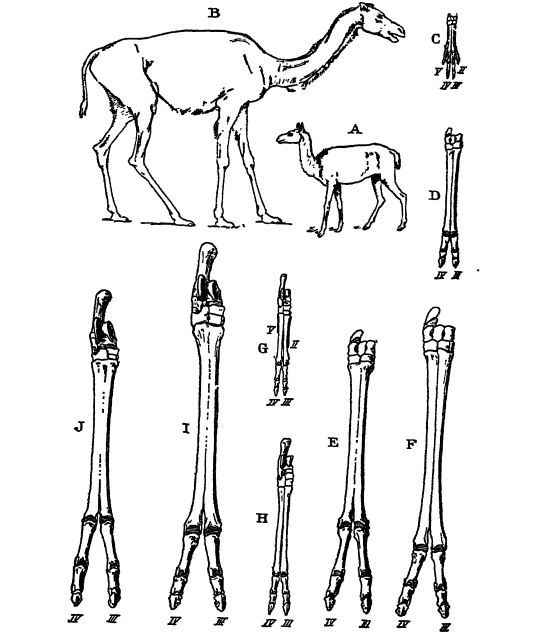
Scott afirma que el modo de evolución de los camellos no difiere significativamente del observado en los caballos. Se observó el mismo aumento en la estatura corporal y en la longitud relativa de las extremidades y los pies, una disminución en el número de dedos de los cinco originales a dos en los camellos (Fig., pág. 631), y a uno en los caballos (Fig., pág. 627), y un desarrollo similar de los dientes de corona alta para moler a partir del tipo de ramoneo de corona baja. Las dos familias de camellos y caballos surgieron en una sola serie.
¶ Los ambípodos de patas cortas
Entre los mamíferos arcaicos del Eoceno existía una reserva de ramoneadores, de hábitos lentos y esqueletos torpes, pero abundantemente representados por especies y géneros. Estos se denominan Amblypoda, nombre que hace referencia a sus pies cortos o romos. Sus patas eran robustas y con forma de pilar, y los pies cortos tenían cinco dedos con almohadillas elásticas, como los de los elefantes. Como todos los mamíferos arcaicos, su cerebro era pequeño en proporción al tamaño de su cuerpo.
Los amblipodos probablemente se originaron en Norteamérica a finales del Mesozoico, ya que las formas más antiguas se conocen en el Paleoceno. Eran comunes en Wasatch, crecieron cada vez más y se diversificaron durante finales del Eoceno, y se extinguieron [ p. 633 ] antes del final de esta época en Bridger. Son mamíferos muy llamativos y característicos del Eoceno norteamericano, y solo unos pocos (Coryphodon) migraron a Europa occidental.
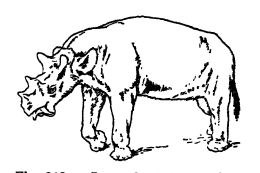
Los más llamativos de los anfípodos fueron los uintatheres, llamados así por el género Uintatherium (ver Fig., abajo), encontrados en las montañas Uinta de Utah, muchos de los tipos crecieron hasta el tamaño de pequeños elefantes, 7 pies de alto en los hombros, y en la forma general de sus piernas y pies anchos, pero no en sus cabezas, se parecían a los proboscidios. Los uintatheres, dice Scott, eran verdaderos gigantes y los animales más fantásticos de su tiempo. Una vez fueron considerados como proboscidios, pero ahora se ha establecido que los dos grupos no están de ninguna manera relacionados directamente. Eran, sin embargo, los animales parecidos a los elefantes del Eoceno norteamericano.
Las cabezas de los uintatheres eran grandes y multicorneales, y, curiosamente, la parte superior de sus cráneos era cóncava o con forma de cuenca, un rasgo característico de ellos. De los cuernos auténticos revestidos de cuerno, como en los rinocerontes, había un par sobre la nariz, y sobre los ojos y las orejas había otros pares de protuberancias óseas más o menos altas que se cree que estaban cubiertas únicamente por piel gruesa. Otra característica sorprendente, pero restringida a los machos, era el desarrollo de los caninos superiores en dos formidables colmillos curvos y afilados como sables. Para evitar que estos colmillos se rompieran durante el uso, la mandíbula inferior desarrolló amplios rebordes óseos para su protección. Se desconoce su uso. En ambos sexos, el juego superior de incisivos estaba suprimido, como en muchos rumiantes actuales. El cerebro de los uintatheres era «absurdamente pequeño» y su baja mentalidad entre los mamíferos es comparable a la de los dinosaurios entre los reptiles. Puede haber sido la causa principal de su extinción, ya que tuvieron que competir con las hordas cada vez más numerosas de mamíferos modernizados con cerebros más grandes y, por lo tanto, más alertas.
¶ Las bestias gigantes o titanotheres
En el Eoceno Inferior, entre los mamíferos modernizados inmigrantes apareció un imgulado de dedos impares (Eotitanops), más pequeño que una oveja y con una apariencia similar a la de un tapir. Esta forma ancestral, según Osborn, evolucionó en once [ p. 634 ] ramas principales, comenzando su desarrollo en el Bridger, continuando en el Uinta superior y desapareciendo en el punto culminante de su desarrollo, a principios del Oligoceno (Río Blanco). Por lo tanto, la evolución más significativa de estos animales ocurre a menos de 60 metros de los estratos del Oligoceno. A finales del Eoceno se extendieron a Mongolia (Protitanotherium), pero aparentemente no vivieron allí mucho tiempo.
Estos son los titanoterios, «bestias gigantes», mamíferos muy característicos del Cenozoico norteamericano. Solo las formas posteriores alcanzan el tamaño de pequeños elefantes, pero Brovtotherium, aparte de los elefantes, es «el producto más imponente de la evolución Tna.rmna.1ia.n». Se desconoce por qué los titanoterios fracasaron en su mejor momento, pero es posible que las condiciones climáticas más secas del Mioceno y la transición de sus hábitats de ramoneo forestal a pastizales abiertos ya hubieran comenzado, y de ser así, podrían haber sido la causa de su desaparición (véase la figura anterior).
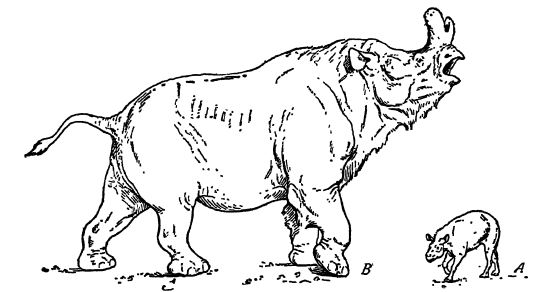
Los titanotheres eran corpulentos, con patas columnares y pies cortos, estos últimos apoyados sobre almohadillas gruesas como las de los elefantes. En todos ellos, las patas delanteras tenían cuatro dedos y las traseras tres, y en las formas más antiguas y pequeñas, los dedos y las pezuñas eran más prominentes. Su rasgo más característico reside en la evolución de la cabeza. En los más antiguos, el cráneo era pequeño, largo y estrecho, y carecía de protuberancias. Sin embargo, los caninos eran prominentes como colmillos y los incisivos se utilizaban para ramonear la vegetación. Evidentemente, los colmillos en las formas primitivas se utilizaban como [ p. 635 ] armas defensivas. En el Uinta, Scott sa 3 rs, los titanotheres eran más grandes y presentaban pequeñas protuberancias sobre los ojos que, con el tiempo, se agrandaron y se desplazaron hacia adelante, hasta que en el río White estos cuernos óseos alcanzaron gran tamaño y se situaron en la nariz. Mientras los cuernos se agrandaban, el cráneo se modificaba para soportar su peso y resistir mejor el impacto al usarlos. Cuando los cuernos se convirtieron en armas, los colmillos se redujeron a la insignificancia y los dientes frontales dejaron de usarse para cortar, ya que la lengua y el labio superior se encargaban de ello. En la plenitud del desarrollo del titanoterio, las cabezas eran largas, muy anchas, grandes y macizas; el perfil, profundamente cóncavo, se asemejaba al de algún fantástico rinoceronte con cuernos nasales. Tanto el cuerpo como las extremidades de estas formas derivadas se asemejaban a los grandes rinocerontes.
El cerebro de la especie más grande era muy pequeño, no más grande que el puño de un hombre, lo que indica que estas grandes bestias «debieron haber sido increíblemente torpes y estúpidas, superando incluso a los rinocerontes modernos en este aspecto» (Scott).
¶ Los rinocerontes
Los rinocerontes pertenecen a los mamíferos ungulados de dedos impares; generalmente tienen tres dedos y piel típicamente gruesa. Por lo general, tienen poco pelo, aunque existió una especie lanuda contemporánea al hombre durante el Pleistoceno en el clima frío de Siberia y el norte de Europa (Ceratotherium antiquitatis). Son ramoneadores y pastores, y viven en bosques, estepas y pantanos. Tienen cabezas grandes, cuellos cortos y cuerpos muy largos y macizos, y sus extremidades son cortas, robustas y columnares, como las de los elefantes. Las formas actuales miden entre 1,20 y 1,90 metros de altura a los hombros, y las especies de un solo cuerno se encuentran en India y Java, mientras que las que tienen dos cuernos en tándem, uno en la nariz y otro en la frente, viven en África y Sumatra. Ninguna de las formas fósiles americanas era tan alta ni tan pesada como la forma actual de mayor tamaño. Los cuernos de los rinocerontes tienen la peculiaridad de que no son huecos como los del ganado vacuno ni de hueso, sino que son crecimientos dérmicos sólidos compuestos de pelos aglutinados, por lo que nunca se encuentran fósiles. Sin embargo, su presencia en formas fósiles siempre se indica por huesos nasales engrosados y rugosos.
Desde el Plioceno medio no ha habido rinocerontes en América del Norte, y sin embargo este continente puede haber sido no solo el lugar de su origen, sino también el de su evolución más significativa. El origen y desarrollo de las formas ancestrales en América del Norte y más tarde de los verdaderos rinocerontes del Viejo Mundo es una historia muy compleja, mucho más que la de los caballos y los titanoterios. Los Tnfl.TnTna1 similares a los ehinoceros aparecieron en América a principios del Eoceno [ p. 636 ] en formas pequeñas, activas y generalizadas (Hyradiyvs, Fig., p. 638) que en el curso del Cenozoico se desplegaron en al menos ocho ramas. Estas se agrupan nuevamente en cuatro líneas principales de evolución, a saber, (1) el stock ancestral de formas pequeñas, indefensas y sin cuernos, similares a los rinocerontes; (2) un tipo acuático especializado, grande y de vida corta que vivió en ríos y lagos; (3) los gigantescos baluchitheres de Asia; y (4) el tronco principal de los rinocerontes auténticos, enormes, de movimientos lentos y generalmente con cuerpo. En el Mioceno y el Plioceno, estos fl.niTna.1 alcanzaron su distribución más amplia, habitando entonces en todos los continentes excepto Australia y Sudamérica. Los troncos ancestrales y acuáticos se extinguieron en Norteamérica durante el Oligoceno, y todos ellos a principios del Plioceno.

Las formas sin cuernos surgieron primero, y posteriormente, en diferentes momentos del Cenozoico, aparecieron las con cuernos. Los cuernos pueden ser simples, dobles en un par transversal (= con cuernos en par), o dispuestos uno tras otro a lo largo de la línea media de la cabeza (= con cuernos en tándem); generalmente se ubican sobre la nariz, pero en algunos casos el cuerno se encuentra en la frente. La presencia de cuernos, como en los titanoterios, provocó cambios en la forma del cráneo (véanse las figuras anteriores y la pág. 637).
El mamífero terrestre más grande descubierto hasta la fecha fue un rinoceronte sin cuernos conocido como Bdiichitherium. [ p. 637 ] Vivió a finales del Oligoceno o principios del Mioceno, y se encontró por primera vez en Baluchistán, aunque desde entonces se ha recolectado en Turkestán y Mongolia. Osborn afirma que medía unos 4 metros de altura hasta los hombros y unos 7,6 metros de largo; en comparación, todos los demás rinocerontes son pequeños. El cuello era largo y parecido al de un caballo, la cabeza larga y estrecha, de unos 1,5 metros de largo, con dos poderosos colmillos. Las extremidades eran muy largas y zancudas, lo que les permitía ramonear entre el follaje de los árboles.
Scott afirma que existen siete ramas de rinocerontes auténticos, tres de las cuales tienen representantes vivos. Parece que estos llamativos animales se originaron en el género norteamericano Trigonias, del bajo río White, una forma sin cuernos con cuatro dedos en las patas delanteras en lugar de tres, como en todos los demás rinocerontes. Otro rinoceronte sin cuernos de la misma época fue el CcenopuS, tridáctilo en todas las patas, y este tipo persistió hasta el Plioceno. Entre los rinocerontes con cuernos, destacan los pequeños diceratheres (con dos cuernos), que comenzaron en el Oligoceno y persistieron hasta el Mioceno (Fig., p. 636). También migraron a Eurasia y se caracterizan por un par de cuernos transversales en la nariz. Originariamente del Viejo Mundo, llegó a Norteamérica, en el Mioceno Medio, el Teleoceras, un tipo acuático con un pequeño cuerno en la nariz y patas grotescamente cortas, de modo que el vientre casi tocaba el suelo (Fig., arriba). Esta especie persistió en gran número hasta el Plioceno inferior.

En el Mioceno Superior y el Plioceno Inferior de este continente, existían al menos cuatro tipos distintos de rinocerontes que [ p. 638 ] vivían en gran abundancia, pero ninguno alcanzó el tamaño ni desarrolló los grandes cuernos de las especies actuales. Se desconoce por qué no sobrevivieron posteriormente y por qué el rinoceronte lanudo de Eurasia no llegó a Norteamérica junto con el mamut.
Los hiracodontos son los animales ancestrales sin cuernos de los que provienen todos los rinocerontes. El más antiguo se encuentra en el Eoceno (Bridger) y todos se extinguieron en el Oligoceno (Río Blanco). Hyrackyus (Fig., abajo), el género más antiguo, está muy generalizado, y de él también podrían haberse desarrollado los caballos y los titanoterios. Era aproximadamente tan grande como una oveja, pero más pesado. Los hiracodontos eran de complexión ligera, con cabezas pesadas y toscas, pero cuellos y extremidades delgados y largos; evocan más a los caballos que a los rinocerontes. De hecho, las patas de las formas del Río Blanco se asemejan a las de los caballos de la misma época (Mesohippus). Para su seguridad dependían de la velocidad, por lo que se les llama rinocerontes de carrera. Scott afirma que es interesante reflexionar sobre el hecho de que, si los hiracodontos del Río Blanco y los tapires ancestrales hubieran sobrevivido hasta la actualidad, habrían desarrollado, en el curso de su evolución, pies unidigitales, como los caballos. En los hiracodontos, al igual que en los caballos, la evolución se centró principalmente en el alargamiento del cuello, las extremidades y los pies, y en el agrandamiento de los dedos medios. Con el aumento de tamaño corporal a lo largo de su existencia, las extremidades se volvieron más robustas, pero todas eran del tipo cursorial o corredor. Solo en un género se desarrolló un par de pequeños cuernos nasales (ColonocercLs).
Los amynodontes eran rinocerontes acuáticos especializados sin cuernos. Probablemente se originaron en Hyrachyus a finales del período Bridger y se extinguieron en el Oligoceno. Amynodon era más pequeño y ligero que el posterior y más grande Metamynodon del río White. Según Scott, este último fue la criatura más pesada y masiva de su época. Su cabeza era grande, [ p. 639 ] deprimida y ancha, con las fosas nasales altas en la cabeza, acorde con sus hábitos acuáticos; el cuello corto; el cuerpo largo y macizo; y las extremidades cortas y robustas, con cuatro dedos en las patas delanteras y tres en las traseras.
¶ Lectura colateral
R. S. Lull, La evolución de la familia de los caballos. American Journal of Science, 4.ª serie, vol. 23, 1907, págs. 161-182.
W. D. Matthew y S. H. Chubb, Evolución del caballo. Museo Americano de Historia Natural, Serie de folletos guía, n.º 36, 1913.
H. F. Osborn, La era de los mamíferos. Nueva York (Macmillan’), 1910.
H. F. Osborn, El rinoceronte gigante extinto Baluchitherium de Asia occidental y central. Natural History", Vol. 23, 1923, págs. 209-22S.
W. B. Scott, Historia de los mamíferos terrestres en el hemisferio occidental. Nueva York (Macmillan), 1913.
| XLII. La evolución de los mamíferos y el auge de la mentalidad en el Cenozoico | Página de título | XLIV. La evolución de los elefantes |