| XXIX. Le charbon et sa présence dans la nature | Page de titre | XXXI. Le Permien et son climat glaciaire |
[ p. 405 ]
Au Paléozoïque moyen, et plus particulièrement au Dévonien, lorsque les terres émergées furent entièrement recouvertes de végétation – substrat indispensable à l’alimentation de tous les animaux –, un habitat propice à la vie animale se développa. Cet habitat fut colonisé par les vers, les escargots, les araignées, les insectes et autres invertébrés, puis par divers groupes de vertébrés. Parmi ces derniers, les premiers à trouver la terre ferme habitable furent les amphibiens ancestraux, qui régnèrent sur leurs milieux respectifs du Dévonien supérieur jusqu’au Pennsylvanien. De ces animaux naquirent des espèces plus complexes, une extraordinaire variété de reptiles, qui non seulement devinrent les maîtres des terres, des mers et des océans avant la fin du Permien, mais envahirent peu après les airs, et dont la domination valut au monde mésozoïque, ou médiéval, le surnom d’« Âge des Reptiles ».
¶ Amphibiens, vivants et fossiles
Les amphibiens se distinguent des poissons par la présence de pattes et non de nageoires, chaque patte étant munie de doigts. Presque tous respirent par des branchies lorsqu’ils sont très jeunes et peuvent conserver ces organes tout au long de leur vie ; les poumons et les narines fonctionnelles sont cependant presque toujours présents à l’âge adulte, bien que chez les salamandres, les membres puissent être réduits à des vestiges ou complètement supprimés (voir Pl., p. 407, Fig. 1-4). Ils possèdent un cœur à trois cavités et une langue musculaire mobile.
Amphibiens vivants. — Il existe environ neuf cents espèces d’amphibiens vivants, principalement des grenouilles (Fig., p. 406). Grenouilles, crapauds, tritons, sirènes, nectures, doryphores et salamandres terrestres sont des exemples d’amphibiens vivants (Pl., p. 407). Tous sont des animaux à sang froid. Le nom de classe Amphibia, qui signifie « qui mènent une double vie », leur a été donné car beaucoup vivent à la fois sur terre et en eau douce. D’autres, cependant, vivent exclusivement dans l’eau et, en règle générale, tous les amphibiens, à leurs premiers stades de développement, sont entièrement confinés à ce milieu.
Chez les grenouilles et les crapauds, les sexes sont toujours séparés et, dans la grande majorité des espèces, les [ p. 406 ] petits œufs sont fécondés dans l’eau et s’y développent sans aucun soin parental. Le développement est donc très semblable à celui des poissons et très différent de celui des vertébrés supérieurs. Les œufs de grenouilles et de crapauds, par exemple, donnent naissance à de petits animaux communément appelés têtards, dotés d’une tête et d’un corps plus ou moins gros et arrondis, se terminant par une longue queue très flexible qu’ils agitent en nageant, comme les poissons (voir Fig. ci-dessous). Ces têtards possèdent des branchies qui, au début, font saillie sur les côtés de la tête, mais qui sont ensuite recouvertes par un opercule. Au bout de deux mois environ, ils atteignent un stade équivalent à celui des dipneustes. Ils subissent alors une métamorphose marquée : les pattes postérieures apparaissent en premier, puis les pattes antérieures, cachées sous l’opercule. La longue queue se raccourcit par résorption interne et les branchies sont également résorbées ou tombent. Les poumons apparaissent ensuite, et pendant un certain temps, les jeunes créatures sont pleinement amphibies, respirant l’eau par leurs branchies et en extrayant l’oxygène libre, tout en inspirant également [ p. 408 ] l’air par leurs poumons ; mais [ p. 407 ] bientôt, les petites grenouilles ou les crapauds s’installent sur la terre ferme et ne respirent plus que de l’air (voir Fig. ci-dessus). Dans cette métamorphose, qui dure de quelques semaines à quelques mois tout au plus, nous assistons à une récapitulation de l’histoire paléozoïque, qui a nécessité des millénaires de sélection des individus les mieux adaptés à leur environnement. Chez les formes qui restent en permanence dans l’eau, la transformation est moins importante, et certaines d’entre elles ressemblent physiquement à des poissons tout au long de leur vie, parmi lesquelles les nectures et les salamandres des rivières des vallées de l’Ohio et de l’Alabama (voir Pl., p. 407, Fig. 1).
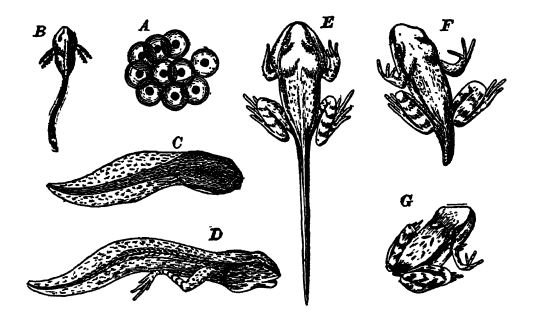
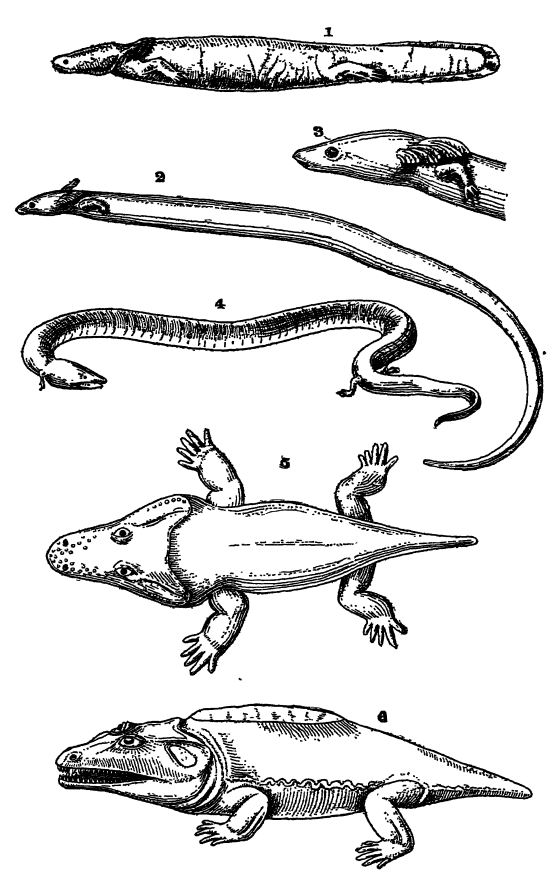
Fig. 1, salamandre à taches noires (Necturus maculaius) des rivières Ohio et Mississippi ; 2 et 3, salamandre à taches noires (Siren lacertina), une petite salamandre, notez les trois paires de branchies et l’absence de pattes postérieures ; 4, amphium à trois branches (Amphiuma tridactyla), avec branchies et poumons ; 5, trématops milleri, un stégocéphale de 90 cm de long, du Texas ; 6, cacops aspidephorus, un stégocéphale de 50 cm de long, du Texas. Fig. 1 et 4 d’après Parker et Haswell ; Fig. 2 et 3 d’après Lankester. Guide du British Museum ; Fig. 5 et 6 d’après
Respiration. — Tous les amphibiens, à l’état juvénile, possèdent deux ou trois paires de branchies externes, ou des branchies internes avec fentes branchiales externes, de petites excroissances plumeuses situées à l’arrière de la tête et riches en vaisseaux sanguins (voir Pl., p. 407, Fig. 1-3). Ces branchies sont également présentes chez les dipneustes, et chez les sirènes et les nectures, elles peuvent persister toute la vie, bien que ce soit plutôt exceptionnel. Chez les salamandres terrestres et chez tous les amphibiens sans queue (grenouilles, etc.), les branchies disparaissent et la respiration adulte est assurée exclusivement par les membres, comme chez les vertébrés supérieurs. Tant que les branchies sont présentes, les voies aériennes menant aux poumons par le nez ne débouchent pas dans la bouche ; elles prennent cette position dès que les branchies disparaissent et que les poumons deviennent fonctionnels. L’air est alors inspiré par les narines et, chez les grenouilles, propulsé dans les poumons par un mouvement de déglutition. De nombreux amphibiens peuvent survivre longtemps sans se nourrir, et dans les régions froides, tous les animaux terrestres hibernent durant l’hiver dans la vase au fond des cours d’eau et des étangs, respirant alors uniquement par la peau.
Ouïe. — Chez les poissons, il n’existe pas d’oreilles fonctionnelles, bien qu’ils possèdent des organes auditifs internes ; chez les amphibiens, en revanche, on observe des organes auditifs externes distincts. Ceux-ci sont particulièrement visibles chez les grenouilles, où la membrane tympanique du tympan est un disque plus ou moins circulaire, enchâssé dans la peau sur les côtés de la tête. La cavité située derrière ce disque, ou tympan, communique avec la partie postérieure de la bouche par un conduit appelé trompe d’Eustache, également présent chez tous les vertébrés supérieurs.
Classification. — Les formes vivantes sont divisées en deux sous-classes selon leur morphologie : les formes adultes dépourvues de queue sont appelées Anoures (voir Fig., p. 406), tandis que les formes primitives conservant une queue sont appelées Urodelas (voir Pl., p. 407, Fig. 1-4). Les amphibiens du Paléozoïque, qui seront abordés plus loin, sont regroupés sous le terme de Stégocéphales.
Les Anoures comprennent les grenouilles et les crapauds que nous connaissons bien. Ils possèdent toujours quatre pattes, dont les pattes postérieures sont longues et puissantes, et les adultes sont dépourvus de branchies externes. On les trouve presque partout dans le monde, à l’exception des régions polaires. Cette sous-classe est d’origine relativement récente, puisque le plus ancien fossile connu date [ p. 409 ] du Jurassique supérieur, bien qu’ils soient beaucoup plus fréquents dans les strates du Cénozoïque. Ils constituent le groupe d’amphibiens vivants le plus diversifié et le plus spécialisé, mais ne présentent pas un intérêt particulier en géologie historique.
Les Urodèles, ou amphibiens à queue, sont d’une importance capitale à cet égard. Bien qu’aucun fossile antérieur au Jurassique ne soit connu, cette sous-classe doit nécessairement avoir son origine chez les Stégocéphales du Paléozoïque. Les Urodèles, aujourd’hui pratiquement confinés aux régions tempérées de l’hémisphère nord, sont des amphibiens qui, à l’âge adulte (salamandres, tritons, sirènes, nectures et axolotls), présentent souvent une apparence étonnamment piscifère (voir Pl., p. 407, Fig. 1). Chez les nectures et les axolotls, on observe une nageoire impaire sur la face dorsale, qui se prolonge autour de la queue diphycerque et le long de la partie postérieure de la face ventrale, comme chez les dipneustes. Cependant, cette nageoire ne possède pas de rayons osseux, contrairement à celle des poissons. Bien que les urodèles ressemblent à des poissons, ils se distinguent aisément par le fait que leurs appendices pairs ne sont pas des nageoires, mais des pattes pleinement développées, quoique petites et faibles, présentant toutes les caractéristiques squelettiques essentielles des membres des vertébrés supérieurs (voir Pl., p. 407, fig. 1-4). En règle générale, on compte deux paires de ces membres : les antérieurs ou pectoraux et les postérieurs ou pelviens. Chez certaines espèces, cependant, seule la paire antérieure est présente, la paire postérieure ayant disparu, et chez quelques espèces, tous les membres sont absents ; il s’agit des cseciliens fouisseurs, vermiformes, des régions tropicales. Le nombre d’orteils à chaque patte varie entre cinq, le nombre habituel, et deux. Certaines espèces, comme les salamandres terrestres, peuvent, à maturité, vivre exclusivement sur terre et perdre toute trace de branchies, tandis que d’autres, comme les nectures, restent aquatiques et conservent leurs branchies tout au long de leur vie. Le necture géant du Japon et de Chine est le plus grand de tous les urodèles, atteignant une longueur de 1,5 mètre. Un individu a vécu en captivité pendant plus de cinquante ans.
Stégocéphales, les amphibiens du Paléozoïque. — Le terme « Stégocéphales » signifie « tête couverte » ou « armure » et fait référence au fait que la surface supérieure du crâne était recouverte d’une épaisse couche d’os dermiques. On les appelle aussi amphibiens à crâne solide et cuirassés, et c’est cette armure qui distingue immédiatement les formes du Paléozoïque et du Trias de toutes les formes actuelles. La cage thoracique était également dotée d’une armure dermique, composée de trois grandes plaques sculptées qui représentaient une partie des os de l’arc pectoral des poissons et de la ceinture scapulaire des vertébrés supérieurs. Chez certaines formes, le corps entier était recouvert de petites écailles [ p. 410 ] imbriquées, celles de la face ventrale étant les plus épaisses, ou bien les écailles étaient présentes uniquement sur la face ventrale, tandis que d’autres (Branchiosauria) étaient essentiellement nus et dépourvus d’écailles. Presque tous les amphibiens vivants sont nus et les écailles sont rarement présentes (Cseciliens). Un groupe des formes anciennes a été appelé Labyrinthodontes, car la dentine des grandes dents coniques était très pliée en ondulations labyrinthiques telles que celles que l’on voit pour la première fois chez les poissons (p. 298).
La première trace d’un vertébré à membres, probablement de type stégocéphalien, est une empreinte de pied (Thinopus) du Dévonien supérieur (Fig., p. 331), mais les squelettes de ces amphibiens cuirassés ne sont connus en Amérique du Nord qu’à partir du Pennsylvanien. Ils étaient alors présents, ainsi qu’au Permien, à leur apogée. L’ensemble de l’espèce a disparu avant la fin du Trias.
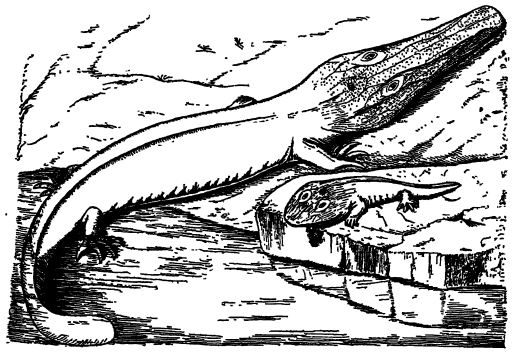
Les amphibiens les plus primitifs possédaient une petite tête et des membres faibles sur un corps allongé se terminant par une queue propulsive semblable à celle des salamandres modernes. Contrairement à ces dernières, ils conservaient cependant plus ou moins l’armure corporelle héritée de leurs ancêtres poissons à écailles et cuirassés. Les formes à grosse tête, observées principalement à la fin du Pennsylvanien et au début du Permien, étaient des descendants précoces de formes primitives. L’évolution marquée des amphibiens anciens coïncide avec leur déploiement dans tous les environnements [ p. 411 ] variés, passant des milieux aquatiques et marécageux aux milieux de transition, puis aux terres émergées, où ils se diversifièrent encore davantage en s’enfouissant sous la surface et en s’élevant vers les arbres.
On connaît de nombreuses espèces de stégocéphales, les gisements de charbon d’Amérique du Nord à eux seuls en ayant livré quatre-vingt-dix. Leur taille varie d’environ 2,5 cm à plus de 3 m (Fig., p. 360). Certains ressemblaient à des lézards : petits et actifs, avec des pattes bien développées et une queue relativement courte ; d’autres étaient d’excellents nageurs, dotés d’une longue queue ; d’autres encore, de taille moyenne, étaient trapus et d’un tempérament lent, à l’instar des crocodiles (Fig., p. 410) ; tandis que quelques-uns évoquaient des têtards gigantesques, presque sans pattes, se tortillant dans l’eau. La présence d’une queue était invariablement la norme.
Chez la grande majorité des Stégocéphales, on observait deux paires de membres, bien que des formes apodes, comme les serpents, soient également connues (voir Fig., p. 360). Les membres étaient bien développés, mais courts et trapus (voir Fig., p. 410, et Pl., p. 407, Fig. 5, 6). Les os des pattes antérieures étaient semblables à ceux des salamandres actuelles par leur nombre, leur forme et leur disposition, et les mains possédaient quatre, ou plus rarement cinq, doigts généralement courts. Les membres postérieurs étaient presque toujours plus longs et plus robustes, et portaient cinq orteils, dont le deuxième ou le troisième était le plus long.
La tête était souvent large et plate, avec une gueule très large occupant presque toute la largeur du crâne, comme chez les grenouilles et les salamandres ; mais chez certaines espèces, la tête était plus ou moins allongée, voire pointue, comme chez les crocodiles. Cependant, la tête de ces derniers est également large chez les jeunes individus, ce qui suggère que la tête large est le type primitif. Parmi les premiers, l’Eryops du Permien du Texas possédait une tête de 60 cm de long et 45 cm de large et ressemblait à un énorme têtard : une tête large et plate, pas de cou, un corps massif et trapu, des pattes courtes et une queue lourde et aplatie (voir Branchiosaurus, fig., p. 410). Comme l’a dit Huxley, ils « erraient avec un gros ventre et peu de pattes, tels Falstaff dans sa vieillesse, au milieu des forêts de charbon ».
Les Stégocéphales vivaient probablement tous soit en eau douce, soit sur la terre ferme, et certains se nourrissaient sur les plages des mers du Trias. On a retrouvé des spécimens de petite taille et actifs dans des troncs creux du Pennsylvanien de Nouvelle-Écosse. La grande majorité des Stégocéphales étaient probablement carnivores et se nourrissaient de coquillages, de vers et d’autres invertébrés aquatiques, mais plus particulièrement de poissons, de reptiles et de petits membres de leur propre espèce.
Le troisième œil des stégocéphales. — Chez tous les stégocéphales bien conservés, les os du toit crânien et ceux du crâne proprement dit étaient percés non seulement par les grandes orbites latérales où se situaient les yeux et par les deux orifices nasaux antérieurs, mais aussi par un unique petit orifice traversant l’os recouvrant [ p. 412 ] le cerveau (voir Fig., p. 410). Cet orifice est d’un grand intérêt, car il abritait un troisième œil appelé œil pinéal. On retrouve un tel orifice chez de nombreux reptiles fossiles et certains reptiles actuels (Sphenodon). Bien que l’organe qu’il contient ne puisse guère être considéré comme fonctionnant comme un véritable œil chez les reptiles actuels, son importance réside dans le fait qu’il s’agit d’un organe vestigial dont l’origine remonte au moins au Pennsylvanien. Les rudiments de cet œil sont présents dans le cerveau de tous les vertébrés actuels, y compris l’homme.
Ascendance des Stégocéphales. — Il devient de plus en plus difficile de distinguer les reptiles du Paléozoïque supérieur de leurs apparentés stégocéphales. Les plus anciens reptiles à tête pleine (Cotylosauria et Pareiasauria) sont très proches des Stégocéphales à tête pleine. À propos de ce type, Osborn déclare : « Os par os, ses différentes parties indiquent une descendance commune du type de crâne des poissons à franges (Crossopterygia). » En d’autres termes, les preuves s’accumulent rapidement pour montrer que les stégocéphales n’étaient pas les vertébrés terrestres à membres les plus primitifs, mais qu’ils descendaient d’une souche plus ancienne (Protopoda), qui a donné naissance d’une part aux amphibiens aquatiques et d’autre part aux reptiles, qui se sont complètement adaptés à la vie terrestre.
¶ Reptiles, vivants et fossiles
Chez tous les vertébrés étudiés jusqu’à présent — poissons et amphibiens —, nous avons constaté que leur habitat est soit entièrement aquatique, soit qu’à tout le moins les petits œufs y sont pondus et fécondés, et que les jeunes y naissent et y passent leur jeunesse. Tous les vertébrés supérieurs s’éloignent progressivement de ce milieu et aucun ne s’y développe directement à partir de l’œuf. Autrement dit, les reptiles sont généralement ovipares : ils pondent de gros œufs, semblables à ceux des oiseaux, contenant une quantité importante de vitellus, et ces œufs sont fécondés avant d’être déposés sur la terre ferme, où ils éclosent sous la chaleur du soleil. C’est là la différence la plus importante et fondamentale entre les vertébrés inférieurs (poissons et amphibiens) d’une part, et les vertébrés supérieurs (reptiles, oiseaux et mammifères) d’autre part.
Reptiles vivants. — Tous les animaux vivants connus sous les noms de tortues, lézards, serpents, alligators, crocodiles, ainsi que les espèces disparues, comme les dinosaures géants du Mésozoïque et les ptérodactyles, appartiennent à la classe des Reptiles. Le mot « reptile » signifie « qui rampe » ou « qui se déplace en rampant » et désigne un animal qui se déplace sur le ventre, comme le serpent, ou qui se déplace difficilement sur ses pattes courtes et écartées, comme l’alligator. Cependant, de nombreux reptiles ne sont en aucun cas des animaux rampants ou qui se déplacent en rampant, comme par exemple de nombreux lézards rapides, certains dinosaures aux pattes massives et les ptérodactyles.
Tous les reptiles vivants sont des animaux à sang froid, comme les poissons et les amphibiens. Leur peau n’est jamais [ p. 414 ] [ p. 413 ] molle, mais toujours plus ou moins durcie par une matière cornée ou osseuse qui se présente le plus souvent sous forme d’écailles plutôt que de plaques. Bien que de nombreux reptiles passent une grande partie de leur temps dans l’eau, ce sont essentiellement des animaux terrestres, puisqu’ils naissent sur terre et respirent exclusivement par leurs poumons. Chaque animal possède une paire de poumons, mais chez les serpents allongés, le poumon gauche est rudimentaire et presque absent. Chez les serpents et certains lézards, les pattes sont soit totalement absentes, soit réduites à des vestiges enfouis dans la chair. Ces reptiles se déplacent par ondulations, soit sur terre, soit dans l’eau. Lorsque des pattes sont présentes, les aisselles et les doigts sont munis de griffes, une caractéristique très rare chez les amphibiens.
De nombreux reptiles possèdent des paupières, et la plupart des lézards et reptiles fossiles présentent également l’œil pinéal. Les organes de l’odorat et de l’ouïe sont aussi bien développés, mais chez les reptiles, la membrane vibrante du tympan n’est plus externe, comme chez les grenouilles, mais située dans une cavité. À l’exception des tortues, presque tous les reptiles ont des dents ; chez les espèces vivantes, celles-ci sont généralement pointues et souvent recourbées pour maintenir leurs proies. Chez de nombreuses espèces fossiles, cependant, les dents étaient adaptées à la coupe et, plus rarement, à la mastication. Chez les serpents et certains lézards, la langue est fine et bifide, et se projette de façon saccadée. Chez d’autres reptiles, comme les crocodiles, la langue est plate et immobile, fixée au plancher buccal.
Développement des reptiles. — Chez certains lézards et serpents actuels, et plus rarement chez les espèces disparues, les femelles sont vivipares, c’est-à-dire qu’elles donnent naissance à des petits vivants et entièrement formés. La grande majorité des reptiles, cependant, sont ovipares. Leurs œufs ressemblent à ceux des oiseaux, mais sont généralement plus ronds et possèdent une enveloppe parcheminée, dure et poreuse, ou, dans certains cas, une coquille calcaire. Comme leur développement diffère considérablement de celui des poissons et des amphibiens, il convient de les décrire plus en détail.
Comme les œufs de reptiles sont gros et se développent sur la terre ferme, leur structure interne diffère forcément beaucoup de celle des petits œufs d’amphibiens qui éclosent dans l’eau. Chez ces derniers, les embryons possèdent des branchies fonctionnelles adaptées à la vie aquatique, tandis que chez les reptiles, un organe totalement différent s’est développé pour fournir l’oxygène nécessaire au développement de l’embryon. Cet organe embryonnaire en forme de sac, appelé allantoïde, transmet à l’embryon l’oxygène reçu à travers la coquille poreuse et élimine le dioxyde de carbone par le processus inverse ; autrement dit, sa fonction est respiratoire (voir fig. AE, p. 415).
[ p. 415 ]
« Le reptile non éclos ou à naître respire grâce aux courants d’une membrane vasculaire située sous la coquille de l’œuf et qui absorbe l’air sec extérieur. Il est intéressant de noter que cette membrane vasculaire, appelée allantoïde, est représentée chez les amphibiens par une vessie rudimentaire se développant à l’extrémité postérieure du tube digestif. L’apparition de cette membrane prénatale, ou membrane fœtale, et d’une autre membrane – à fonction protectrice – appelée amnios, qui forme une poche d’eau autour du fragile embryon, a impliqué une étape majeure de l’évolution. Cette étape a marqué l’émancipation totale de l’eau et de la respiration branchiale, et les deux membranes fœtales, l’amnios et l’allantoïde, persistent non seulement chez tous les reptiles, mais aussi chez les oiseaux et les mammifères » (J.A. Thomson dans Outlines of Science).
Les œufs de reptiles sont gros et contiennent une grande quantité de vitellus, sur lequel l’embryon se développe. À l’un des pôles de l’œuf se trouve la vésicule germinative fécondée qui se développe en embryon (Fig. A, ci-contre). Au cours de son développement initial, un repli en forme de croissant, composé de deux couches et appelé repli amniotique, se forme à chaque extrémité [ p. 416 ] de l’embryon allongé. Ce repli arqué recouvre l’embryon et finit par se souder pour le recouvrir d’une membrane protectrice. Cette membrane est l’amnios. Entre l’amnios et l’embryon se trouve une cavité peu profonde contenant un liquide amniotique aqueux, qui baigne et protège la surface externe de l’embryon (Fig. B, p. 415). L’amnios étant présent uniquement dans les œufs de reptiles, d’oiseaux et de mammifères, ces groupes sont collectivement appelés les Amniotes, tandis que les poissons et les amphibiens, qui en sont dépourvus, sont appelés les Anamnias, c’est-à-dire « sans amnios ».
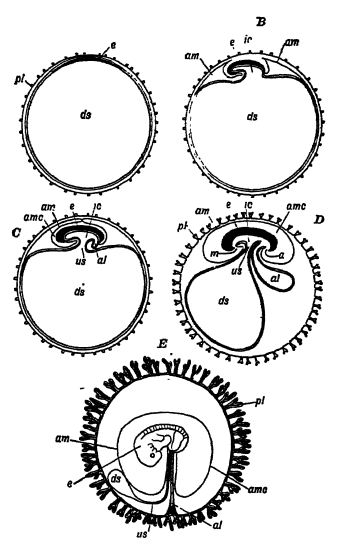
a. gnus; at, allantoïde; am, amnios; amc, cavité amniotique; ds, cavité de la vésicule embryonnaire, devenue plus tard le sac vitellin; e, embryon; e, cavité intestinale; m, bouche; placenta, organe présent uniquement chez les mammifères, formé des parois de l'utérus et de la membrane embryonnaire ; tous deux sont abondamment vascularisés, et bien qu'il n'y ait pas de continuité directe entre les deux circulations sanguines, les nutriments et les gaz respiratoires sont échangés par osmorégulation; u, cordon ombilical. D'après Köniker.
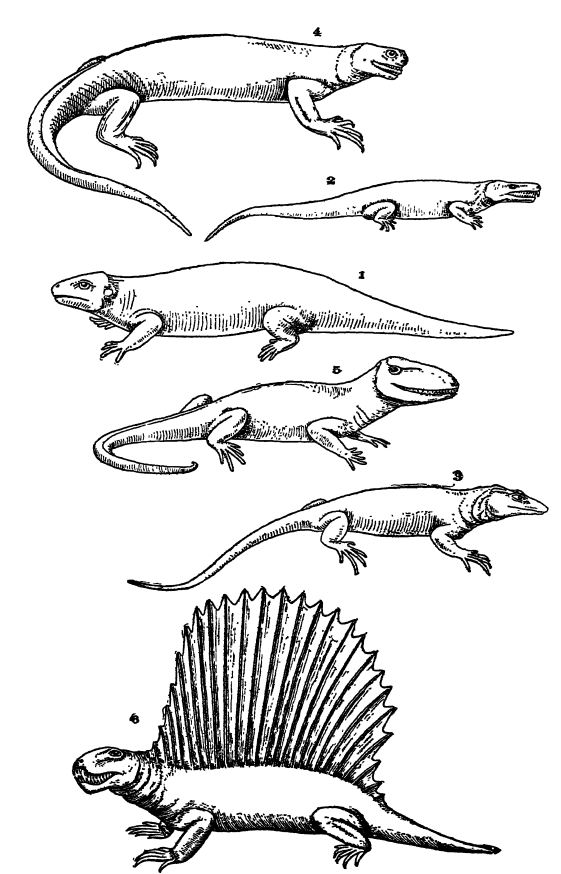
Reptiliens du Paléozoïque. — La plupart des reptiles du Pennsylvanien et du Permien inférieur étaient des animaux trapus, lents et plutôt étalés, se prélassant souvent au soleil. Ils ressemblaient encore à bien des égards aux stégocéphales, leurs congénères, mais présentaient une nette tendance à la réduction de la taille du crâne et à la disparition de la carapace. Chez toutes les espèces, les pieds se terminaient par cinq doigts ou orteils. Rares étaient ceux qui semblaient agiles, et certains, les « lézards-navires » ou « lézards à dos palmé », arboraient une curieuse crête médiane dorsale très haute (Pelycosauria, Pl., p. 413, Fig. 6). « Ces structures », explique Osborn, « pourraient s’être développées par compétition sociale ou raciale et sélection au sein de cette famille de reptiles, plutôt que comme organes offensifs ou défensifs par rapport à d’autres familles de reptiles. » La plupart des formes du Paléozoïque supérieur étaient des animaux à l’allure féroce en raison de leurs grandes dents recourbées et préhensiles. Tous les représentants américains semblent avoir été carnivores, se nourrissant d’insectes, de poissons cuirassés, d’amphibiens cuirassés et d’autres reptiles. D’autres possédaient des dents broyeuses, indiquant un régime alimentaire composé de mollusques et de crustacés.
Osborn affirme que l’environnement qui a transformé les amphibiens primitifs du Paléozoïque supérieur en reptiles du Permien était une région chaude, terrestre et semi-aride, propice au développement d’un système nerveux sensible, de mouvements vifs, d’une armure écailleuse, de membres fins, d’une queue vibrante et de la capture de nourriture à la fois par des dents pointues et recourbées et par les griffes d’une main et d’un pied à cinq doigts. Cette évolution est aussi remarquable et radicale que celle des mammifères qui lui a succédé. Avant la fin du Permien, les reptiles avaient conquis toutes les terres émergées et s’étaient également approprié les eaux terrestres et marines.
Il semble probable que les Reptilia soient apparus encore plus tôt que le Pennsylvanien inférieur, car la dernière partie de cette période a vu apparaître non seulement de véritables reptiles, mais aussi des formes hautement spécialisées. Du Pennsylvanien et du Permien plus ancien du Texas, de l’Oklahoma et du Nouveau-Mexique, Williston et Case nous ont fait connaître de nombreuses espèces différentes [ p. 417 ] d’Amphibia primitifs, auxquelles est associée une société encore plus vaste et complexe de Reptilia primitifs, animaux qui atteignaient une longueur de 2,4 mètres (voir Pl., p. 413).
L’ordre des Cotylosauria regroupe les reptiles à crâne plein, ainsi nommés car leur tête était recouverte de plaques sculptées (Pl., p. 413, Fig. 1, 2). Ils étaient les reptiles les plus primitifs. De constitution trapue et basse, ils se déplaçaient lentement et avec maladresse, mais leur longue queue leur permettait de bien nager. Tous étaient carnivores, bien que certains se nourrissaient également de plantes. Leur aire de répartition s’étend du Pennsylvanien supérieur au Trias supérieur, et on les trouve principalement en Amérique du Nord et en Europe, bien que certains spécimens soient présents en Afrique du Sud.
L’ordre des Théromorphes (qui signifie « bête sauvage »), qui comprend les Pélycosaures, un groupe très spécialisé de formes de vie exclusivement terrestres. Ils étaient actifs et ressemblaient à des dinosaures, avec des queues assez longues, et certains atteignaient une longueur de 2,4 mètres. Les Pélycosaures étaient trapus et possédaient une crête ou nageoire dorsale osseuse plus ou moins haute. Tous étaient farouchement carnivores. Leur aire de répartition s’étendait de la fin du Paléolithique inférieur au Permien supérieur et ils étaient plus abondants en Amérique du Nord, et plus rares en Europe et en Afrique.
L’Afrique du Sud est une autre région où sont conservés de nombreux squelettes exceptionnels d’amphibiens et de reptiles anciens. On les trouve fréquemment dans les épaisses formations de schistes et de grès continentaux du Karoo (jusqu’à 2 900 mètres d’épaisseur), datant du Permien et du Trias. Selon Osborn, c’est au Permien inférieur et moyen que l’on trouve le plus grand nombre de spécimens, époque où la Terre abritait déjà une remarquable diversité de reptiles.
Cet ensemble de reptiles africains comprenait de grandes formes herbivores au corps rond (paréiasaures), ayant des membres massifs et des têtes rondes et solides ; Des reptiles agiles, dotés de grandes orbites et adaptés à la course rapide (dromosaures), des « reptiles terribles » à la tête mammalienne (thériodontes), et de nombreux reptiles prédateurs aux membres fins et aux dents acérées semblables à des canines. Il existait aussi des reptiles carnivores géants (dinocéphales), des animaux très massifs au dos fortement arqué, au front large et proéminent, et aux mâchoires courtes et larges pourvues de dents marginales. Dépassant même ceux-ci en taille, on [ p. 418 ] trouvait les « reptiles aux dents acérées » (anomodontes), dont le crâne mesurait de quelques centimètres à un mètre de long, et dont les mâchoires étaient recouvertes de corne et crochues comme celles des tortues. Enfin, au début du Trias, apparurent les « reptiles à dents de chien » (cynodontes), les plus mammaliens de tous.
Ancêtres des reptiles supérieurs. — Comme mentionné précédemment, les reptiles les plus primitifs sont les Cotylosauria. Lull nous apprend que cet ordre, représenté par Limnoscelis (Fig. 2, p. 413), pourrait bien avoir constitué la souche centrale à partir de laquelle auraient évolué, directement ou indirectement, les lézards, les alligators et les dinosaures. Les cotylosauriens étaient des reptiles des marais, lents, à longue queue et possédant quatre pattes de proportions égales.
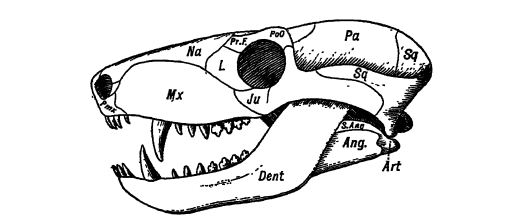
Ancêtres des mammifères. — Au Permien et au Trias d’Afrique et d’Amérique du Nord, on trouve des reptiles carnivores du sous-ordre des Thériodontes. Leur taille varie de petites formes à des spécimens aussi grands qu’un tigre. Ils présentent un grand intérêt car les formes africaines sont considérées comme ayant donné naissance aux mammifères ovipares les plus primitifs, tandis que les formes américaines seraient à l’origine des reptiles supérieurs. Leurs dents étaient différenciées et localisées, comme chez les mammifères, en incisives, canines et molaires (voir Fig., p. 417).
¶ Lecture collatérale
E. C. Case, Les couches rouges permo-carbonifères d’Amérique du Nord et leur faune vertébrée. Carnegie Institution of Washington, Publication n° 207, 1915.
E. C. Case, L’environnement de la vie au Paléozoïque supérieur en Amérique du Nord; une étude paléogéographique. Ibid., Publication n° 283, 1919.
R. L. Moodie, Les amphibiens des couches houillères d’Amérique du Nord. Ibid., Publication n° 238, 1916.
H. F. Osborn, L’origine et l’évolution de la vie, pp. 177-233. New York (Scribner), 1917.
S. W. Williston, Vertébrés du Permien américain. Chicago (University of Chicago Press), 1911.
S. W. Williston, Les reptiles aquatiques du passé et du présent. Chicago (University of Chicago Press), 1914.
S. W. Williston, La phylogénie et la classification des reptiles. Journal of Geology, Vol. 25, 1917, pp. 411-421.
| XXIX. Le charbon et sa présence dans la nature | Page de titre | XXXI. Le Permien et son climat glaciaire |