[ pág. 49 ]
Ascendencia de los simios antropoides — Clima, bosques y vida en Europa occidental durante el Pleistoceno — Transición al Pleistoceno, o era del hombre — La primera glaciación, sus efectos sobre el clima, los bosques y la vida animal — La raza prehumana Trinil de Java — Los eolitos o pedernales primitivos — La segunda glaciación — Heidelberg, la raza humana más antigua conocida — La tercera glaciación
Los ancestros parcialmente conocidos de los simios antropoides y los ancestros desconocidos del hombre probablemente se originaron entre los bosques y llanuras aluviales del sur de Asia y pronto comenzaron a migrar hacia el oeste, hacia el norte de África y Europa occidental.
Ya en el Oligoceno, un precursor de los grandes simios (Propliopithecus), muy similar a los gibones, apareció en el desierto que bordea el Fayum, al norte de Egipto. A principios del Mioceno, los gibones arborícolas se expandieron hacia Europa y continuaron durante el Plioceno en las formas conocidas como Pliopithecus y Pliohylobates, siendo este último un gibón auténtico por sus proporciones; se extendió hacia el norte hasta la actual región de Alemania. Otro simio que llegó temprano a Europa fue el Dryopithecus; se encuentra en el Mioceno en el sur de Francia; sus dientes afilados recuerdan a los del orangután, la mandíbula es profunda y, en cierto modo, se asemeja a la del hombre de Piltdown. Un tercer simio (Neopithecus) se encuentra en el Plioceno Inferior cerca de Eppelsheim, Alemania, y se conoce únicamente por un molar inferior, que recuerda la dentición de Dryopithecus y, más remotamente, la de Homo. En el Plioceno, en las colinas Siwalik de Asia, se encuentra Paleopithecus, una forma generalizada que se cree emparentada con el chimpancé, el gorila y el gibón; los premolares superiores se asemejan a los del hombre.
Ninguno de estos antropoides fósiles, ni de Europa ni de Asia, puede considerarse ancestro del hombre, aunque tanto Neopithecus [ p. 50 ] como Dryopithecus han sido ubicados en o cerca de la línea de ascendencia humana por autoridades de la talla de Branco y Gaudry. Cuando Lartet descubrió Dryopithecus, Gaudry1 lo consideró, con diferencia, el simio más parecido al hombre, atribuyéndole incluso inteligencia suficiente para trabajar el sílex. Sin embargo, un conocimiento más completo de este animal ha demostrado que algunos antropoides actuales son más parecidos al hombre que Dryopithecus. Este animal está estrechamente emparentado con el linaje ancestral del chimpancé, el gorila y el orangután. La mandíbula, es cierto, se parece a la del hombre de Piltdown (Eoanthropus), pero los dientes que rechinan son mucho más primitivos y hay pocos motivos para pensar que sean ancestrales de cualquier tipo humano.[1]

[ pág. 51 ]
Entre estos antropoides fósiles, así como entre las cuatro formas vivientes, no descubrimos evidencia de parentesco directo con el hombre, pero sí evidencia contundente de descendencia del mismo linaje ancestral. Estas pruebas de ascendencia común, ya observadas en las razas humanas actuales, se hacen mucho más evidentes en las razas paleolíticas antiguas; de hecho, no podemos interpretar la anatomía de los hombres de la Edad de Piedra Antigua sin un estudio de las características principales de los simios antropoides actuales: el gibón, el orangután, el chimpancé y el gorila.

El gibón es el simio vivo más primitivo en cuanto a cráneo y dentición, pero el más especializado en la longitud de sus brazos y otras adaptaciones extremas a la vida arbórea. Al igual que en los demás antropoides, el rostro es abreviado, la región nasal es estrecha (catarro) y la caja craneana es ensanchada, pero la parte superior del cráneo es lisa y la frente carece de las crestas prominentes sobre las órbitas; por lo tanto, el perfil del cráneo del gibón (Fig. i6) es más humano que el de los demás simios antropoides. En el suelo, el gibón camina erguido, lo que le permite el libre uso de sus brazos y el movimiento independiente de sus dedos. En el cerebro se observa un notable desarrollo de los centros de la vista, el tacto y el oído. Son estas características del gibón moderno las que preservan con cambios relativamente leves el tipo del ancestro original del hombre, como lo señaló Elliot Smith.2

Las extremidades del orangután son menos alargadas y menos especializadas para la vida arbórea que las del gibón, pero más que las del chimpancé y el gorila. El cráneo es redondeado y de gran altura vertical, con amplias crestas óseas sobre las órbitas y una gran cresta media en la parte superior del cráneo en los machos mayores. La mandíbula inferior del orangután es robusta y profunda, y, aunque se utiliza como arma de combate, los colmillos caninos son mucho menos prominentes que los del gibón, el chimpancé o el gorila.
En el chimpancé observamos las prominentes crestas óseas sobre los ojos, similares a las de los humanos de las razas Trinil y Neandertal. De todos los simios antropoides, la mandíbula inferior del chimpancé es la que más se asemeja a la del hombre de Piltdown. Las hileras de dientes prognáticas o salientes y el mentón retraído sugieren las de las razas Heidelberg, Piltdown y Neandertal. Cuando el chimpancé camina (Fig. 18), los brazos llegan por debajo de las rodillas, mientras que en las razas superiores del hombre solo llegan hasta la mitad de los muslos.

[ pág. 54 ]

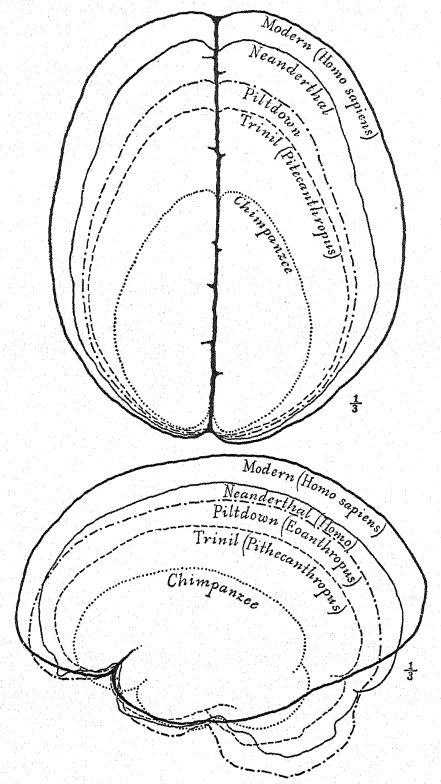
Del tronco ancestral desconocido de los simios antropoides y del hombre, el gibón fue el primero en ramificarse en el Oligoceno; el orangután se ramificó posteriormente en una dirección muy distinta. El tronco del chimpancé y del gorila se ramificó en una fecha más reciente y está más estrechamente emparentado con el del hombre. Se han encontrado cinco razas humanas primitivas en Europa en las épocas glacial o pleistocena, pero no se han encontrado rastros de otros primates, excepto los macacos, emparentados con la división inferior de los babuinos, en Europa en el Pleistoceno. Modificado según Gregory. (Para el último descubrimiento, véase el Apéndice, Nota VII).
Así, la extremidad anterior, aunque mucho más corta que la del gibón, es relativamente más larga que la de cualquier raza humana, reciente o antigua. Observamos también en el chimpancé caminante (Fig. 18) que la parte superior de la pierna, el fémur, es relativamente larga, mientras que la parte inferior, la tibia, es relativamente corta. De hecho, tanto en el brazo como en la pierna, los huesos superiores son relativamente largos y los inferiores, relativamente cortos. Estas proporciones, herencia de la vida arbórea, contrastan marcadamente con las observadas en los brazos y piernas de la raza humana neandertal, en la que las extremidades son de tipo terrestre o caminante.

Observamos también en el chimpancé un contraste entre la capacidad de agarre del dedo gordo del pie, que es una especie de pulgar, y la falta de dicha capacidad en la mano, donde el pulgar es prácticamente inútil; en todos los simios, esta función es característica del pie; en el hombre, solo de la mano. El pulgar oponible, con su capacidad de unir el pulgar con cada uno de los dedos, es la única característica ausente en todos los simios antropoides y que se desarrolló tempranamente entre los ancestros del hombre.
El cráneo del chimpancé es más largo que el del orangután. La característica más prominente en la vista superior es la protuberancia extrema de las órbitas, rodeadas por una cresta ósea supraorbitaria y cirreumorbitaria. Esta cresta también está muy desarrollada en el cráneo de neandertal, así como en el de Pithecanthropus o Trinil, pero, hasta donde sabemos, está completamente ausente en el de Piltdown. Al igual que en el orangután y el gorila, se desarrolla una cresta a lo largo de la parte media superior del cráneo para la inserción de los poderosos músculos de las mandíbulas, una cresta completamente ausente en el gibón y probablemente ausente en todos los verdaderos ancestros del hombre.
El gorila ilustra a la perfección las especializaciones que se inician en el chimpancé, atribuibles a una vida en parte arbórea y en parte terrestre, en la que el cráneo y las mandíbulas se utilizan como poderosos órganos de combate. La cabeza se alarga debido al crecimiento del hocico hacia adelante, lo que le confiere un prognatismo extremo. Las extremidades y el cuerpo del gorila se alejan del tipo primitivo de simios arbóreos, de extremidades delgadas, y están parcialmente adaptados a un hábito bípedo y terrestre.
En cuanto a la evolución psíquica,3 Elliot Smith observa que el modo de vida arbóreo de los primeros ancestros del hombre desarrolló movimientos rápidos, alertas y ágiles que estimularon el desarrollo progresivo de las porciones posterior y lateral del cerebro. El sentido del olfato se había desarrollado bien en una vida terrestre previa, pero una vez que estas criaturas abandonaron la tierra y se trasladaron a los árboles, la guía del olfato se volvió menos esencial, ya que la vida entre las ramas de los árboles favorece el desarrollo de los sentidos de la vista, el tacto y el oído. Además, exige una agilidad y rapidez de movimiento que requieren centros motores eficientes en el cerebro para coordinar y controlar las acciones que requiere la vida arbórea. La especialización de la vista despierta la curiosidad para examinar los objetos con mayor minuciosidad y guía las manos hacia movimientos más precisos y hábiles.
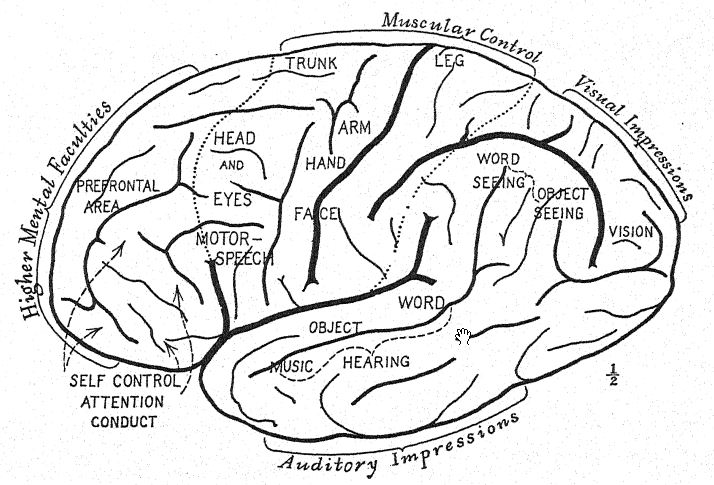
La anatomía del hombre está llena de recordatorios remotos de esta existencia arbórea original, lo que explica también el desarrollo muy grande y temprano de las porciones posteriores del cerebro, en las que se encuentran los diversos sentidos de la vista, el tacto y el oído.
El primer paso de la vida arbórea a la terrestre se caracteriza por la capacidad de caminar más o menos erguido sobre las extremidades traseras, liberando así los brazos. Esta capacidad se desarrolla en mayor o menor grado en todos los simios antropoides; con la práctica, se convierten en caminantes expertos. La mayor libertad que la postura erguida otorga a los brazos, a los movimientos de las manos y a los movimientos separados de los dedos es especialmente notable en el gibón. El desarrollo de las facultades manuales influye en el mayor crecimiento y especialización del cerebro; por lo tanto, el cerebro y la postura erguida se complementan. En el gibón se observa un marcado aumento del tamaño de las porciones cerebrales que irrigan los centros del tacto, la visión y la audición.
El debate sobre cómo se formaron los ancestros del hombre se ha centrado principalmente en las afirmaciones rivales de cuatro líneas de evolución estructural: primero, la asunción de la actitud erguida; segundo, el desarrollo del pulgar oponible; tercero, el crecimiento del cerebro; y cuarto, la adquisición de la capacidad del habla. El argumento a favor de la actitud erguida sugerido por Lamarck, y hábilmente expuesto por Munro4 en 1893, indica que el cultivo de la habilidad con las manos y los dedos es la base de la supremacía mental del hombre. El argumento de Elliot Smith de que el crecimiento constante y la especialización del propio cerebro han sido el factor principal que ha guiado a los ancestros del hombre paso a paso hacia arriba indica que un avance como la actitud erguida se produjo porque el cerebro había posibilitado los movimientos hábiles de las manos.
La verdadera concepción de la evolución prehumana, que tuvo lugar durante el Mioceno y el Plioceno, se basa más bien en el desarrollo coincidente de estas cuatro facultades distintivamente humanas. De las proporciones de las extremidades en la raza neandertal, se desprende que la postura parcialmente erguida y la marcha se dieron mucho antes en la era geológica de lo que imaginábamos. La estrecha relación entre el uso del pulgar oponible y el desarrollo de las facultades mentales superiores del hombre se sustenta hoy en día en el descubrimiento de que uno de los mejores métodos para desarrollar la mente del niño es insistir en el uso constante de las manos, ya que se ha comprobado que la acción y la reacción entre la mano y el cerebro desarrollan la mente. Una acción y reacción similar entre el pie y el cerebro desarrolló la marcha erguida que liberó a la mano de su función locomotora y de agarre de las extremidades y, mediante el perfeccionamiento resultante del movimiento de los pulgares y los dedos, convirtió la mano en un órgano listo para la creciente especialización que exigía la fabricación de herramientas de pedernal.
Creemos que esta es la etapa alcanzada a finales del Plioceno, cuando el ancestro humano emerge de la era de los mamíferos y entra en la era del hombre, el período en el que comienza propiamente la prehistoria del hombre. La postura es erguida, la mano posee un pulgar oponible bien desarrollado, y los centros cerebrales relacionados con los sentidos superiores y con el control de todos los movimientos de las extremidades, manos y dedos están bien desarrollados. La capacidad del habla puede ser aún rudimentaria. Los centros cerebrales anteriores, encargados del almacenamiento de experiencias y el desarrollo de ideas, son ciertamente muy rudimentarios.
¶ Cambio de entorno en Europa
Considerando que el origen y el desarrollo de cualquier criatura se ven favorecidos por una lucha por la existencia lo suficientemente intensa como para exigir el ejercicio pleno y frecuente de sus facultades mentales y físicas, resulta interesante rastrear la secuencia de acontecimientos naturales que prepararon a Europa occidental para la llegada de las primeras ramas de la raza humana. Los bosques y las plantas reflejan aún más vívidamente que los animales las condiciones cambiantes del medio ambiente y la temperatura que marcaron la llegada y las diversas vicisitudes de la gran Edad de Hielo.
[ pág. 61 ]
Los bosques del centro de Francia en el Plioceno, así como los del valle del Arno en el norte de Italia, eran muy similares a los bosques actuales del centro de Estados Unidos, con árboles como el sasafrás, la acacia, la acacia de tres espinas, el zumaque, el ciprés calvo y el tulipán. Así, las regiones que albergaban la rica fauna forestal y de pradera del norte de Italia en el Plioceno Superior abundaban en árboles que hoy en día son familiares en Carolina del Norte y del Sur, incluyendo incluso especies típicamente americanas como el liquidámbar (Liquidambar styraciflua), el liquidámbar (Nyssa sylvatica) y el laurel, además de los ya mencionados. Al sur, a lo largo del Mediterráneo, también prosperaban árboles propios de un clima más tropical: el bambú, la palmera sabal y la palmera enana; lo más interesante es la presencia del sabal, que ahora prospera en las selvas subtropicales del centro de Florida. La secuoya también era abundante. Hacia finales del Plioceno, los primeros indicios de la llegada de la Época Glacial fueron un descenso de la temperatura y, quizás en las zonas montañosas más altas, el inicio de las etapas glaciales.
Los ancestros de los bosques modernos de Europa predominaron en el centro de Francia: el roble, el haya, el álamo, el sauce y el alerce. Estos bosques, que sobrevivieron a las vicisitudes de las épocas glaciales, dieron origen a los bosques de la Europa posglacial, mientras que todos los tipos puramente americanos desaparecieron de Europa y ahora solo se encuentran en las regiones templadas de Estados Unidos.
Hemos visto que se han descubierto pocos simios antropoides en el Plioceno Medio o Superior de Europa; la línea mono-gibón desaparece con los Pliohylobates del Plioceno Superior. Sin embargo, estos animales rara vez se encuentran en forma fósil, debido a su refugio en los árboles en épocas de inundaciones y peligro, por lo que no debemos asumir necesariamente que los antropoides se extinguieron en Francia. Los primates que se encuentran en el Plioceno Superior pertenecen a los tipos inferiores de los monos del Viejo Mundo, emparentados con el langur actual de la India y con el macaco y el babuino. La evidencia, hasta donde llega, indica que los ancestros del hombre estaban evolucionando en esa época en Asia y no en Europa. Esta evidencia, sin embargo, quedaría completamente desmentida si se pudiera demostrar que los eolitos, o pedernales primitivos, encontrados en varias partes de Europa desde el Oligoceno hasta el Pleistoceno son en realidad artefactos de origen humano o prehumano.
Español Los mamíferos de Europa en el Plioceno se derivaron de migraciones muy remotas desde América del Norte y, más directamente, desde el sur de Asia. El elemento oriental es muy fuerte, incluyendo tipos de rinocerontes ahora peculiares de Sumatra y el sur de Asia, numerosos mastodontes muy similares a los tipos del sur de Asia de la época, gacelas y antílopes, incluyendo tipos relacionados con los elands existentes, y tipos primitivos de caballos y tapires. Entre los carnívoros en Europa similares a las especies del sur de Asia estaban las hienas, los osos perro (Hyaenarctos), las civetas y los pandas (Ailurus); también estaban los tigres dientes de sable y numerosos otros felinos. En los árboles se encontraron los monos del sur de Asia y el norte de África; y en los bosques el ciervo axis, ahora restringido a Asia. Pero el animal afroasiático más distintivo de este período se encontró en los ríos; En concreto, el hipopótamo, que llegó a Italia a principios del Plioceno y se extendió hacia el sur, a través del puente terrestre siciliano, hasta el norte de África, y hacia el este, a lo largo de la costa sur del Mar Negro, hasta las colinas de Siwalik en la India. Así pues, muchos de los ancestros de lo que hemos denominado el grupo de mamíferos afroasiáticos del Pleistoceno ya habían llegado a Europa a principios del Plioceno. En el Plioceno medio y tardío llegaron tres tipos de mamíferos muy importantes que desempeñaron un papel fundamental en el Pleistoceno temprano. Estos son:
Los verdaderos caballos (Equus stenonis) de origen remoto norteamericano.
El primer ganado vacuno auténtico (Leptobos elatus), originario del sur de Asia.
Los verdaderos elefantes, primero Elephas planifrons y después E. meridionalis, más conocido como mamut del sur, ambos originarios de Asia.
[ pág. 63 ]
Los bosques y las riberas fluviales del valle del Arno, cerca de Florencia, albergaban a todos estos animales afroasiáticos en el Plioceno Superior. De aquí recibieron los nombres que nos recuerdan esta región de Italia tal como es hoy, como el rinoceronte etrusco (Dicerorhinus etruscus), el macaco florentino (Macacus florentinus), el caballo de Steno (Equus stenonis) y el ganado etrusco (Leptobos etruscus), que fue el primer buey en llegar a Europa.
En Italia y Francia, estos mamíferos afroasiáticos se mezclaron con ancestros del grupo euroasiático de bosques y praderas, más resistente. De estos, los más elegantes eran una variedad de ciervos con astas muy elaboradas o muy ramificadas, de ahí que se los conozca como ciervos «policladinos». En los bosques vagaban los jabalíes de Auvernia (Sus arvernensis), también los osos de Auvernia (Ursus arvernensis), los linces, los zorros y los gatos monteses. En los ríos nadaban la nutria y el castor, estrechamente emparentados con las formas preexistentes. Entre las rocas de las altas colinas se encontraban las picas o liebres sin cola (Lagomys), además de hámsteres, topos y musarañas.
Muchos de los animales más característicos de las mesetas secas modernas de África habían desaparecido de Europa antes del final del Plioceno, a saber, especies de gacelas, antílopes y caballos hipparion, todos ellos adaptados a las tierras altas y los desiertos de África. En la fauna plioceno reciente restante de autores franceses encontramos evidencia de que el Plioceno en toda Europa occidental cerró con un clima húmedo, cálido y templado, con extensos bosques y ríos intercalados con praderas favorables para la vida de una gran variedad de ciervos ramoneando, así como de elefantes, caballos y ganado vacuno. La flora del Plioceno Medio, tal como se encuentra en Meximieux, indica una temperatura media anual de 16° a 16° C.
Una de las pruebas de la disminución gradual de la temperatura hacia el final del Plioceno en Europa es el retroceso hacia el sur y la desaparición de los monos y simios; el gibón del Mioceno superior se encuentra tan al norte como Eppelsheim, cerca de Worms, Alemania; en el Plioceno inferior los monos y simios se encuentran sólo en los bosques del sur de Francia; en el Plioceno superior se registran sólo en los bosques del norte de Italia; la evidencia, hasta donde llega, indica un retroceso gradual hacia el sur.
Finalmente, al final del Plioceno, existían relaciones geográficas muy estrechas hacia el este con la vida mamífera de la India a través de lo que entonces era el istmo de los Dardanelos, y hacia el sur con la vida mamífera de África a través del puente terrestre siciliano. Esto indicaría que las largas rutas migratorias hacia el este y el oeste eran abiertas y propicias para la llegada a Europa occidental de nuevos migrantes procedentes del Lejano Oriente, incluyendo quizás a las razas humanas más primitivas. No existe la menor evidencia de que el hombre del Plioceno o sus ancestros existieran en Europa, salvo la que pueden proporcionar los problemáticos eolitos o los pedernales más primitivos.
¶ La primera glaciación
En el Plioceno Superior, corrientes marinas frías6 provenientes del norte comenzaron a fluir a lo largo de la costa sureste de Inglaterra, con indicios de un descenso gradual de la temperatura que culminó en una época en que el mar abundaba en moluscos árticos, que se han conservado en los «Weyboum Crags», una formación geológica a lo largo de la costa de Norfolk. Esta corriente ártica fue el precursor de la Primera Etapa Glacial.
No parece que se formara un casquete glaciar de extensión considerable en Gran Bretaña en esta etapa, pero por esta época se formó el primer gran casquete glaciar en la Norteamérica británica al oeste de la Bahía de Hudson, que extendió sus capas de hielo hasta Iowa y Nebraska. En este último estado, los bosques de píceas y otras coníferas indican la aparición de una flora templada fría antes de la glaciación. En los Alpes suizos, la nieve descendió 1200 metros por debajo de la línea de nieve actual, y en Escandinavia y el norte de Alemania se formaron las primeras grandes capas de hielo de las que fluyeron los glaciares y ríos que transportaron el «Viejo Diluvio» o la «deriva más antigua». Acompañando la ola de frío a lo largo de la costa oriental de Inglaterra, observamos, en los famosos depósitos fósiles conocidos como el «Lecho Forestal de Cromer», [ p. 65 ] que yacen sobre los riscos de Weybourn, la llegada del abeto (Abies) desde el norte. Esto es sumamente significativo, ya que hasta entonces solo se conocía en la región ártica de Crinnell Land, y esta fue su primera aparición en Europa central. Otro presagio de las condiciones septentrionales fue la primera aparición del buey almizclero en Inglaterra, que se atribuye7 a los depósitos del «Forest Bed».
Si bien Gran Bretaña se vio menos afectada en esta época que otras regiones, no cabe duda de la vasta extensión de la Primera Etapa Glacial en la América Británica, Escandinavia y los Alpes; en esta última región, Penck y Bruckner la han denominado «etapa Günz». Los depósitos de «deriva» tienen un espesor general de 30 m (98 pies), pero están en gran parte cubiertos y enterrados por los de la mucho más extensa Segunda Etapa Glacial. La capa de hielo escandinava8 no solo ocupó la cuenca del Báltico, sino que inundó Escania —la parte sur de Suecia— y se extendió hasta Hamburgo y Berlín. En los Alpes, los glaciares descendieron por todos los grandes valles montañosos hasta las tierras bajas del promontorio, lo que implica una depresión de la línea de nieve hasta 1200 metros por debajo de su nivel actual.

¶ La primera etapa interglaciar. Eolitos
Las pruebas de que una ola de frío prolongada atravesó Gran Bretaña durante la primera glaciación se ven en sus efectos posteriores, concretamente en la modernización de los bosques y en la desaparición, tanto en Gran Bretaña como en Francia, de una cantidad considerable de animales que abundaban en el Plioceno Superior. Sin embargo, la mayor parte de la vida de los mamíferos del Plioceno sobrevivió, lo que demuestra que, si bien las condiciones climáticas muy frías y las fuertes precipitaciones de humedad pudieron haber caracterizado las regiones que rodeaban los campos de hielo, el resto de Europa occidental, como mucho, atravesó un período frío prolongado durante el clímax de la Primera Etapa Glacial. A esto le siguió, durante el Primer Interglaciar, el regreso de un período algo más cálido que el actual.
Esta Primera Etapa Interglaciar se conoce como el Norfolkiano, debido a que se reconoció por primera vez en Europa en los depósitos conocidos como el «Lecho Forestal de Cromer», Norfolk, que contienen ricos registros no solo de los bosques de la época, sino también de las nobles formas de mamíferos que vagaban por Gran Bretaña y Francia en la época del Norfolkiano. Los bosques de Norfolk, en la latitud 52° 40 N, abundaban principalmente en árboles aún autóctonos de esta región, como el arce, el olmo, el abedul, el sauce, el aliso, el roble, el haya, el pino y la pícea, una flora forestal que se corresponde estrechamente con la de las costas de Norfolk y Suffolk de Inglaterra en la actualidad, aunque encontramos en esta flora fósil varias especies exóticas que le confieren un carácter ligeramente diferente.9 A partir de esta flora arbórea, Reid concluye que el clima del sureste de Inglaterra era casi el mismo que en la actualidad, pero ligeramente más cálido.
Notamos especialmente que se había producido un cambio muy grande en la desaparición total en estos bosques de los árboles que en el Plioceno eran comunes a Europa y América, como se ha descrito más arriba; en otras palabras, la flora de Europa se empobreció mucho durante la primera ola de frío.
En el sur de Francia, al igual que en la actualidad, las condiciones climáticas interglaciares fueron más suaves, pues encontramos numerosas especies de plantas, que ahora están representadas en el Cáucaso, Persia, el sur de Italia, Portugal y Japón. Así, la Primera Etapa Interglaciar, que fue relativamente corta, disfrutó de una temperatura que ahora corresponde a unos 4° de latitud más al sur.
Esta Primera Etapa Interglacial también se conoce como St.-Prestien, ya que entre las numerosas localidades de Francia e Italia que preservan la vida vegetal y mamífera de la época, la de St. Prest, en la cuenca parisina, es la más famosa. Aquí, en 1863, Desnoyers10 informó por primera vez del descubrimiento de varios huesos de mamíferos con líneas de incisión, que consideró obra del hombre. En aquel entonces, estos depósitos se consideraban de la era del Plioceno, lo que dio lugar inmediatamente a una teoría generalizada [ p. 68 ] sobre la aparición del hombre ya en el Plioceno. El origen humano de las incisiones descubiertas por Desnoyers ha sido objeto de controversia durante mucho tiempo y ahora se considera muy improbable. Líneas similares podrían ser de origen animal, concretamente, marcas dejadas por garras o dientes, o debidas a la presión accidental de superficies cortantes afiladas. Sin embargo, no pretendemos expresar una opinión válida sobre la causa de estas incisiones. La supuesta confirmación de la evidencia de Desnoyers sobre la existencia del hombre del Plioceno fue el supuesto hallazgo por Abbott de varios pedernales trabajados, dos in situ, en el lecho forestal de Cromer, Norfolk. Muchos años después, en depósitos similares de St. Prest, se descubrieron los supuestos eolitos, a los que Rutot se refiere como Étage Prestien. Por lo tanto, la edad de los depósitos de St. Prest es un asunto de sumo interés e importancia.

[ pág. 69 ]
St. Prest no es plioceno; es más bien el yacimiento pleistoceno más antiguo de la cuenca de París,11 y estos huesos de mamíferos incisos probablemente datan del Primer Interglaciar. El lecho que ha producido los huesos incisos y la rica serie de fósiles consiste en arenas y gravas fluviales gruesas, que forman parte de una terraza alta, a 30 m (98 pies) sobre el nivel actual del río Eure. Esta, al igual que otras terrazas altas, contiene una fauna característica del Primer Interglaciar, incluyendo el mamut meridional (E. meridionalis) y el caballo de Steno (E. stenonis). También encontramos aquí otros mamíferos muy característicos del Pleistoceno temprano, como el rinoceronte etrusco (D. etruscus), el hipopótamo gigante del Pleistoceno temprano (H. major), el castor gigante del Pleistoceno temprano (Trogontherium), tres formas del castor común (Castor), y una de bisonte (Bison antiquus). Esta vida mamífera de St. Prest es muy similar a la de Norfolk, Inglaterra; a la de Malbattu en Francia central, Puy-de-Dome; de Peyrolles, cerca de la desembocadura del Ródano, en el sur de Francia; de Solilhac cerca de Puy; de Durfort, Gard; de Cajarc, Lot-et-Garonne; y finalmente a la del valle del Arno, en el norte de Italia.
Una razón por la que ciertos autores, como Boule y Depéret, han situado esta etapa en el Plioceno Superior es que entre los mamíferos se encuentran numerosas formas supervivientes del Plioceno, como los tigres dientes de sable (Machaerodus), el ciervo policladino con sus elaboradas astas (C. sedgwicki), el rinoceronte etrusco y el primitivo caballo de Steno. Sin embargo, recientemente hemos descubierto que, con la excepción del ciervo policladino, estos mamíferos ciertamente sobrevivieron en Europa hasta la Segunda Etapa Interglaciar, y se dice que existen evidencias de que algunos incluso persistieron hasta la Tercera Etapa Interglaciar.
Por lo tanto, es la extinción o desaparición en Europa de muchos de los animales, muy abundantes incluso a finales del Plioceno, lo que marca esta fauna como del Pleistoceno temprano. Ya no se encuentran simios antropoides; de hecho, no hay evidencia de la supervivencia de ningún primate, excepto los macacos, que sobreviven en los Pirineos hasta finales del Pleistoceno; el tapir ha desaparecido por completo de los bosques europeos; pero la desaparición más significativa es la del mastodonte, que se cree que permaneció en el norte de África y que, sin duda, sobrevivió en América hasta muy finales del Pleistoceno. La fauna de Europa occidental, al igual que la flora, ha perdido parte de su aspecto pliocénico, mientras que conserva otra, tanto en su fauna mamífera como en su flora forestal.
Además, el entorno vital en su conjunto adquiere un aspecto novedoso con la llegada, principalmente desde el norte, de animales y plantas más resistentes que habían estado evolucionando durante un largo período en los bosques templados y praderas de Eurasia, al noreste y noroeste. De esta región euroasiática provino el ciervo rojo (Cervus elaphus), el ciervo gigante (Megaceros), y de los pantanos septentrionales, el alce cabeciblanco (Alces latifrons). La presencia de miembros de la familia de los cérvidos (Cervidae) en gran número y con diversas líneas de descendencia es uno de los rasgos más distintivos del Primer Interglaciar. Además de las nuevas formas septentrionales mencionadas, existía el corzo (Capreolus), que aún sobrevive en Europa, pero del que ya no existen registros. [ p. 71 ] El hermoso ciervo axis (Axis), que ahora se ha retirado al sur de Asia. El ciervo policladino, observado por primera vez en el valle del Arno, está representado en el Primer Interglaciar por el ciervo de Sedgwick (C. sedgwicki) en Norfolk, y por la especie C. dicranius del norte de Italia, donde también se encuentra el ciervo de los Carnutes (C. carnutorum).

Observamos que predominan los tipos de animales que ramonean, viven en el bosque y en los ríos. Entre los carnívoros que frecuentan el bosque se encuentran el glotón, la nutria, dos tipos de oso: el lobo, el zorro y la marta; otro habitante del bosque es el jabalí, emparentado con el Sus scrofa, también existente en Europa.
Así, a principios del Pleistoceno, los bosques europeos estaban repletos de una vida salvaje muy similar a la de la prehistoria, a la que se unía el elemento oriental: los grandes elefantes, rinocerontes e hipopótamos que conectaban Europa con el Lejano Oriente. Entre estos migrantes orientales del Pleistoceno temprano se encontraban dos recién llegados: el ganado salvaje primitivo (Bos primigenius) y el primer bisonte (Bison priscus).
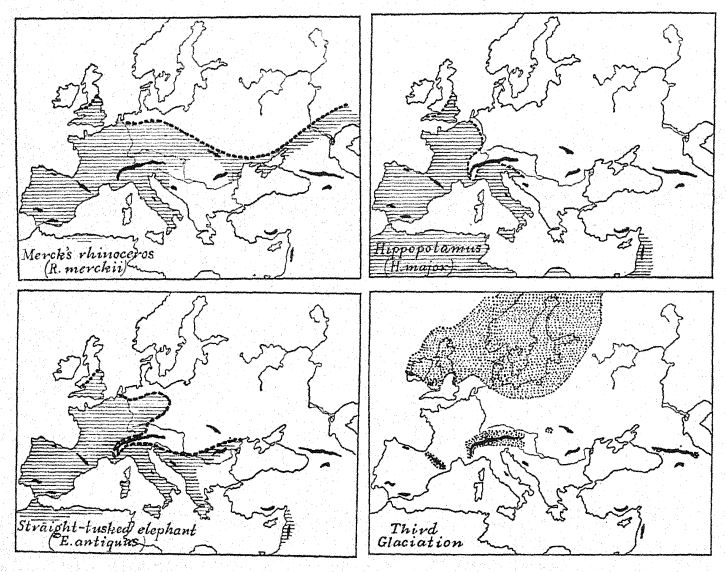
El mapa teórico de Europa occidental durante el Primer Interglaciar (Fig. 12, también Fig. 56) nos permite comprender estas migraciones desde el noreste y desde Oriente. Como lo indican los canales fluviales hundidos descubiertos en la antigua plataforma continental, la línea costera se extendía mucho hacia el oeste hasta los límites de la meseta continental que ahora está hundida profundamente bajo el océano; las Islas Británicas estaban separadas de Francia no por el mar sino por un amplio valle, mientras que el Rin, con el Támesis como afluente occidental, fluía hacia el norte sobre una extensa llanura aluvial, que es el lecho actual de la cuenca del Mar del Norte. No es improbable que los ricos depósitos de vida mamífera en el ‘Forest Bed of Cromer’, Norfolk, fueran arrastrados por afluentes de este antiguo río Rin.
En todos los grandes ríos de esta ampliada Europa occidental habitaban los hipopótamos, y a lo largo de las riberas y en los bosques ramoneaban los rinocerontes etruscos. Entre las especies de ganado vacuno salvaje (Bos, Leptohos) que pastaban en las praderas de la región de Norfolk, en Gran Bretaña, se encontraban especies de ganado vacuno salvaje (Bos, Leptohos), junto con dos especies de caballos, incluyendo una forma más ligera parecida al caballo de Steno (E. stenonis cocchi) del Valle del Arno y una más pesada, probablemente perteneciente a los bosques. El elefante gigante de este período es el mamut meridional (E. meridionalis trogontherii), un descendiente algo especializado del mamut meridional del Plioceno del valle del Arno; este animal es más conocido por un magnífico ejemplar descubierto en Durfort (Fig. 42) y conservado en el Museo de París. Se dice que alcanzó una altura de más de 3,6 metros, en comparación con los 3,4 metros, la altura de los elefantes africanos más grandes que existen. Es probable que todos estos migrantes del sur de Asia que llegaron a Europa estuvieran parcial o totalmente cubiertos de pelo, como adaptación al clima cálido y templado de veranos e inviernos fríos. Al sur, en el clima aún más suave de Italia, se registra la llegada de otra gran especie, conocida como el elefante «antiguo» o «de colmillos rectos» (E. antiquus). Este animal aún no había llegado a Francia ni a Gran Bretaña.
Los grandes machasrodontos, o tigres dientes de sable, se alimentaban de los indefensos miembros de esta heterogénea fauna y se extendían por Europa, el norte de África y Asia. No parece que los verdaderos leones (Felis leo) hubieran entrado aún en Europa.
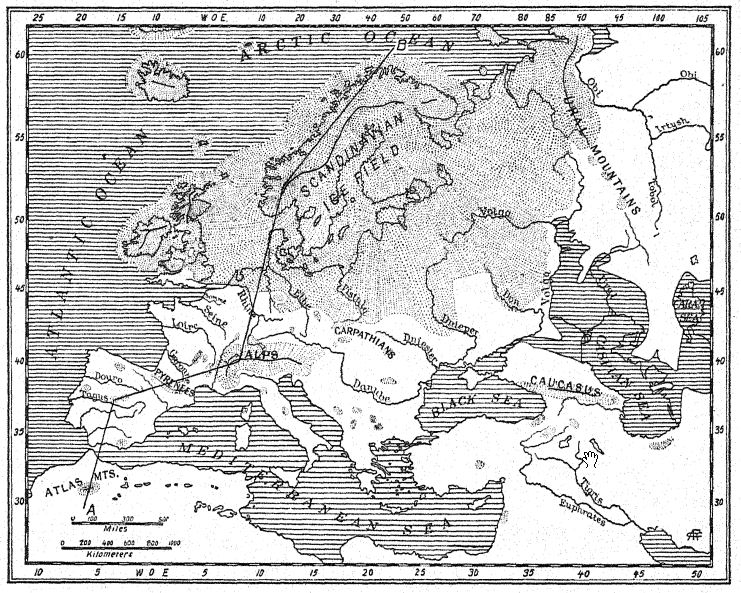
La intercomunicación de la vida en una vasta área que se extiende 9.600 kilómetros desde el valle del Támesis, al oeste, hasta la India, al sureste, se evidencia por la presencia de seis o más especies similares o relacionadas de elefantes y rinocerontes. A dos mil quinientos kilómetros al sureste de las estribaciones del Himalaya, manadas similares de mamíferos, pero en una etapa anterior de su evolución, vagaban por la isla de Java, que entonces formaba parte del continente asiático.
¶ La raza Trinil de Java
El interés humano en esta gran multitud de seres vivos reside en que las rutas migratorias abiertas por estas grandes razas animales también pudieron haber proporcionado un camino para las primeras razas humanas. Así, el descubrimiento de la raza Trinil en Java central, en medio de una fauna estrechamente relacionada con la de las estribaciones del Himalaya y más remotamente relacionada con la del sur de Europa, tiene una relación más directa con nuestro tema de lo que a primera vista parece.
En el río Bengawan, en Java central, Eugen Dubois, cirujano del ejército neerlandés, había estado excavando en busca de fósiles con la esperanza de encontrar restos prehumanos. En 1891, encontró cerca de Trinil un yacimiento de numerosos huesos de mamíferos, incluyendo un molar superior que consideró perteneciente a una nueva especie de simio. Al retirar cuidadosamente la roca, apareció la parte superior de un cráneo a aproximadamente un metro del diente. Una excavación posterior al final de la temporada de lluvias reveló un segundo molar y un fémur izquierdo a unos 15 metros del lugar donde se encontró el cráneo, incrustados y fosilizados de la misma manera. Estas partes dispersas fueron descritas por Dubois13 en 1894 como el tipo de Pithecanthropus erectus[2], término que designa al [ p. 74 ] hombre-mono erguido. El término específico erectus se refiere al fémur, del cual el autor observa: "Por lo tanto, debemos concluir que el fémur de Pithecanthropus fue diseñado para las mismas funciones mecánicas que el del hombre. Las dos articulaciones y el eje mecánico corresponden tan exactamente a las mismas partes en el hombre que la ley de perfecta armonía entre la forma y la función de un hueso requerirá la conclusión de que esta criatura fósil tenía la misma postura erguida que el hombre y también caminaba sobre dos piernas. … De esto se sigue necesariamente que la criatura tenía el libre uso de las extremidades superiores, ahora superfluas para caminar, y que estas últimas sin duda ya estaban muy avanzadas en esa línea de diferenciación que las desarrolló en la humanidad en herramientas y órganos del tacto. . . . De un estudio del fémur y el cráneo se deduce con certeza que este fósil no puede clasificarse como simio. . . . Y, como con el cráneo, también con el fémur, las diferencias que separan a Pithecanthropus del hombre son menores que las que lo distinguen del antropoide más alto. … Aunque muy avanzada en su proceso de diferenciación, esta forma del Pleistoceno aún no había alcanzado el tipo humano. El Pithecanthropus erectus es la forma de transición entre el hombre y los antropoides que, según las leyes de la evolución, debieron existir. Es el ancestro del hombre.


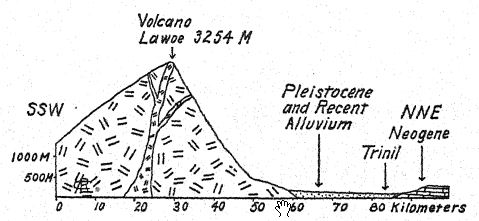
De este modo, el autor situó a Pithecanthropus en una nueva familia, del orden Primates, a la que llamó Pithecanthropidae.
La edad geológica de los huesos mencionados es de suma importancia. Los restos de Pithecanthropus se encontraban en un depósito de aproximadamente un metro de espesor, compuesto por areniscas tobáceas sueltas y gruesas; debajo, un estrato de arcilla dura de color gris azulado; y debajo, una brecha marina. Por encima de la capa de Pithecanthropus se encontraban los estratos «Kendeng», una arenisca tobácea multiestratificada de unos 15 metros de espesor. Dubois y otros consideraron que esta serie geológica era del Terciario tardío o del Plioceno; por consiguiente, Pithecanthropus pasó a ser conocido como el tan esperado «hombre-mono del Plioceno». Investigaciones posteriores realizadas por geólogos expertos han tendido a atribuir la edad al Pleistoceno temprano.17 Según Elbert18, los estratos de Kendeng que recubren la capa de Pithecanthropus corresponden a un período pluvial temprano de bajas temperaturas y, en términos temporales, a la [ p. 76 ] Edad de Hielo de Europa. Incluso en Java se pueden distinguir tres divisiones del Pleistoceno, incluyendo el primer período de bajas temperaturas al que se refiere la capa de Pithecanthropus.
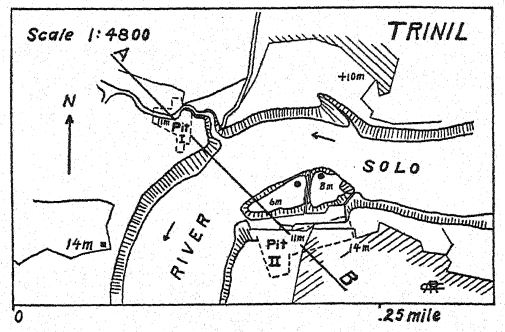
Los mamíferos fósiles contenidos en la capa Pithecanthropus también han sido estudiados a fondo,19 y tienden a confirmar la referencia original al Plioceno superior. Proporcionan una fauna muy rica similar a la de las colinas Siwalik de la India, que incluye el puercoespín, el pangolín, varios felinos, la hiena y la nutria. Entre los primates además de Pithecanthropus hay un macaco. Entre los ungulados más grandes hay dos especies de rinoceronte relacionadas con las formas indias existentes, el tapir, el jabalí, el hipopótamo, el ciervo axis y el ciervo rusa, el búfalo indio y el ganado salvaje. Cabe destacar que aparecen tres especies de elefantes del Plioceno tardío, todas conocidas como Stegodon, y especialmente la especie Stegodon ganeza, así como Elephas hysudricus, una especie relacionada con E. antiquus, o el elefante de colmillos rectos, que entró en Europa a principios del Pleistoceno. Se han encontrado fósiles de los mismos animales en las estribaciones del Himalaya de la India, a unos 4000 kilómetros al noroeste. Los depósitos de la India se consideran del Plioceno Superior,20, ya que este es el período final de la vida de los Siwaliks superiores de la India.
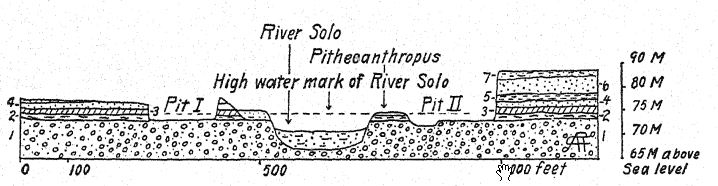
[ pág. 77 ]

Ciertamente, Java formaba parte del continente asiático, y manadas similares de grandes mamíferos vagaban libremente por las llanuras desde las faldas del Himalaya hasta las orillas del antiguo río Trinil, mientras que simios similares habitaban los bosques. En esa época, el orangután pudo haber penetrado en los bosques de Borneo, que actualmente constituyen su hogar; es el único simio encontrado hasta la fecha en el Plioceno superior de la India. Por lo tanto, podemos anticipar el descubrimiento, en cualquier momento, en la India de una raza similar al Pithecanthropus.
La edad geológica de la raza Trinil debe considerarse, por tanto, como Plioceno tardío o Pleistoceno temprano.
Este gran descubrimiento de Dubois suscitó una discusión extensa y acalorada, en la que participaron los anatomistas y paleontólogos más destacados del mundo. Algunos consideraban el cráneo como el de un gibón gigante, otros como prehumano y otros como una forma de transición. Sin embargo, podemos formarnos nuestra propia opinión a partir de una comprensión más completa de los propios especímenes, teniendo siempre presente que es una cuestión de si el fémur y el cráneo pertenecen al mismo individuo o incluso a la misma raza. En primer lugar, nos sorprende el marcado parecido que guarda la parte superior del cráneo, tanto al observarlo de lado como desde arriba, con el de la raza neandertal. Esto justifica plenamente la opinión del anatomista Schwalbe21 de que el cráneo del Pithecanthropus se acerca más al del hombre de Neandertal que al del [p. 78 ] El más alto de los simios antropoides. Según las mediciones de Schwalbe, el índice de la altura del cráneo (Kalottenhöheindex) puede compararse con otros de la siguiente manera:
| La raza humana más baja | 52 por ciento. |
| Hombre de Neandertal | 40,4 por ciento. |
| Pithecanthropus, o raza Trinil | 34,2 por ciento. |
Esto concuerda con la estimación de la capacidad cerebral[3] de 855 c. cm. (Dubois) en comparación con 1230 c. cm., la menor capacidad cerebral encontrada en un miembro de la raza neandertal. En segundo lugar, como se ve desde arriba, nos sorprende la gran longitud del cráneo en comparación con su anchura, siendo el índice cefálico o proporción de anchura a longitud del 73,4 por ciento (Schwalbe) en comparación con el 73,9 por ciento en el cráneo de tipo neandertal; esta dolicocefalia concuerda con el hecho de que todas las razas humanas más tempranas encontradas hasta ahora tienen cabezas alargadas, aunque según Schwalbe22 todos los antropoides tienen cabezas anchas. Esta es una distinción muy importante. La tercera característica es la prominencia y anchura de las crestas óseas de las cejas sobre las órbitas, que son casi tan grandes como en el chimpancé y superan con creces las [ p. 79 ] de la raza neandertal y del australiano moderno. El perfil de la cabeza de Trinil restaurado por McGregor (Fig. 38) exhibe esta prominente cresta ósea y la frente baja y retraída. Según la opinión más reciente de Schwalbe23, el Pithecanthropus puede considerarse uno de los ancestros directos del hombre de Neandertal e incluso de la especie humana superior, el Homo sapiens. También considera que cuando se conozca la mandíbula inferior de la raza Trinil, se descubrirá que es muy similar a la del hombre de Heidelberg, concluyendo que el Pithecanthropus y el casi emparentado hombre de Heidelberg pueden considerarse ancestros comunes de la raza neandertal, por un lado, y de las razas superiores, por otro. Sin embargo, existen razones para excluir al Pithecanthropus de la línea ancestral directa de las razas humanas superiores.

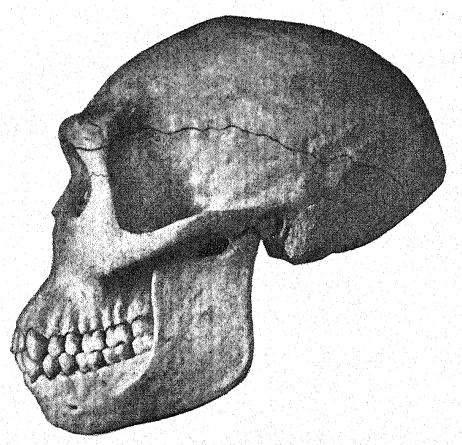
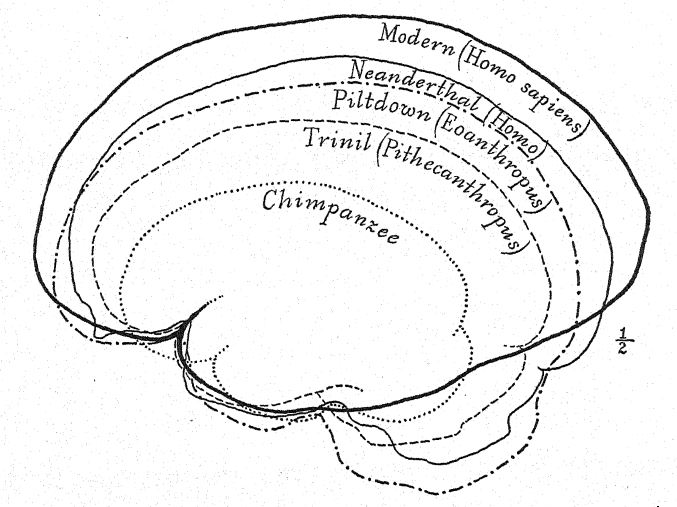
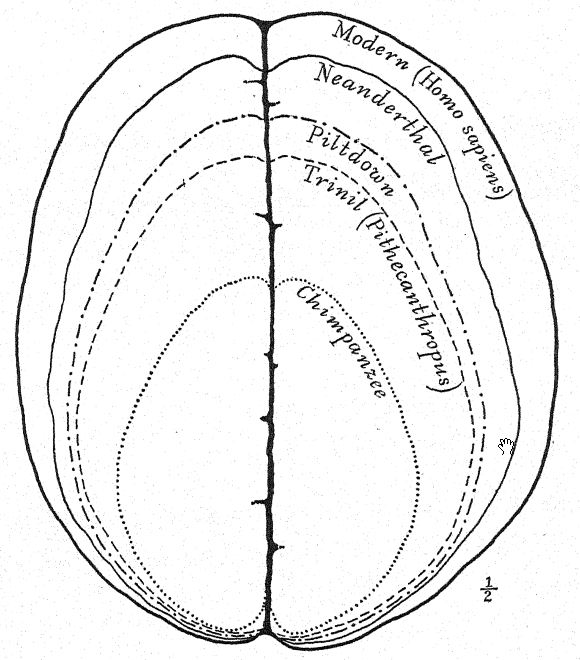
Esta etapa prehumana tiene, sin embargo, una gran importancia en la historia del desarrollo del hombre. En nuestra opinión, es precisamente la etapa que, teóricamente, deberíamos anticipar encontrar en los albores del Pleistoceno. Buchner adopta una perspectiva similar, presentando en un admirable diagrama (Fig. 117) el resultado de su comparación de doce caracteres diferentes en los cráneos de Pithecanthropus, los neandertales, los australianos y los tasmanos. Uno de los principales objetivos de la investigación de Buchner fue una comparación muy detallada del cráneo de Trinil con el de la humilde y ahora extinta raza tasmana, que, como observamos en el diagrama, ocupa una posición apenas superior a la de la raza Spy-Neandertal.
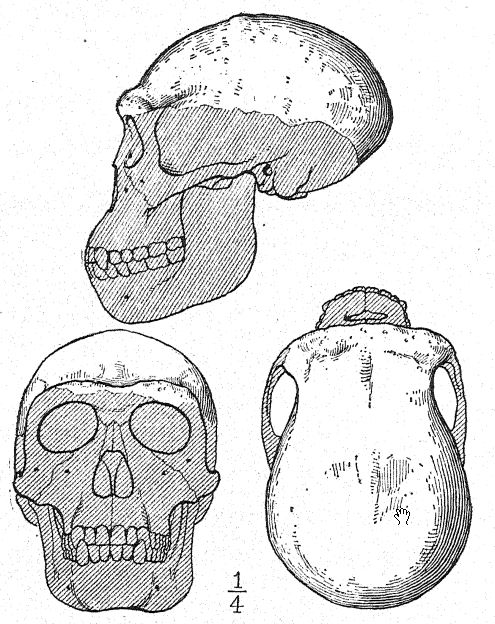
Si el fémur se corresponde con el cráneo, los Trinils eran una raza alta, alcanzando una altura de 1,70 m, en comparación con los 1,60 m de los neandertales. El fémur (Fig. 122) presenta una curvatura muy leve en comparación con la de cualquier simio o lémur, y en este aspecto es más humano; es notablemente alargado (455 mm), superando al de los neandertales; la tibia probablemente era correspondientemente corta. Los dos dientes superiores de molienda conservados son mucho más humanos que los del gibón, pero no se parecen lo suficiente a los del hombre como para confirmar la teoría prehumana. Dubois observa:25 «Que el diente pertenezca a alguna forma homínida no requiere mayor demostración. Aparte de su tamaño y la mayor rugosidad de la superficie de molienda, se diferencia del molinillo humano en que la cúspide menos desarrollada del Pithecanthropus es la cúspide posterior junto a la mejilla, mientras que en el hombre generalmente es la cúspide posterior junto a la lengua. La simplificación de la corona y la raíz del molinillo Trinil es tan extensa como suele serlo en el hombre».
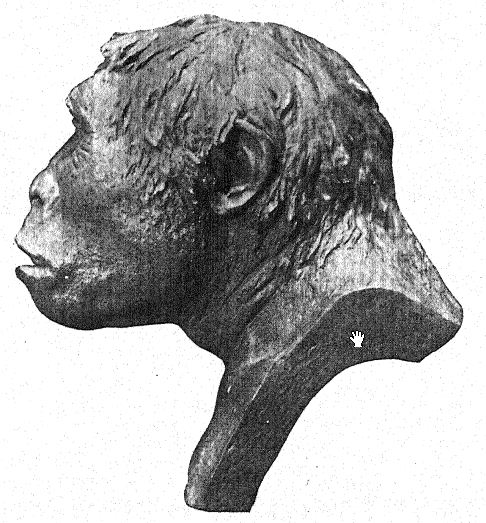
Se han realizado diversos esfuerzos para complementar los escasos y dispersos materiales recopilados por Dubois. La expedición Selenka de 1907-1908 trajo consigo un molar inferior izquierdo humano como único resultado de una búsqueda exprés de más restos de Pithecanthropus. [ p. 82 ] También se dice que Dubois posee el fragmento de una mandíbula inferior de aspecto primitivo procedente de la cordillera conocida como las colinas de Kendeng, en cuya base sur se encuentra el pueblo de Trinil.
Nos queda considerar el estadio de evolución psíquica alcanzado por la raza Trinil, y esto naturalmente depende de la actitud erguida y de lo poco que se sabe del tamaño y las proporciones del cerebro.
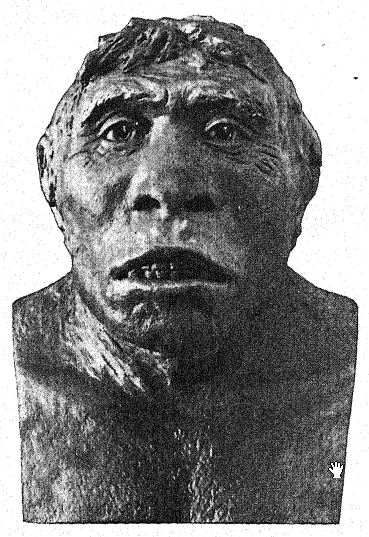
Adoptar la postura erguida no se trata simplemente de aprender a equilibrar el cuerpo sobre las extremidades posteriores.26 Implica cambios en el interior del cuerpo, la pérdida de la cola, la liberación de los brazos y el establecimiento del diafragma como el músculo principal de la respiración. El fémur del Pithecanthropus se asemeja tanto al del hombre que apoya la teoría de que la postura erguida pudo haber sido adoptada por los ancestros del hombre ya en el Oligoceno. Parecería que el Pithecanthropus tenía libre uso de los brazos y es posible que el control del pulgar y los dedos se hubiera cultivado, quizás en la fabricación de herramientas primitivas de madera y piedra.
El descubrimiento del uso de la madera como herramienta y arma probablemente precedió al del uso de la piedra.
Elliot Smith describe esta etapa de desarrollo de la siguiente manera:27 «… La emancipación de las manos del progreso arrojó toda la responsabilidad sobre las piernas, que se volvieron más eficientes para su función de soporte una vez que perdieron sus poderes prensiles y se alargaron y especializaron para una rápida progresión. Así, la postura erguida se estereotipó y fijó, y las extremidades se especializaron, y estos simios erguidos emergieron de sus bosques ancestrales en sociedades, armados con palos y piedras y con los rudimentos de todos los poderes que finalmente les permitieron conquistar el mundo. La mayor exposición al peligro que estos espíritus más aventureros encontraron una vez que emergieron al aire libre, y las constantes luchas que estas primeras criaturas semihumanas debieron haber tenido en encuentros con enemigos definidos, no menos que con las fuerzas de la naturaleza, proporcionaron los factores que rápidamente eliminaron a aquellos no aptos para las nuevas condiciones y, por selección natural, convirtieron a los supervivientes en verdaderos hombres».
La frente poco desarrollada del Pithecanthropus y la diminuta área frontal del cerebro indican que la raza Trinil tenía una capacidad limitada para aprovechar la experiencia y la tradición acumulada, pues en esta área prefrontal del cerebro se localizan las facultades de atención y control de las actividades de todas las demás partes del cerebro. En el cerebro del simio predominan las áreas sensoriales del tacto, el gusto y la visión, y estas están bien desarrolladas en el Pithecanthropus. El área central del cerebro, que es el almacén de los recuerdos de las acciones y de los sentimientos asociados a ellas, también está bien desarrollada, pero el área prefrontal, que es la sede de la facultad de sacar provecho de la experiencia o de recordar las consecuencias de respuestas anteriores a la experiencia, está desarrollada en un grado muy limitado.28 Así, mientras que el cerebro de Pithecanthropus se estima en 855-900 c. cm., en comparación con los 600 c. cm. del cerebro de simio más grande y los 930 c. cm. del cerebro más pequeño registrado en la raza humana, indica una etapa muy baja de inteligencia.
¶ Ausencia de Paleolitos y presencia de Eolitos en Europa Occidental
Volviendo a las condiciones del Primer Interglaciar en Europa, observamos que los cauces fluviales discurrían por los mismos valles que en la actualidad, pero que en las primeras glaciaciones los cauces eran mucho más anchos y se elevaban desde 100 hasta 45 metros por encima de los niveles actuales, relativamente estrechos, de los ríos. Las vastas inundaciones de la glaciación posterior llenaron estos valles, pero algunas de las terrazas altas ya se habían formado. Es fundamental destacar que nunca se han encontrado pedernales prechelleanos ni paleolitos auténticos en las arenas o gravas de estas terrazas altas.
Los eolitos hallados en este nivel de “terraza alta” en St. Prest pertenecen a la cultura Prestian de Rutot,29 que considera esta estación como del Plioceno Superior. Estos, al igual que otros supuestos pedernales eolíticos, son muy toscos, pero, a pesar de su rudeza, generalmente presentan una parte con la forma adecuada para ser agarrada con la mano, mientras que la otra parte es afilada o puntiaguda para cortar. Se admite generalmente que estos pedernales son en su mayoría de formas accidentales, y ha habido poca o ninguna prueba de que hayan sido moldeados por manos humanas. Sobre este punto, Boule30 observa: “En cuanto a los eolitos, he combatido la teoría no solo porque me parece improbable, sino porque una larga experiencia geológica me ha demostrado que a menudo es imposible distinguir las piedras partidas, cortadas o retocadas por agentes puramente físicos de ciertos productos de artesanía rudimentaria”.
Por otro lado, es interesante en este punto citar las palabras de MacCurdy:31 “Mi opinión, basada en mi experiencia personal, … es que se ha establecido la existencia de una industria primitiva, anterior a lo que comúnmente se acepta como Paleolítico. Esta industria se remonta al Mioceno Superior y continúa durante el Terciario Superior hasta el Cuaternario Inferior, inclusive. Las características distintivas de la industria se mantienen prácticamente sin cambios a lo largo de todo el período, ya que la subdivisión del período en épocas se basa en la estratigrafía [etapas geológicas] y no en características industriales. Como los requisitos en cuanto a herramientas eran muy simples y el suministro de material en forma de lascas naturales y fragmentos de sílex era muy abundante, la capacidad inventiva de la población permaneció latente durante siglos. El martillo y el cuchillo fueron las herramientas originales. Ambos se adquirían ya hechos. Una lasca natural de borde afilado servía para una, y un nódulo o fragmento para la otra. Cuando el filo de la lasca se desgastaba por el uso, la pieza se desechaba o se retocaba para su posterior uso. Si el martillo o la lasca no se sujetaban cómodamente en la mano, las puntas o filos problemáticos se eliminaban o reducían mediante cincelado. El inventario de herramientas aumentaba lentamente con el lento crecimiento de las necesidades. A medida que estas se multiplicaban y el suministro natural de materia prima disminuía, este último se complementaba con la fabricación de lascas artificiales. Cuando se aprendió la lección de asociar formas definidas de herramientas con usos específicos, surgieron tipos especiales, en particular el utensilio amigdaloide y el puñal. Luego vino la transición del Eolítico al Paleolítico, una etapa que Rutot ha investigado a fondo.
No es improbable que la raza Trinil estuviera en una etapa de cultura Eolítica; es muy probable que las razas prehumanas de esta era geológica muy remota utilizaran más de un arma de madera y piedra.
¶ La Gran Segunda Glaciación (Fig. 25, pág. 65)
A principios del Pleistoceno, una elevación general del sur de Europa unió las islas del Mediterráneo con Europa al norte y con África al sur, creando amplias conexiones terrestres entre ambos continentes que proporcionaron rutas migratorias tanto hacia el norte como hacia el sur. En esa época, ciertos mamíferos típicamente africanos, como el elefante de colmillos rectos y el león, probablemente se dirigían hacia el norte; Sicilia, por entonces, obtuvo su numerosa fauna de elefantes e hipopótamos, y la isla de Malta se conectó con el continente, así como con las islas orientales de Chipre y Creta. Parece probable que la conexión entre la península itálica y Malta se renovara más de una vez.
La aproximación de la segunda glaciación está indicada a lo largo de la costa sureste de Gran Bretaña por el hundimiento de la tierra y el ascenso del mar, acompañados por una nueva corriente ártica, que trajo consigo una invasión de moluscos árticos que se depositaron en una capa de lechos marinos directamente sobre los que contienen [ p. 87 ] [ p. 88 ] [ p. 89 ] rica y cálida fauna y flora del ‘Forest Bed of Cromer’, Norfolk.®^ También parece probable que una corriente fría del norte barriera las costas occidentales de Europa, y Geikie estima que se produjo una disminución de la temperatura de no menos de 20° Fahr., un cambio tan grande como el que se experimenta ahora al pasar del sur de Inglaterra al Cabo Norte.

La segunda glaciación fue, con mucho, la mayor, tanto en Europa como en América. En la región de los Pirineos, que en el período mucho más tardío de la Tercera Etapa Interglacial se convirtió en un territorio predilecto para el hombre paleolítico, existían glaciares de gran extensión. Esto se aprecia al compararlo con las condiciones actuales. El mayor de los glaciares actuales de los Pirineos tiene solo 3 kilómetros de longitud y termina a una altura de 2180 metros sobre el nivel del mar. Durante la mayor glaciación, la nieve parece haber descendido 1290 metros por debajo de su nivel actual. Desde los Pirineos, a través del valle del Gállego, hasta España, fluía un glaciar de 61 kilómetros de longitud, mientras que al norte, el glaciar del valle del Garona se extendía a lo largo de 72 kilómetros hasta un punto cercano a Montréjeau. Incluso en sus tramos más bajos, este glaciar tenía más de 800 metros de espesor. Al este se extendía un glaciar de 61 kilómetros de longitud, que llenaba el valle del Ariège y cubría los yacimientos de grandes cavernas paleolíticas como la de Niaux; es probable que en esta época comenzara la formación de esta caverna. Que estos glaciares fueron anteriores al período de la cultura achelense del Paleolítico Inferior lo demuestra el hecho de que con frecuencia se encuentran herramientas achelenses en la superficie de las morrenas formadas por estos antiguos témpanos de hielo.
Al norte se extendía el vasto campo de hielo escandinavo, que se extendía por Gran Bretaña y más allá de los valles del Rin, el Elba y el Vístula, llegando casi hasta los Cárpatos. Incluso las cadenas montañosas menores estaban cubiertas de glaciares, incluyendo la cordillera del Atlas en el norte de África.
En América del Norte, desde el gran centro al oeste de la bahía de Hudson, la capa de hielo extendió su deriva hacia el sur hasta Missouri, Iowa, Kansas y Nebraska, más allá de los límites de glaciaciones anteriores y posteriores.
[ pág. 90 ]
Los materiales de las principales «terrazas altas» de los grandes valles fluviales de Europa occidental se depositaron en esta época.
¶ Vida de la segunda etapa interglacial cálida
El largo período cálido que siguió a la Gran Glaciación es notable por presentar las primeras pruebas de la presencia del hombre en Europa occidental. Se trata del período de la raza humana de Heidelberg (Homo heidelbergensis), conocida únicamente a partir de una única mandíbula descubierta por Schoetensack en las arenas de Mauer, cerca de Heidelberg, en 1907. No se han encontrado otras pruebas de la existencia del hombre en ninguno de los depósitos que se formaron durante este vasto intervalo de tiempo geológico, a menos que aceptemos la teoría de Penck y de Geikie de que las canteras pre-chellianas y chellianas del río Somme pertenecen a la Segunda Etapa Interglacial.
La vasta duración de este período interglaciar se evidencia tanto en Europa como en América por el profundo corte y desgaste de las corrientes de aire arrastradas por la segunda glaciación. Penck cree que esta larga etapa cálida representa un período mayor que el intervalo completo entre la tercera glaciación y la actualidad. El clima inmediatamente después del retroceso de los glaciares era fresco y húmedo en las regiones glaciares, pero a esto le siguió un período tan prolongado de calor y sequedad que los glaciares de los Alpes se retrajeron hasta un punto muy por encima de sus límites actuales.
En una de las antiguas terrazas altas del río Inn, en el norte del Tirol, se encuentra un depósito que contiene la flora forestal predominante de la época, del cual Penck concluye que la temperatura en Innsbruck era 2 °C superior a la actual. En correspondencia con esto, la línea de nieve se situaba 300 metros por encima de su nivel actual, y los Alpes, salvo los picos más altos, estaban casi completamente desprovistos de hielo y nieve. Una planta característica es el rosal alpino póntico (Rhododendron ponticum), que florece actualmente a una temperatura media de 14-18 °C, lo que indica que el clima de Innsbruck era tan benigno como el de las laderas italianas de los Alpes actuales. Este rododendro se encuentra actualmente en el Cáucaso. Otras especies meridionales de la época eran el espino cerval, [p. 91 ] emparentada con una especie que ahora habita en las Islas Canarias, y el boj. También había plantas más resistentes, como el abeto (Pinus sylvestris), la pícea, el arce, el sauce, el tejo, el olmo, el haya y el serbal. Los bosques de la misma época en Provenza eran, en su mayor parte, similares a los que se encuentran actualmente en esa región; de treinta y siete especies, veintinueve aún se dan en esta parte del sur de Francia. En general, el aspecto del sur de Francia en esa época era sorprendentemente moderno. Los bosques incluían robles, olmos, álamos, sauces, tilos, arces, zumaques, cornejos y espinos. Entre las plantas trepadoras se encontraban la vid y la clemátide. También se encontraron algunas especies que desde entonces se han replegado al sur, como especies de laurel dulce y laurel, que ahora se encuentran confinadas en las Islas Canarias. La alta humedad de la época se evidencia por la presencia de ciertas especies de coníferas que requieren mucha humedad. Al igual que en el Primer Interglaciar, la presencia de higueras indica inviernos suaves.
Resulta difícil imaginar bosques de estas características modernas, que más al norte incluían especies aún más templadas y resistentes, como el escenario de la gran vida africana y asiática que se extendía por toda Europa occidental en aquella época. Fue la presencia de hipopótamos, elefantes y rinocerontes lo que dio a Lyell, Evans y otros primeros observadores la impresión de que una temperatura y vegetación tropicales eran características de este largo período de vida. Estos animales se consideraban anteriormente prueba de un clima casi tropical, pero la evidencia más fiable de los bosques, reforzada por la presencia de numerosos tipos resistentes de animales forestales y de pradera, ha descartado todas las teorías iniciales sobre temperaturas extremadamente cálidas durante el Segundo Interglaciar.
Los restos de lo que todavía se conoce como «fauna cálida» se encuentran principalmente en las arenas y gravas de los antiguos lechos del Neckar, el Garona y el Támesis, así como de otros ríos del norte y del sur, también en Essex, Inglaterra. Lo más sorprendente es que la vida de los mamíferos en Europa occidental permaneció completamente inalterada durante la vasta segunda glaciación que acabamos de describir; las pocas extinciones que se produjeron, así como la cantidad de recién llegados, pueden atribuirse a las nuevas conexiones geográficas con África por el sur y al constante avance de la migración desde el lejano oriente.
Hubo cuatro arribos muy importantes y distintivos provenientes del mundo afroasiático: el elefante de colmillos rectos o antiguo (E. antiquus), el rinoceronte de hocico ancho (D. merckii), el león africano (Felis leo) y el hicena africana (H. striata), que revelan estrechos vínculos geográficos con el norte de África. De estos, el elefante antiguo y el rinoceronte de hocico ancho eran compañeros cercanos; compartían las mismas regiones y las mismas temperaturas, sus restos se encuentran juntos con mucha frecuencia y sobrevivieron hasta el final de la gran etapa vital de Europa occidental, que concluyó con la llegada de la cuarta glaciación. Contrastan con la otra pareja de grandes mamíferos que ya estaba presente en Europa en el Plioceno y el Primer Interglaciar, es decir, el mamut austral, en esta etapa conocido como trogonte, que tenía preferencia por la compañía del hipopótamo (H. major); parecería que estos animales eran menos resistentes porque ambos [ p. 94 ] desaparecieron de Europa un poco antes que el antiguo elefante y el rinoceronte de Merck.
El león africano parece haber sido un competidor del tigre dientes de sable, ya que este último animal ahora es menos abundante, aunque hay razones para creer que sobrevivió hasta la Tercera Etapa Interglaciar. Junto con el antiguo tipo plioceno del tigre dientes de sable se encontraron también el rinoceronte etrusco, el oso primitivo de Auvernia (Ursus arvernensis) y el oso gigante (Trogontherium cuvieri).
Los bosques del norte de la época eran frecuentados por el alce de cara ancha, el ciervo gigante y el corzo, así como por nobles ejemplares de ciervo (Cervus elaphus). En los bosques abiertos y prados, el ganado salvaje (Bos primigenius) comenzó a ser más numeroso [ p. 95 ] y también abundaba el bisonte (Bison priscus). Entre las formas que frecuentaban prados o bosques se encontraban caballos de mayor tamaño, como los caballos de Mosbach y de Süssenborn. En este conjunto de tipos norteños y meridionales, cabe destacar que los tipos de mamíferos de bosque y prado euroasiáticos predominan en gran medida en número y variedad sobre los tipos africano-asiáticos; esto, junto con la flora, es una indicación de que el clima era de carácter templado; Es probable, por lo tanto, que todos los mamíferos estuvieran bien protegidos con una cubierta pilosa y adaptados a un clima templado. El hecho de que la fauna en su conjunto permaneciera prácticamente inalterada durante la segunda glaciación demuestra, no que migrara al sur y luego regresara, sino que las regiones no glaciadas de Europa occidental eran templadas, no frías.
¶ La carrera de Heidelberg
Hombre de Heidelberg. Elefante antiguo. Rinoceronte etrusco. Caballo de Mosbach. Jabalí. Alce de cara ancha. Ciervo común. Corzo. Bisonte primitivo (bisonte europeo). Uro primitivo. Oso de Auvernia. Oso de Deninger. León. Gato montés. Lobo. Castor.
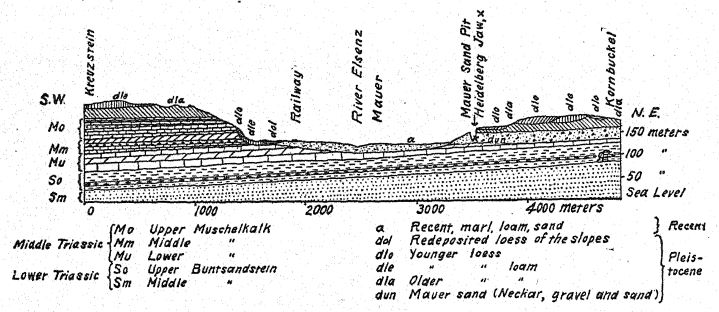
Para nosotros, la vida mamífera más interesante es, con diferencia, la que se encuentra al sur de la desembocadura del Neckar, a lo largo del antiguo arroyo Elsenz, donde se depositaron las «arenas de Mauer» inferiores, que contienen la mandíbula inferior del hombre de Heidelberg y los restos de numerosos animales de la época. La enumeración de toda esta fauna es fundamental, ya que indica las condiciones climáticas templadas que rodearon a la primera especie auténtica de hombre descubierta hasta la fecha en Europa. El descubridor, Schoetensack,35 atribuyó estos mamíferos y al hombre de Heidelberg al Primer Interglaciar, y Geikie ha expresado recientemente una opinión similar. La presencia del rinoceronte etrusco parecería indicar una antigüedad tan grande, pero la evidencia aportada por este animal primitivo se ve eclipsada por la de tres mamíferos muy característicos del Segundo Interglaciar: el rinoceronte de colmillos rectos o [ p. 96 ] el antiguo elefante (E. antiquus), el león y el caballo de Mosbach. Con la única excepción del rinoceronte etrusco, todas estas especies, que frecuentaban el antiguo arroyo Elsenz y se depositaron en las «arenas de Mauer», también se encontraban en los bosques y prados de la región hoy conocida como Baden, donde se encuentran los yacimientos de mamíferos fósiles de Mosbach, cerca del Neckar. Una vida mamífera similar, de una época algo más reciente, se encuentra en las gravas fluviales de Süssenborn, cerca de Weimar. Los caballos de Mauer, de Mosbach y de Süssenborn[4] eran mucho más grandes y de características más especializadas que el caballo de Steno del Primer Interglaciar.
Así, los Heidelberg, la primera raza humana registrada en Europa occidental, aparecen en el sur de Alemania a principios del Segundo Interglaciar, en medio de una imponente fauna de mamíferos de aspecto septentrional y que contiene muchas especies que viven en los bosques, como osos, ciervos y alces; en los prados y bosques ramoneaba el elefante gigante de colmillos rectos (E. antiquus), que por la simple estructura de sus dientes de molienda se considera similar en hábitos al elefante africano que ahora habita los bosques de África central; la presencia de este animal indica un clima relativamente húmedo y un país bien boscoso. El rinoceronte etrusco [ p. 97 ] se diferenciaba de la forma más grande de Merck en la posesión de dientes de molienda de corona relativamente corta, adaptados a los hábitos de ramoneo y a un país boscoso; en la cabeza tenía dos cuernos; era un tipo de extremidades largas y de movimientos rápidos; Las manadas de bisontes y de ganado salvaje (urus) que vagaban por las llanuras estaban ahora sujetas al ataque del león.

El descubrimiento en 1907 de una mandíbula inferior humana en la base de las arenas de Mauer es uno de los más importantes en la historia de la antropología. El hallazgo se realizó a una profundidad de 24,10 m (79 pies) desde la superficie superior de un alto acantilado (Fig. 46), en antiguas arenas de río que desde hacía tiempo se sabía que albergaban la antiquísima fauna mamífera descrita anteriormente. Durante años, los trabajadores habían recibido instrucciones de vigilar atentamente la presencia de restos humanos. La mandíbula, evidentemente, se había deslizado con las arenas del río y se había separado del cráneo, pero se mantenía en perfecto estado de conservación. Puede citarse primero la descripción del autor.36 La mandíbula muestra una combinación de características nunca antes encontradas en ningún fósil ni en el hombre reciente. La protuberancia de la mandíbula inferior, justo debajo de los dientes frontales, que da forma al mentón humano, está completamente ausente. Si los dientes hubieran estado ausentes, habría sido imposible diagnosticarlo como humano. A partir de un fragmento de la sínfisis de la mandíbula, bien podría haberse clasificado como un antropoide similar a un gorila, mientras que la rama ascendente se asemeja a la de una gran variedad de gibón. La certeza absoluta de que estos restos son humanos se basa en la forma de los dientes: molares, premolares, caninos e incisivos son esencialmente humanos y, aunque de forma algo primitiva, no muestran rastros de ser intermedios entre el hombre y los simios antropoides, sino más bien de derivar de un ancestro común más antiguo. Sin embargo, los dientes son algo pequeños para la mandíbula; el tamaño del borde permitiría el desarrollo de dientes mucho más grandes; solo podemos concluir que no se sometió a una gran tensión sobre los dientes y, por lo tanto, el poderoso desarrollo de los huesos de la mandíbula no fue diseñado para su beneficio. La conclusión es que la mandíbula, considerada indudablemente humana por la naturaleza de sus dientes, se sitúa cerca del punto de separación entre el hombre y los simios antropoides. En comparación con las mandíbulas de las razas neandertales, halladas en Spy (Bélgica) y en Krapina (Croacia), podemos considerar la mandíbula de Heidelberg como preneandertaloides; de hecho, es un tipo generalizado.
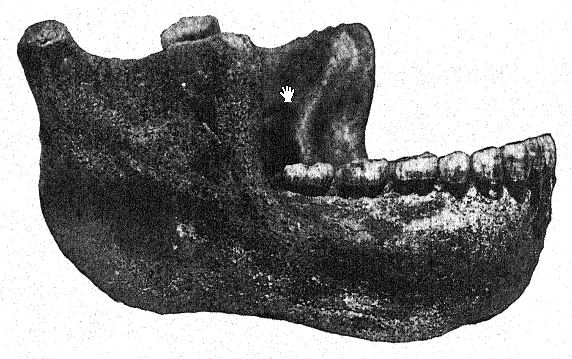
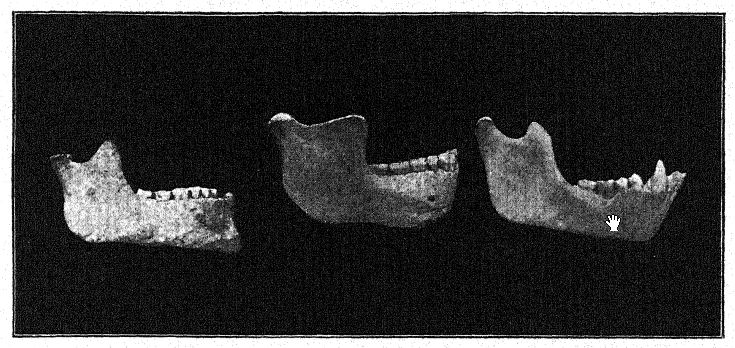
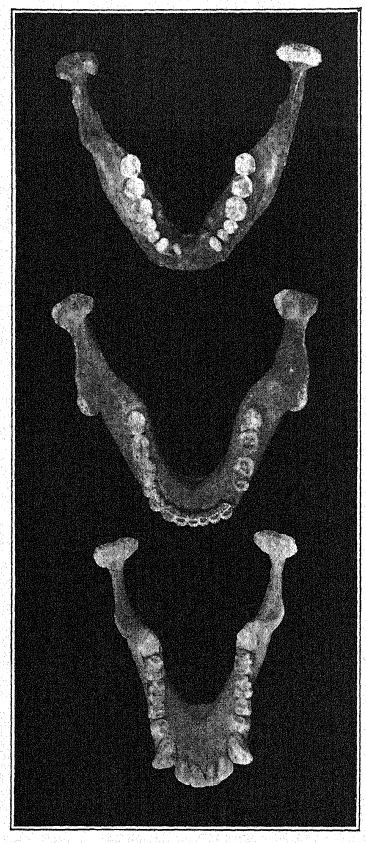
Con un espíritu conservador, Schoetensack denominó al tipo representado por esta mandíbula Homo heidelbergensis. Otros autores lo han considerado de un rango genérico distinto; por ello, Bonarelli lo denominó Palaoanthropus heidelbergensis.37 La mandíbula en sí es extremadamente maciza; los caninos, a diferencia de los de los simios antropoides y de la raza de Piltdown, no sobresalen de la línea de los demás dientes y, por lo tanto, no se utilizaban como armas de ataque y defensa como en los antropoides, en quienes estos dientes [ p. 100 ] están prominentemente desarrollados como colmillos. Como señaló Schoetensack, los dientes no son muy macizos en proporción a la mandíbula, que es la mandíbula humana más poderosa conocida, incluso superando a la mandíbula esquimal más grande, lo que indica un cráneo de carácter muy macizo y primitivo. Se asemeja a la del simio en la recesión del mentón, de ahí que se le haya denominado amentalis. Presenta un gran desarrollo de la apófisis coronoides de la mandíbula para la inserción del músculo temporal. Es posible que esta mandíbula se utilizara como herramienta en las últimas etapas de la preparación de pieles, como era práctica habitual entre los esquimales. Observamos que las poderosas ramas óseas de la mandíbula, vistas desde arriba, se cierran sobre el espacio dejado para la lengua; de hecho, el hueso se cierra tanto que interfiere gravemente con el libre uso de la lengua para el habla articulada.
Parecería que en la mandíbula, y probablemente en todas las demás características del cráneo, a medida que se conozcan, la raza de Heidelberg se encontrará como un neandertal en desarrollo, es decir, una forma ancestral primitiva, más poderosa y más parecida a un simio. En cuanto al mentón retraído, los verdaderos neandertales de Spy, Malarnaud, Krapina y La Chapelle se sitúan exactamente a medio camino entre las razas más inferiores del hombre reciente y los simios antropoides.
No solo entre los esquimales, sino en general entre las razas salvajes de Australia y otros países, las mandíbulas se utilizan como herramientas; entre los australianos, los dientes están muy desgastados, pero se conservan admirablemente. Vistos desde arriba, observamos que los dientes de Heidelberg forman un arco perfecto, o una disposición en forma de herradura, mientras que en todos los simios las dos líneas de dientes son casi paralelas. Así pues, si bien puede haber amplias diferencias de opinión respecto a las relaciones del hombre de Heidelberg, todos coinciden en que el descubrimiento de Schoetensack nos ofrece uno de los grandes eslabones o tipos perdidos en la cadena del desarrollo humano.

La vida típica de los mamíferos del Segundo Interglaciar, tal como se encuentra en Mosbach y Sussenborn, pertenece quizás a una etapa algo más reciente del Segundo Interglaciar que la de las arenas de Mauer, pues en estas localidades escasea el rinoceronte etrusco [ p. 102 ] y abunda el rinoceronte de hocico ancho, más especializado. Este animal se diferencia de la forma etrusca en la posesión de dientes de molienda de corona relativamente larga, mejor adaptados al pastoreo. En la cabeza se encontraban dos cuernos. Una variedad del mamut del sur (E. trogontherii) es tan característica del Segundo Interglaciar que Pohlig se refiere a este período de vida como la etapa E. trogontherii. Por la estructura de sus dientes, se considera similar en hábitos al elefante asiático, que ahora habita los bosques de la India, pero presenta la peculiar frente cóncava distintiva del mamut y muy diferente de la frente convexa del elefante indio. Los osos de este período pertenecen a las especies primitivas U. deningeri y U. arvernensis, pues hasta la fecha no existe un registro certero de la presencia del auténtico oso pardo europeo (U. arctos). El tigre dientes de sable de esta época se conserva en las cavernas de los Pirineos, cerca de Montmaurin, asociado con los restos de la hiena rayada (H. striata), una especie que se distribuyó ampliamente por Europa occidental a principios del Pleistoceno. Esta especie fue contemporánea de la hiena moteada (H. crocuta), y posteriormente reemplazada por ella, de la cual descendió la muy resistente hiena cavernaria (H. crocuta spelcea) del «Período del Reno». Observamos que el ciervo policladino del Plioceno Superior y la Primera Glaciación ha desaparecido de Europa occidental; tampoco existen rastros del ciervo axis. El hipopótamo está representado por la especie gigante H. major.
¶ Primeras migraciones del reno hacia el norte
Los animales que hemos descrito pertenecen a las regiones más cálidas y templadas de Europa. En las regiones cercanas a los glaciares ya se encontraba el reno; de hecho, este animal típicamente nórdico está documentado en las gravas de Süssenborn, cerca de Weimar.
Existe evidencia de una sucesión de cambios climáticos en la región de Heidelberg. La mandíbula de Heidelberg, con su fauna de mamíferos de clima templado, se encontraba en la misma base del acantilado de Mauer. [ p. 103 ] Pero más arriba en el acantilado (Fig. 46), en un nivel correspondiente, se encuentran restos de mamíferos que indican un marcado descenso de la temperatura y que algunos expertos atribuyen al período de clima frío que caracterizó el norte de Europa hacia el final del Segundo Interglaciar. El reno también se encuentra en las gravas de las terrazas altas del río Murr, cerca de Steinheim; así, en Mauer, Sussenbom y Steinheim, encontramos pruebas de que el reno había comenzado a extenderse por las regiones más frías de Europa, y existen indicios de que llegó incluso hasta los Pirineos, tan al sur.
La evidencia del primer período frío y árido, que en su momento afectó considerablemente el clima de Europa occidental, también se encuentra en la capa del llamado «loess antiguo», que yace en el acantilado sobre las «arenas de Mauer». Este loess cubre los cálidos depósitos de mamíferos de las «arenas de Mosbach», así como las «terrazas altas» de muchos de los antiguos valles fluviales. Tanto en Europa como en América, la secuencia climática de la Segunda Etapa Interglacial, de húmedo a seco, parece haber sido la misma.
Así, tras la retirada de los campos de hielo de la segunda glaciación, el clima fue inicialmente frío y húmedo; luego siguió una larga etapa cálida, propicia para la expansión de los bosques; a esta le siguió finalmente un período de aridez en el que se formaron los depósitos de loess más antiguos. En Rusia, la tercera glaciación también estuvo precedida por un clima árido y estepario, con fuertes vientos favorables al transporte de loess.
No se han encontrado paleolitos ni otras pruebas de ocupación humana en este período frío y seco, ya que no existe evidencia en ninguna parte de Europa de campamentos en este «loess antiguo», como los que encontramos en el «loess» depositado durante un período árido similar hacia el final del Tercer Interglaciar y de nuevo durante el Postglaciar. Tampoco tenemos registro de vida de mamíferos en este «loess antiguo» de Europa.
[ pág. 104 ]
¶ La Tercera Glaciación
Este período árido en el norte de Europa y Norteamérica fue seguido por el clima húmedo y frío de la tercera glaciación. Penck estima que el avance de estos nuevos campos de hielo comenzó hace 120.000 años y que el período de avance y retroceso de los glaciares fue de al menos 20.000 años. En los Alpes, la línea de nieve descendió 1.250 metros por debajo del nivel actual; en consecuencia, esta glaciación fue más severa que la primera, pero algo menos severa que la segunda. En el norte de Europa, el campo de hielo escandinavo no cubrió un área tan extensa como durante la segunda glaciación, aunque Gran Bretaña y Escandinavia volvieron a quedar profundamente sepultadas por el hielo; el manto glaciar y los glaciares fluyeron en dirección oeste y suroeste a través de Dinamarca y la parte sur de la cuenca del Báltico hacia Holanda y el norte de Alemania. En los Alpes, la tercera glaciación envió enormes témpanos de hielo a lo largo del valle del Rin, hacia el este de Francia y el valle del Po, donde esta glaciación fue aún más extensa que la segunda. Pero el glaciar más grande de esta época fue el del Isar, un afluente meridional del Danubio, que nace en los Alpes bávaros.
Durante la Tercera Etapa Glacial, se formaron algunas de las terrazas intermedias a lo largo del Rin y otros ríos que fluyen desde los Alpes. En Gran Bretaña,39, mientras que durante la segunda glaciación los campos de hielo se extendían hasta el Támesis al sur, durante la tercera glaciación no se extendieron más allá de la región central; sin embargo, prevaleció un clima ártico en el sur de Inglaterra, con condiciones y temperaturas propias de la tundra, como lo indican los depósitos vegetales de Hoxne40 en Suffolk. Incluso antes del inicio de la tercera glaciación en Europa, se había formado un gran casquete glaciar sobre el Labrador, en la costa oriental de Norteamérica, y las capas de hielo que fluían hacia el sur y el suroeste se extendieron hasta Illinois, depositando las grandes derivas ilinoianas.
[ pág. 105 ]
A lo largo de los límites de estos grandes campos de hielo, en ambos países, prevalecía un clima frío y húmedo, ya que una condición fundamental de la glaciación es la fuerte precipitación de nieve. En el norte de Europa, entre los grandes campos de hielo alpinos y escandinavos de la tercera glaciación, sin duda prevaleció un clima frío; en la región del río Neckar, cerca de Cannstatt, se encuentra un depósito conocido como marga de mamut, que Koken creía contemporáneo del período de la tercera glaciación, aunque la evidencia no es convincente.41 Aquí se encuentran restos fósiles del reno abedul, así como de dos importantes recién llegados a Europa procedentes de las regiones de tundra del extremo noreste, ar,imglg [ p. 106 ] que habían vagado por las fronteras meridionales de la capa de hielo escandinava desde las tundras del norte de Rusia y Siberia. Esta es la primera aparición en Europa occidental del mamut lanudo (E. primigenius) y del rinoceronte lanudo (D. antiquitatis). En esta «tierra arcillosa» también se encuentran dos especies de caballo: el ciervo gigante (Megaceros), el ciervo macho, el bisonte europeo y el uro. Si el mamut lanudo y el rinoceronte lanudo entraron en Alemania oriental en esta época, sin duda se retiraron hacia el norte con la llegada del clima templado cálido de la Tercera Etapa Interglacial, ya que no se ha vuelto a encontrar rastro de estos animales en Europa hasta la llegada de la cuarta glaciación.
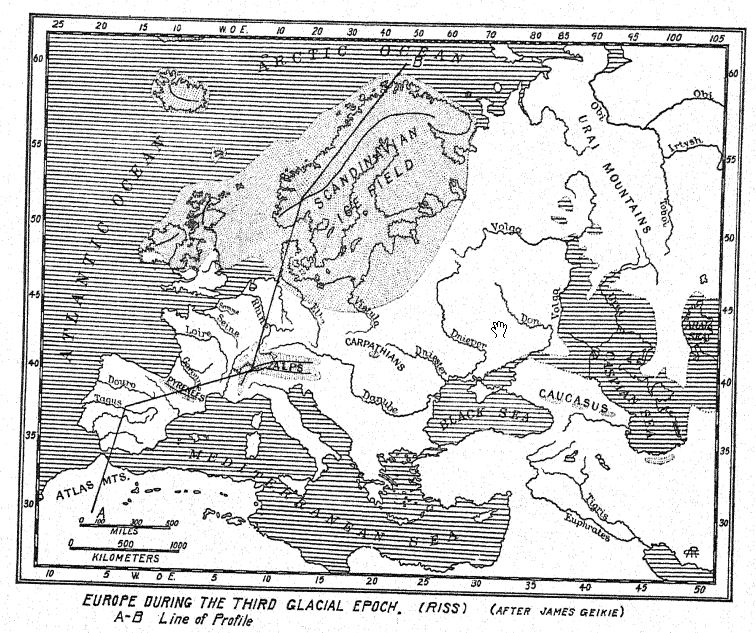
¶ Bibliografía
(1) Gaudry, 1890.1.
(2) Smith, GE, 1912.1, pág. 582.
(3) Op. cit.
(4) Munro, 1893.1.
(5) Osborn, 1910.1, págs. 306, 307.
(6) Geikie, J., 1894.1, págs. 329-336; 1914.1, pág. 227.
(7) Dawkins, 1883.1, págs. 576-579.
(8) Geikie, J., 1914.1, pág. 248.
(9) Reid, C., 1908.1.
(10) Desnoyers, 1863.1.
(11) Haug, 1911.1, pág. 1507.
(12) Geikie, J., 1894.1, pág. 682; 1914.1, pág. 250.
(13) Dubois, 1894.1.
(14) Fischer, 1 913. 1.
(15) Schwalbe, 1899.1; 1914.1.
(16) Büchner, 1914.1.
(17) Volz, 1907. 1.
(18) Elbert, 1908.1.
(19) Selenka, 1911.1.
(20) Peregrino, 1913.1.
(21) Schwalbe, 1899.1, págs. 227, 228. (22) Op. cit., pág. 223.
(23) Schwalbe, 1914. i, págs. 601-606.
(24) Büchner, 1914.1, pág. 129.
(25) Dubois, 1894,1, pág. 14.
(26) Keith, 1912. 1.
(27) Smith, GE, 1912. 1, pág. 595.
(28) Op. cit.
(29) Rutot, 1907.1.
(30) Boule, 1913. 1, págs. 266, 267.
(31) MacCurdy, 1905.1, págs. 468, 469.
(32) Geikie, J., 1914.1, pág. 251.
(33) Op. cit. pág. 255.
(34) Op. cit. pág. 238.
(35) Schoetensack, 1908.1.
(36) Op. cit. págs. 25-43.
(37) Bonarelli, 1909.1.
(38) Penck, 1909. 1.
(39) Geikie, J., 1914.1, pág. 258.
(40) Op. cit., págs. 257-262.
(41) Schmidt, 1912.1, pág. 181
¶ Notas al pie
Un artículo reciente de A. Smith Woodward describe el cuarto espécimen conocido de Dryopithecus, descubierto recientemente en el norte de España (véase Woodward, 1914.2). ↩︎
Existe una vasta literatura sobre Pithecanthropus. La utilizada principalmente en la presente descripción incluye a Dubois,13 Fischer14 Schwalbe,15 Buchnerd16. ↩︎
En el cráneo de Trinil restaurado por McGregor (Fig. 36) la capacidad craneal es de 900 c. cm. ↩︎
Estos caballos ahora se identifican respectivamente como E. mauerensis, E. moshachensis y E. süssenbornensis. ↩︎
Esta glaciación, tal como se produce en el norte de Europa, ha sido denominada Polandian por Geikie; en los Alpes, Penck la ha denominado Ilinoian; en América se la conoce como Ilinoian debido a las grandes acumulaciones que depositó sobre el estado de Illinois. ↩︎