| XXXVI. Les Dragons du Moyen Âge | Page de titre | XXXVIII. Le Crétacé inférieur et l'apparition des premières plantes à fleurs (angiospermes) |
[ p. 528 ]
¶ Ammonites
Caractéristiques générales. — Les mers du Mésozoïque étaient caractérisées par une abondance de céphalopodes à coquille appelés ammonides (voir Pl., p. 477, Fig. 4-16). Ces magnifiques coquilles enroulées, apparentées aux nautiles, présentaient une grande variété de tailles, de formes, d’ornementations et de types de cloisons (les plaques transversales divisant la cavité de la coquille en chambres). Plus de six mille formes ont été décrites par les paléontologues. La taille moyenne se situait probablement entre 7,5 et 10 cm, bien que leur diamètre puisse atteindre 2,4 m (Pacjiydiscus seppenradensis du Crétacé supérieur d’Allemagne). Si le tube enroulé de ces grandes coquilles était déplié, leur longueur serait comprise entre 6 et 10,5 m. Les ammonides surpassaient donc les nautiles du Paléozoïque autant en taille qu’en diversité spécifique. Il semble qu’ils aient été des animaux plus actifs, de meilleurs nageurs et flotteurs que les nautiles, et qu’ils [ p. 529 ] aient donc moins rampé sur le fond marin. S’ils étaient de bons nageurs, ils devaient également posséder un hyponome fonctionnel (voir Fig., p. 226), comme les nautiles. L’hypothèse qu’ils nageaient bien repose sur la finesse de leurs coquilles, la large répartition de certaines espèces et la faible profondeur, ou l’étroitesse, des cônes enroulés (voir Fig., p. 528). Cette observation s’applique à pratiquement tous les tubes enroulés dans un plan, comme un ressort de montre. En revanche, lorsque les coquilles sont spiralées, plus ou moins déroulées, ou droites, elles sont plus épaisses et l’on suppose que ces animaux rampaient plutôt que de nager (voir Pl., p. 575, Fig. 16). On ne les considère plus comme des formes dégénérées, comme auparavant, mais comme des adaptations nouvelles, voire régressives, à la recherche de nourriture sur les fonds marins. Quelques-unes* étaient peut-être même fixées au fond marin (Nipponites). Avant de poursuivre, l’étudiant est invité à se remémorer les informations concernant les nautiles à la page 225.

Histoire du terme. — Le mot ammonite remonte à 1732, à l’époque où les fossiles étaient considérés comme des minéraux, comme l’indique la terminaison -ite. Ces coquilles furent ainsi nommées en raison d’une ressemblance supposée avec les cornes de bélier, représentées comme l’un des attributs du dieu égyptien Ammon. En Inde, ces fossiles sont commercialisés et utilisés dans certains rites religieux à travers le pays.
Comparaison avec les nautiles. — À bien des égards, les coquilles des ammonites ressemblent à celles du nautile perlé, mais elles en diffèrent radicalement sur un certain nombre de points. Elles sont presque toujours plus ornementées, plus étroites ou moins profondes, et présentent souvent une carène médiane distincte le long du centre des tours externes. De plus, l’ouverture de ces coquilles possède souvent des lobes latéraux (replis), et en général, une carène médiane peut se prolonger en une pointe acérée, le rostre (voir Fig., p. 528). En revanche, le siphon chez les nautiles est situé près du centre des cloisons, tandis que chez les ammonides, il est toujours placé près du cône ou en contact avec celui-ci, sur sa face externe ou ventrale (également appelée ventre). Chez les nautiles, l’ouverture de la coquille n’est jamais fermée par un opercule, mais chez les goniatidés et les ammonides, elle était probablement fermée, chez la plupart des espèces, par une enveloppe lorsque l’animal était au repos. Cette enveloppe, lorsqu’elle est mince et composée de chitine, est appelée anaptyque, mais lorsqu’elle est épaisse et constituée de carbonate de calcium en deux parties articulées, elle est appelée opercule. aptychus. La chambre corporelle est de longueur variable, atteignant parfois une longueur équivalente à un tour et demi de la coquille.
Nature des septa. — Cependant, toutes ces différences, bien qu’importantes, ne sont pas aussi précieuses pour la classification [ p. 530 ] que la nature des septa (voir p. 226). Chez les nautiles, ces cloisons de la coquille cloisonnée sont simples et plus ou moins concaves ; chez les ammonidés, elles ne sont simples que dans leur partie centrale, et chaque septum devient de plus en plus cannelé ou ondulé vers sa jonction avec la coquille externe (Pl., p. 477, Fig. 10 et 14). Lorsque la coquille externe est absente, comme c’est souvent le cas chez ces fossiles, la ligne de suture, ou bord des septa, apparaît toujours comme une ligne ondulée présentant un motif distinct, dont la complexité augmente avec l’âge (voir Fig., p. 366). C’est cette suture complexe, avec ses lobes (orientés vers l’intérieur) et ses selles (courbées vers l’ouverture de la coquille), qui confère à ces fossiles toute leur valeur pour la reconstitution de la chronologie du Mésozoïque. Elle témoigne d’une évolution progressive, déterminée par la séquence des strates et vérifiée par la croissance ou l’ontogenèse de l’individu. L’importance de ces sutures pour la datation a été mise en évidence pour la première fois par Yon Buch en 1830.
On se demande souvent pourquoi ces animaux ont développé un septum aussi complexe. Chez les nautiles, on observe rarement un septum lobé, et leur chambre de vie est non seulement de profondeur variable, mais aussi de largeur importante. Autrement dit, la face ventrale du cône est arrondie et large, et les coquilles sont généralement bien plus profondes que celles des ammonites. Par conséquent, les muscles latéraux qui maintiennent la coquille à l’animal bénéficient non seulement d’une plus grande surface d’appui chez les nautiles, mais, plus important encore, le cône est plus ou moins rond et repose donc plus uniformément sur toutes les parties de l’animal. Chez les ammonites, en revanche, les coquilles sont lenticulaires et se déplacent donc plus facilement sur le côté lorsqu’elles se déplacent dans l’eau. C’est probablement l’étroitesse et la nature compacte des coquilles qui ont conduit à la lobation des septa, par nécessité d’accroître la force de maintien de l’animal sur sa coquille. Dans tous les cas, les cloisons lobées ne s’observent que dans les coquilles lenticulaires des céphalopodes (nautiles, goniatides, ammonides).
Évolution. — Les nautiles à coquille profonde de la fin du Silurien ont donné naissance aux goniatidés, petits et étroits, à cloisons peu lobées. Chez la plupart des goniatidés du Dévonien (Pl., p. 322, Fig. 1-6), la suture des lobes et des selles est abrupte, tandis qu’au Mississippien, la majorité des espèces présentent non seulement un plus grand nombre de lobes et de selles, mais ceux-ci sont presque tous arrondis. On observe ainsi une évolution progressive chez les céphalopodes à coquille étroite, depuis les nautiles véritables jusqu’à divers groupes de goniatidés à coquille plus ou moins étroite, qui ont finalement donné naissance, au Carbonifère, à de nombreuses lignées évolutives parmi les ammonidés primitifs. Ces derniers se sont établis dans les eaux plus chaudes du Permien, en Méditerranée téthysienne (Fig., p. 431). La plus grande variation parmi eux s’est [ p. 531 ] produite à la fin du Permien et au début du Trias. À la fin du Trias, ils ont quasiment disparu, comme expliqué dans le chapitre consacré à cette période. Au Jurassique, une nouvelle évolution rapide s’est produite à partir d’un genre (Phylloceras), et l’apogée de leur développement a été atteint à cette époque. Le déclin des ammoniacés a commencé au Crétacé inférieur, et au Crétacé supérieur, les populations ont peu retrouvé leur vigueur d’antan. Leur extinction complète est survenue à la fin du Crétacé, durant la période critique de la révolution laramienne.
¶ Belemniies, calmars ou poissons-encre
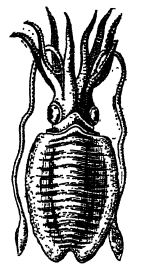
Description générale. — Au Mésozoïque, on observe également l’apparition des bélemnites, ancêtres des seiches (aussi appelées calamars, voir fig. ci-contre). Le mot « bélemnite » vient du grec « belemnite », qui signifie « fléchette ». Ces fossiles étaient autrefois considérés comme les foudres de Thor, dieu du tonnerre (fig. A, p. 532). Hugh Miller rapporte que les gens de la campagne leur attribuaient des vertus miraculeuses pour guérir le bétail ensorcelé. Les bélemnites étaient des céphalopodes très actifs et carnivores, se nourrissant de poissons, de crabes et de mollusques.
Ils possédaient de grands yeux bien développés, étaient dépourvus de coquille externe et, du fait de la présence de seulement deux branchies internes, ont été nommés Dibranchiata. Ils avaient dix bras, probablement huit courts et deux longs et protractiles, comme chez les calmars actuels (Fig. ci-contre). Au Mésozoïque, ces bras étaient souvent munis, sur leur face interne, de crochets chitineux recourbés pour la préhension, et, plus rarement, de ventouses. Par la suite, les ventouses devinrent le type dominant. À l’origine, les bélemnites possédaient un squelette interne massif, mais au cours du Mésozoïque, les calmars émergèrent de cette souche. Ces animaux ont continué, au fil des ères géologiques, à perdre progressivement leur squelette ancestral, bien qu’il en subsiste des vestiges chez toutes les formes vivantes. Chez les calmars, le cerveau est hautement spécialisé et ils représentent l’expression la plus aboutie de la vigilance parmi les invertébrés. C’est pourquoi on les a surnommés les « pirates des profondeurs ».
Sac à encre. — Tous les Dibranchiates possèdent un sac à encre interne contenant de la sépia, un liquide brun-noir qui se mélange facilement à l’eau de mer. En cas de danger, [ p. 532 ] les animaux projettent ce liquide devant eux, puis s’enfuient à reculons, s’éloignant ainsi de ce bouclier d’eau colorée. La sépia utilisée par les artistes est un pigment fabriqué à partir de cette substance. Humboldt, dans son ouvrage Cosmos (1844), affirme que cette encre est si bien conservée chez les spécimens du Jurassique qu’elle permet encore aujourd’hui de reproduire la couleur des animaux. Comme les autres céphalopodes, les Dibranchiates sont dotés d’un siphon par lequel l’eau est propulsée vers l’avant, ce qui leur permet de se propulser vers l’arrière. Ils peuvent également nager grâce à leurs nageoires latérales.
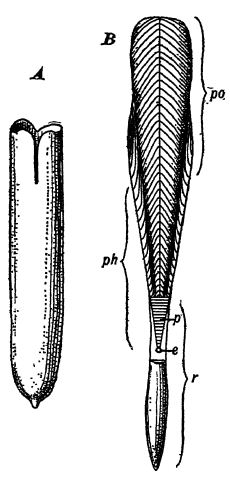
Nature du squelette. — Les bélemnidés sont apparus au début du Trias (Pl., p. 477, Fig. 3) à partir des orthocérides du Paléozoïque (Pl., p. 236, Fig. 19). Ces animaux ont progressivement émergé de leur coquille externe et l’ont enveloppée de leur manteau, formant ainsi un squelette interne – une structure interne témoignant de leur lignée (voir Fig. ci-contre). Parmi les fossiles, la partie généralement conservée est appelée la garde : un corps calcaire solide, plus ou moins fusiforme, pointu à une extrémité et présentant une profonde fossette circulaire à l’autre. Ces gardes peuvent atteindre 60 cm de long et 10 cm d’épaisseur. Dans la fossette des gardes, et s’étendant au-delà, se trouve le phragmacone cloisonné et siphonné, vestige de leur ancêtre orthocéride. Cette structure se prolonge dorsalement par une pièce plus ou moins longue, composée soit de calcaire, soit de chitine, appelée proöstracum (voir Fig. B ci-dessus). Les bélemlides sont très caractéristiques du Mésozoïque, constituent de beaux spécimens stratigraphiques, et on en connaît environ quatre cents espèces, réparties en une soixantaine de genres.
[ p. 533 ]
Les véritables calmars du Mésozoïque ne conservaient que le proostracum et un vestige complètement modifié du phragmacone, formant ensemble une épaisse et large sécrétion de carbonate de calcium, à la texture réticulaire et donc légère : l’os de seiche. Au-dessus du proostracum, et se prolongeant vers la tête, se trouvait la plume, ainsi nommée car elle ressemble à une plume d’oie, faite de chitine. Chez certains calmars ou seiches actuels, il ne reste que la plume. Dans le Jurassique supérieur de Solenhofen, on a trouvé des os de seiche mesurant jusqu’à 60 cm de long, indiquant des animaux dont la longueur corporelle probable était de 1,80 à 2,40 m ; dans les mers actuelles vivent les plus grands spécimens, dont le corps mesure 5,50 m de long, les deux bras protractiles atteignant une longueur de 9 m une fois complètement déployés.
¶ Lecture collatérale
P. Bartsch, Pirates des profondeurs — Histoires de calmars et de pieuvres. Rapport annuel de la Smithsonian Institution, pour 1916, 1917, pp. 347-375.
C. Diener, Lebensweise und Verbreitxmg der Ammoniten. Neues Jahrbuch fiir Mineralogie, etc., 1912, Vol. 2, pp. 67-89.
C. O. Dunbar, Phases d’adaptation des céphalopodes. Dans « Adaptation organique », à paraître aux Presses universitaires de Yale.
R. Ruedemann, Observations sur le mode de vie des céphalopodes primitifs. Bulletin de la Société géologique d’Amérique, Vol. 32. 1921, pp. 315-320.
E. Boese, Les ammonites permo-carbonifères des Glass Mountains, dans l’ouest du Texas, et leur signification stratigraphique - Université du Texas, Bulletin 1762, 1917.
S. S. Buckman, Ammonites de type, vol. 1-4 (autres en préparation). Londres (Wesley), 1909-1923.
A. Hyatt et J. P. Sinna, Les genres de céphalopodes du Trias d’Amérique du Nord. U. S. Geological Survey, Professional Paper 40, 1905.
J. B. Reeside, Jr., Quelques ammonites jurassiques américaines des genres Quenstedticeras, Cardioceras et Amoeboceras, famille des Cardioceratidés. Ibid., Professional Paper 118, 1919.
J. P. Smith, Les faunes d’invertébrés marins du Trias moyen d’Amérique du Nord. Ibid., Professional Paper 83, 1914.
| XXXVI. Les Dragons du Moyen Âge | Page de titre | XXXVIII. Le Crétacé inférieur et l'apparition des premières plantes à fleurs (angiospermes) |