| XXXVII. Ammonites et calmars | Page de titre | XXXIX. Le Crétacé supérieur et la naissance des montagnes Rocheuses |
[ p. 534 ]
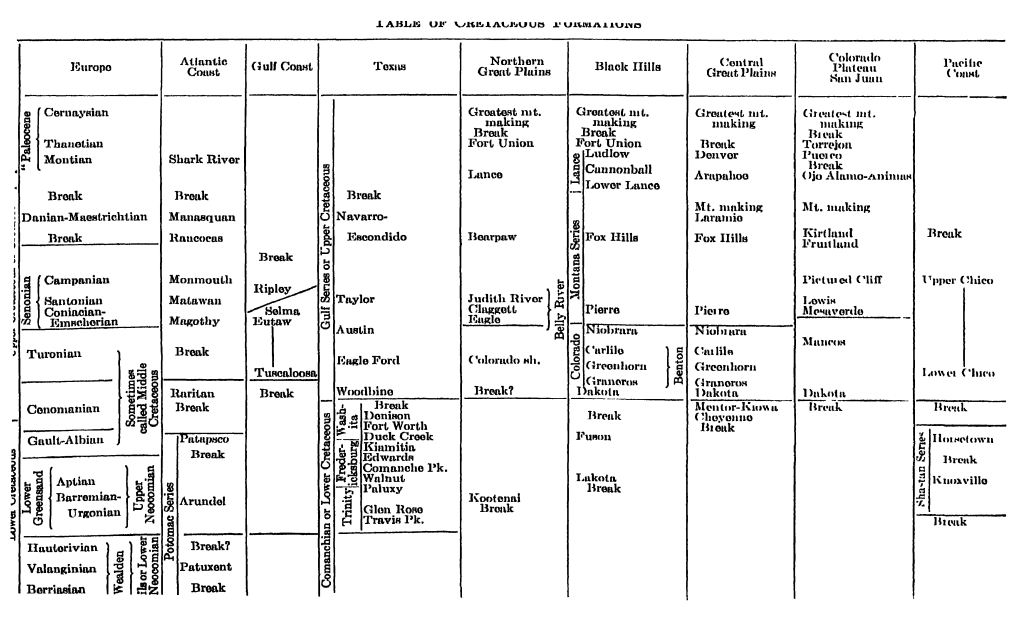
Histoire du terme Crétacé. Au début du XXe siècle, les géologues européens considéraient les roches recouvrant le Jurassique et sous-jacentes au Cénozoïque comme constituant le système crétacé (ainsi nommé du latin creta, craie). Initialement, en Angleterre et en France, ce système ne comprenait que des dépôts de craie du Crétacé supérieur, mais progressivement, des formations d’autres matériaux y furent ajoutées, les fossiles établissant clairement leurs liens. Le Crétacé finit ainsi par englober une telle masse de strates hétérogènes qu’avec l’accroissement des connaissances, il devint nécessaire de séparer les formations en Crétacé inférieur et Crétacé supérieur. Cette classification est aujourd’hui généralement acceptée en Europe. En Angleterre, les caractéristiques physiques de la partie terminale du Crétacé inférieur et de la base du Crétacé supérieur sont si similaires qu’aucune rupture de sédimentation n’est facilement perceptible, tandis que sur le continent, la séparation est généralement aisée. De plus, le Crétacé inférieur correspondait à une transgression marine relativement localisée, avec des dépôts continentaux largement répartis (Wealden, et dans une moindre mesure). Les sédiments du Néocomien (voir tableau, p. 537) sont d’origine marine, tandis que ceux du Crétacé supérieur témoignent probablement de la plus forte expansion océanique de toute l’histoire géologique. Cette expansion, cependant, a débuté tardivement au Crétacé inférieur et a atteint son apogée au Crétacé inférieur (Cénomanien et Sénonien). Ces observations montrent que la Terre n’a connu, au cours du Crétacé, qu’un seul cycle majeur de diastrophisme. Les données physiques permettent donc de classer ses formations en deux périodes, appelées en Amérique Crétacé inférieur et Crétacé supérieur, bien que la limite de séparation diffère de celle en Europe.
Il convient de préciser ici que les stratigraphes américains s’accordent de nouveau sur le fait que leurs strates du Crétacé ne doivent pas être divisées en deux systèmes de roches ou périodes de temps indépendants, mais plutôt en deux divisions d’importance moindre que les périodes elles-mêmes. Cette thèse repose sur le fait que, durant la dernière partie du Crétacé inférieur (tel que défini en Amérique), ou à l’époque de Washita, la mer mexicaine a largement transgressé le continent, s’étendant jusqu’au centre du [ p. 535 ] Kansas et peut-être même jusqu’au sud du Wyoming. Plus au nord, l’équivalent possible de la partie supérieure de Washita a généralement été classé comme Benton ou partie inférieure du Crétacé supérieur marin. Du Mexique jusqu’au centre du Texas, la mer semble avoir continué sans interruption durant le Crétacé supérieur (de Washita = Cénomanien à Eagle Ford ou Woodbine = Turonien inférieur).
Étant donné que la partie supérieure, et peut-être une plus grande partie, de la formation Washita semble dater du Cénomanien européen, qui marque la base de leur Crétacé supérieur, il s’ensuit que la limite entre le Crétacé inférieur et le Crétacé supérieur en Amérique ne coïncide pas avec cette période, sauf dans des régions comme la province des montagnes Rocheuses où l’équivalent supposé de la partie supérieure de la formation Washita se situe à la base du Crétacé supérieur marin. En Europe, cette limite se trouve à la base du Cénomanien, tandis qu’en Amérique, elle se situe au sommet de la formation Washita, comme dans la province texano-mexicaine, mais cette limite est plus ancienne que celle de la région des montagnes Rocheuses. (Voir tableau, p. 537.)
Stanton affirme (1922) que le Comanchien est une bonne série provinciale et que le terme ne devrait s’appliquer qu’à la mer du Mexique. Son application en tant que terme de série au Crétacé inférieur de la côte Pacifique n’est, selon lui, pas justifiée par l’état actuel des connaissances, et la reconnaissance du Comanchien comme système d’application mondiale l’est encore moins.
Crétacé inférieur d’Europe. — En Europe, le Crétacé inférieur se présente en deux phases : (1) une phase septentrionale de faible profondeur, plus froide et plus ou moins saumâtre, se terminant au nord et à l’ouest par des couches continentales (Wealdien et Néocomien) ; et (2) une phase méridionale normale, marine et d’eau plus chaude, largement répandue dans la région méditerranéenne de Téthidan. C’est de cette région, et plus particulièrement de France et de Suisse, que proviennent les divisions stratigraphiques. Celles-ci sont présentées dans le tableau des formations du Crétacé, page 537.
Dépôts de craie. — Bien que la craie ne soit pas le matériau dominant du Crétacé, elle a néanmoins donné son nom au vaste système de roches qui a suivi le Jurassique, en raison de sa couleur blanche caractéristique et de ses affleurements dans les falaises bordant la Manche (bassin anglo-parisien).
On a longtemps cru que la craie était un dépôt océanique, à l’instar des vases à Globigerina (voir p. 72 et p. 115 de la partie I) des abysses actuels. Or, les types de fossiles qu’on y trouve indiquent des mers peu profondes, et ces formations sont souvent accompagnées de sable, tandis que dans les zones adjacentes, les strates équivalentes ne contiennent pas de craie. Au Crétacé inférieur du Texas, les formations de Kiamitia et d’Edwards présentent de la craie associée à des dépôts d’eau peu profonde. Dans le centre du Wyoming, la formation de Niobrara renferme une importante craie blanche, mais dans l’ouest de cet État, les dépôts du même âge sont des schistes sableux [ p. 536 ] ou des grès. Au centre du Kansas, les prétendus dépôts de craie, équivalents de la formation de Niobrara, sont en réalité des boues jaunes très fines, presque dépourvues d’organismes crayeux. Au centre du Texas, la craie d’Austin est également contemporaine de la formation de Niobrara. Dans l’ouest de l’Alabama, la craie de Selma atteint 305 mètres d’épaisseur, mais est d’âge considérablement plus récent, et se transforme latéralement en marnes, argiles et sables. On considère donc aujourd’hui que ces craies sont des accumulations organiques formées principalement par les squelettes calcaires de minuscules plantes et animaux pélagiques, dans des mers épicontinentales ou de plateau continental peu profondes, adjacentes à des terres basses au climat doux.
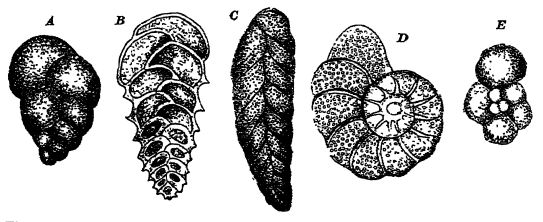
La craie blanche est un calcaire à granulométrie très fine (95 à 98 % de carbonate de calcium dans les régions anglo-parisiennes), composé principalement de tests calcaires entiers ou brisés de foraminifères flottants ou benthiques (voir fig. ci-dessous) et de fragments d’algues calcaires flottantes extrêmement petites (rhabdosphères et coccosphères). On y trouve souvent associés de nombreux invertébrés benthiques à coquille épaisse, vivant en eaux peu profondes, tels que des oursins, des bryozoaires, des brachiopodes et des mollusques. En Europe, les craies contiennent fréquemment aussi des silex de formes et de tailles variées, mais en Amérique, ces altérations secondaires sont rares.
Points importants concernant le Crétacé inférieur américain. — En Amérique du Nord, le Crétacé inférieur présente deux phases de développement marin indépendantes : (1) la formation du géosynclinal mexicain, qui s’étend largement sur le Mexique et vers le nord à travers le Texas jusqu’au Colorado et au Kansas (série comanchienne) ; (2) une phase de développement pacifique connue sous le nom de série de Shastan, appartenant aux géosynclinaux calloman et colombien britannique. On distingue également deux zones de dépôt en eau douce : (3) les strates de Kootenai, largement répandues dans les montagnes Rocheuses canadiennes et riches en gisements de charbon ; et (4) les strates du Potomac, d’étendue limitée, le long de la frontière atlantique des États-Unis. (Voir pl., p. 539.)
[ p. 537 ]
[ p. 538 ]
Un autre élément majeur est le soulèvement, au Jurassique, de la région centrale de la Cordillère, depuis l’Alaska arctique jusqu’au sud du Mexique, et la formation d’un géosynclinal à l’est de celui-ci, qui deviendra au Crétacé supérieur la mer des Montagnes Rocheuses ou mer Coloradique. Ce mouvement crustal n’est qu’un parmi tant d’autres survenus au Crétacé ; un autre est la fragmentation du Gondwana, la plaque continentale reliant l’Amérique du Sud et l’Afrique, accompagnée d’éruptions de lave cataclysmiques au Brésil. Les faits les plus marquants concernant la vie au Crétacé inférieur en Amérique du Nord sont l’apparition des plantes à fleurs et la quasi-absence de dinosaures.
¶ Mers mexicaine ou comanchienne
Série Comanche. — Robert T. Hill a mené, il y a plusieurs années, une étude approfondie des strates du Crétacé inférieur au Texas. En 1887, il leur proposa le nom de Série Comanche, car elles se trouvent sur le territoire des Indiens Comanches. Ses idées ont progressivement prévalu et les roches et leurs fossiles sont aujourd’hui de plus en plus reconnus comme représentant un cycle de sédimentation et de mouvements d’eau : la division Comanche. Cette succession est particulièrement bien connue dans le centre du Texas, où l’on trouve une abondance de fossiles sur une épaisseur de 457 mètres (1 500 pieds). Hill a également divisé le Comanche en trois groupes de formations : la formation inférieure Trinity, la formation moyenne Fredericksburg et la formation supérieure Washita.
Dans le centre du Mexique, où ce regroupement n’est pas encore clairement établi, les géologues adoptent une séparation en deux parties : l’Éocrétacé (inférieur) et le Mésocrétacé (moyen).
Du sud de l’Arkansas au sud-est de l’Arizona, en passant par le centre du Texas, et de là à travers presque tout le Mexique jusqu’à l’isthme de Tehuantepec, on trouve des calcaires et des marnes d’âge comanchien. Ces dépôts témoignent de la plus importante submersion océanique qu’ait connue le Mexique, subissant son apogée au Crétacé inférieur moyen. Ils constituent par ailleurs la plus grande superficie de roches et de faune du Crétacé inférieur en Amérique du Nord (voir pl., p. 539). Aguilera, du Service hydrographique du Mexique, indique que ces dépôts forment la majeure partie des calcaires des montagnes plissées du Mexique et contrastent nettement, à cet égard, avec la nature sableuse des sédiments de Shastan. Leur épaisseur moyenne varie entre 1 000 mètres (Mazapil) et 1 200 mètres (Vera Cruz), et ils sont presque entièrement composés de calcaire (voir fig., p. 540). À Bisbee, en [ p. 540 ] Arizona, l’épaisseur est d’environ [ p. 539 ] 1 430 mètres (4 700 pieds), débutant par des conglomérats, puis passant à des grès, des schistes et des calcaires (198 mètres [650 pieds]), et se terminant par des schistes rouges et des grès veinés. Dans le nord-ouest du Mexique, près d’Hermosillo, on observe une succession de strates très similaire, d’une épaisseur de près de 915 mètres (3 000 pieds), dont les couches supérieures contiennent du charbon, selon Dumble. Au centre du Texas, zone typique des strates comanchiennes, l’épaisseur est de 300 mètres (1 000 pieds), composée de calcaire, de craie, mais principalement de marnes, et s’amincissant à quelques centaines de mètres de sables et de marnes en Arkansas. Dans le sud du Texas et au Mexique, on pense que les mers comanchiennes se prolongent sans interruption jusqu’à celles du Crétacé supérieur.
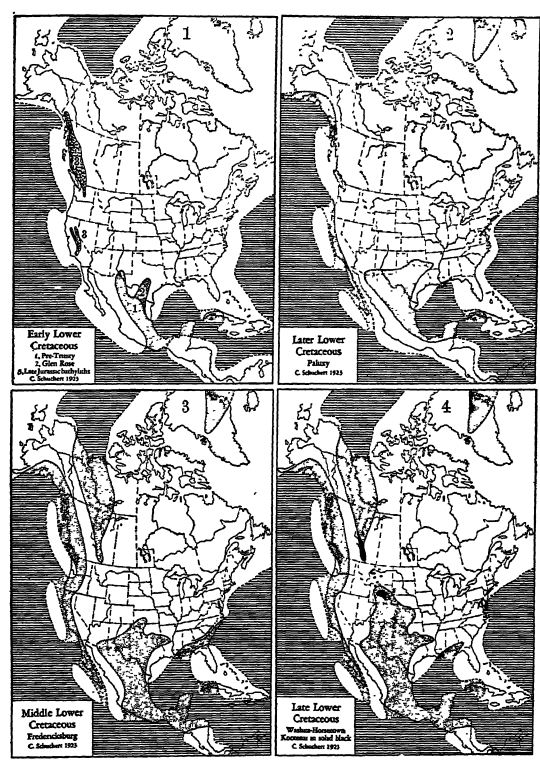
Epeiric and shelf seas dotted; oceans ruled. Fresh-water deposits in solid black, fci Map 1 are shown the more important areas of late Jurassic igneous intrusions, and in Map 2 the beginning of the spread of Mexican waters into the greater Comanchian sea. Note in Map 2 the small areas of Potomac deposits (solid black) along the Atlantic piedmont, and in Maps 3 and 4, the Kome beds of Greenland. Note also the presence of the Cordilleran Intennontane geanticline (see p. 141).
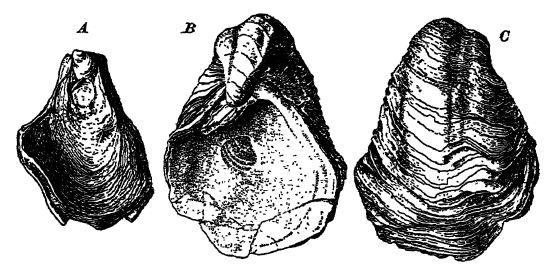
Extension des Grandes Plaines de la mer de Comanchie. — À la fin de la période comanchéenne (Washita), la mer de la région texane s’étendait vers le nord, traversant le Nouveau-Mexique et l’Oklahoma, au sud et à l’ouest des monts Ouachita, jusqu’au centre-est du Kansas et au sud-est du Colorado (voir p. 539, carte 4). Cette extension fut éphémère et ses dépôts sont principalement composés de boues et de marnes, avec des grès à l’ouest et au sud-ouest. Leur épaisseur est généralement inférieure à quelques centaines de pieds, et diminue jusqu’au point de fuite au nord et à l’est. La faune y est toujours limitée, mais comprend des coquilles ressemblant à des huîtres (Gryphoea, fig., p. 541), souvent abondantes. De toute évidence, les eaux étaient peu profondes, turbides et plus ou moins adoucies par les rivières se jetant dans cette vaste baie.

[ p. 541 ]
La flore du début du Crétacé inférieur (Trinity), telle qu’on la trouve au Texas, était encore de caractère jurassique. En revanche, les plantes du sommet du Crétacé inférieur (Washita), découvertes au Kansas, montrent qu’un grand changement s’était opéré dans la flore forestière, car les plantes à fleurs du genre Angiospencil étaient désormais présentes en grand nombre et sous des formes semblables à celles du Crétacé suivant.
La faune d’invertébrés marins était abondante et variée, comprenant environ quatre cents espèces. Elle présentait les caractéristiques typiques des milieux marins au Mexique et dans tout le Texas, s’estompant progressivement vers le nord, au Kansas et au Colorado. Les espèces étaient essentiellement des mollusques, les ammonites représentant environ un tiers des assemblages au Mexique et au Texas, bien que ces dernières soient de moins en moins présentes plus au nord. Parmi les bivalves, les formes les plus caractéristiques étaient les chamidés Requienia et Monopleura, et les rudistes Radiolite, etc. (Fig., p. 542).
La faune du Comanchien, dans son ensemble, est très distincte de celle du Crétacé supérieur, mais la transition entre la faune de Kiowa ou de Denison et celle de Woodbine (Crétacé supérieur) ne présente guère plus de contraste que celle observée au sein des divisions du Benton. Compte tenu des différences de faune dues aux faciès littoraux, d’eaux profondes et récifaux, tous présents dans le Comanchien, cette faune forme une unité ne présentant que les changements progressifs attendus au sein d’une série (Stanton 1922).
Les faunes comanchiennes sont également largement répandues en Amérique centrale, et dans les hautes Andes du nord-ouest et de l’ouest de l’Amérique du Sud, on retrouve des assemblages de ces animaux, bien que fortement modifiés par l’évolution locale. De manière générale, on peut dire que les faunes comanchiennes étaient en grande partie d’origine méditerranéenne (Téthys), ou plutôt que le golfe de Carpentarie était en communication directe avec les mers du Portugal, d’Espagne et du sud de la France.
¶ Mers Pacifique ou Shastan
Série Shastan du géosynclinal californien. Dès 1869, Gabb et Whitney appliquaient le terme Shastan aux roches du Crétacé inférieur de Californie, reconnaissant à juste titre que [ p. 542 ] les strates « contiennent des fossiles semblant représenter des âges allant du Gault au Néocomien inclus (voir tableau, p. 537) ». Cette formation est totalement distincte, tant du point de vue faunique que lithologique, de la série comanchienne. La perturbation nevadienne, à la fin du Jurassique, a considérablement réduit la largeur du géosynclinal californien, le transformant en une dépression étroite mais profonde à l’ouest de la Sierra Nevada et des monts Klamath, à travers toute la Californie occidentale, mais s’étendant largement en Oregon à l’ouest des Blue Mountains. À l’ouest de cette dépression se trouvait une zone frontalière dont font partie les chaînes côtières actuelles. La mer de Shastan (LeConte) s’est étendue dans cette dépression en subsidence. Les volumineux sédiments déversés dans la mer californienne étaient à gros grains et provenaient des rivières descendant des hautes terres. vers l’est.
Les dépôts sont essentiellement composés de schistes sableux avec de minces bandes de grès, des conglomérats locaux et, plus rarement, de minces bancs de calcaire. Leur épaisseur en Californie du Nord semble se situer entre 9 000 et 10 000 pieds, dont environ un tiers date de la période de Knoxville et le reste de celle de Horsetown.
L’épaisseur maximale de la formation de Knoxville, en Californie du Nord, est d’environ 6 100 mètres (20 000 pieds), selon Diller et Stanton. Sur cette épaisseur extraordinaire, environ 4 800 à 5 200 mètres (16 000 à 17 000 pieds) sont généralement pauvres en fossiles, mais on y trouve occasionnellement des couches riches en Aucella piochii, un bivalve d’origine boréale (voir fig., p. 504). Quant à la flore associée, Knowlton la rattache sans hésiter au Jurassique (voir p. 504).
[ p. 543 ]
Géosynclinal colombien britannique. — Des dépôts du Crétacé inférieur sont également connus dans le nord de l’État de Washington et le long des côtes canadiennes et alaskiennes. Ces dépôts sont principalement composés de grès et de schistes sableux, et comprennent généralement des épaisseurs considérables de matériaux volcaniques (de quelques centaines de mètres à 1 020 mètres), de laves, de tufs et de couches de cendres. Dans les îles de la Reine-Charlotte, où ces strates renferment des gisements de charbon, leur profondeur est estimée à 2 900 mètres. Ailleurs, bien que légèrement inférieures, les épaisseurs dépassent rarement 600 mètres, mais par endroits, les strates du Crétacé inférieur ont été entièrement érodées. Enfin, le long de la côte arctique de l’Alaska, on observe également des strates du Crétacé inférieur dépourvues de fossiles, qui atteignent plus de 1 500 mètres d’épaisseur au cap Lisbume.
Les sables et les boues du Crétacé inférieur du Pacifique recouvrent en grande partie, en discordance, des formations plus anciennes, souvent métamorphisées. Cette discordance est généralement marquée, comme dans les monts Klamath et la chaîne côtière, ou bien de type érosif. On observe cependant aussi des contacts en discordance. Les faunes appartiennent au domaine indo-pacifique et se distinguent nettement de celles des mers de Comanche, ce qui indique que les deux provinces étaient complètement séparées l’une de l’autre par une barrière terrestre : la masse continentale occidentale du Mexique. (Voir pl., p. 539.)
Les flores de la haute formation de Knoxville (trente-trois espèces) et de la formation de Horsetown (dix-neuf) étaient de type Crétacé inférieur (basse formation du Potomac), et les plantes à fleurs de type moderne (angiospermes) y sont totalement inconnues. Dans la haute formation de Knoxville (1 220 mètres), la petite faune de mollusques marins était encore d’origine boréale (faune à Aucella crassicollis). Un changement marqué de la faune s’est produit à l’époque de Horsetown, la vie marine prenant alors un aspect méditerranéen (Téthys de l’Inde, également du Japon), une migration qui s’est manifestée en premier lieu en Californie et en Oregon. Cette faune comprenait également environ quatre-vingts espèces de mollusques, dont plus de trente étaient des ammoniacées. Cet ensemble est connu sous le nom de faune indo-pacifique. (Stanton.)
¶ Les phases d’eau douce du Crétacé inférieur
Dépôts continentaux de Kootenai. — Dans le sud de l’Alberta (col d’Oowsnest) et dans le sud-est de la Colombie-Britannique, les formations du Crétacé inférieur présentent une épaisseur considérable le long de l’axe des montagnes Rocheuses et sont d’origine deltaïque. Il s’agit des dépôts de Kootenai, appartenant au géosynclinal des montagnes Rocheuses. Ces dépôts sont principalement composés de grès et de schistes sableux de [ p. 544 ] texture et d’aspect très variés, et contiennent de nombreux gisements de charbon de bonne qualité. L’épaisseur maximale des dépôts de Kootenai est de 1 615 mètres (5 300 pieds), avec vingt-deux gisements de charbon exploitables, couvrant une superficie de près de 7 780 kilomètres carrés (3 000 milles carrés). À l’est, l’épaisseur des dépôts de Kootenai diminue rapidement jusqu’à 61 mètres (200 pieds), avec seulement deux gisements de charbon. Dans la région de Crowsnest, l’épaisseur cumulée du charbon atteint 66 mètres (216 pieds) sur une superficie de 596 kilomètres carrés (230 milles carrés). Dowling estime les réserves de charbon de cette région à près de 8 milliards de tonnes, dont environ 400 milliards de tonnes peuvent être classées comme anthracite.
La formation de Kootenai est également présente aux alentours de Great Falls, dans le Montana, où elle renferme aussi du charbon. Son épaisseur y est toutefois inférieure à 137 mètres en moyenne, atteignant 457 mètres vers l’ouest et diminuant vers le sud. Des vestiges de la partie houillère de la formation de Kootenai subsistent jusqu’au centre du Wyoming. D’autres formations d’âge similaire sont présentes au Wyoming et dans le Dakota du Sud (formations de Lakota et de Fuson), où un millier de souches de cycas silicifiées ont été découvertes (dont une est illustrée à la figure, p. 27). Dans la région des Grandes Plaines, la formation de Kootenai repose directement sur le Jurassique (Morrison) et pourrait inclure des équivalents des formations de Lakota, de Fuson et de Dakota (Lee).
Flore de Kootenai. — Les fossiles de Kootenai sont essentiellement des plantes, une flore de quatre-vingt-six espèces étant connue. Elle présente encore des caractéristiques jurassiques, car aucune plante à fleurs n’y est présente. Les formes présentes sont principalement des fougères (trente-quatre espèces), des cycadées (dix-neuf) et des conifères (vingt-cinq), comme on peut le voir sur la figure, p. 507. Cette flore est très similaire à celle du Bas-Potomac, à la frontière atlantique, vingt formes étant communes aux deux régions. Les botanistes considèrent généralement que la flore de Kootenai n’est pas plus récente que le Crétacé inférieur moyen.
Formation de Blairmore. — Dans la Cordillère canadienne, la formation de Kootenai est directement recouverte par la formation de Blairmore, d’une épaisseur de 518 mètres (1 700 pieds). Les deux formations présentent des caractéristiques lithologiques très similaires. Un conglomérat les sépare, et toutes deux sont d’origine continentale. Viennent ensuite, en net contraste, les schistes marins de Benton. On ne trouve pas de formation de Dakota ici, ni en Alberta, ni dans le nord du Montana. Par conséquent, la formation de Blairmore semble dater du Crétacé inférieur (J.H. Sinclair, 1917).
Dépôts continentaux du Potomac. — Dans le chapitre consacré aux événements du Jurassique, il a été indiqué que les montagnes de l’est de l’Amérique du Nord se sont réduites à un paysage vallonné durant cette période. Il est également vrai que les principaux cours d’eau, les grands fleuves actuels coulant vers l’est, tels que le Potomac, le James, le Roanoke, etc., se sont formés simultanément. À la toute fin du Jurassique, lors de la perturbation du Potomac, il semble que la ligne de rivage de l’océan Atlantique se soit étendue plus à l’ouest (voir p. 503) et que le plateau continental se soit incliné légèrement vers le large. Autrement dit, l’est des Appalaches se fragmentait et s’enfonçait dans les profondeurs de l’océan Atlantique. En conséquence, les cours d’eau sont devenus plus [ p. 545 ] actifs et les sédiments érodés de la région soulevée ont été transportés vers l’est, à un niveau inférieur, et se sont répandus sur le plateau pénéplainisé du Piémont (la pénéplaine de Weverton), composé d’anciennes roches cristallines, pour former la formation du Potomac. Ces dépôts fluviaux sont aujourd’hui visibles depuis les environs de Philadelphie, en passant par Baltimore et Washington, à travers la Virginie, jusqu’en Géorgie et en Alabama (voir pl., p. 539). On en trouve également des affleurements épars dans les îles au large de la Nouvelle-Angleterre.
La formation de Potomac est particulièrement bien développée dans le Maryland, où son épaisseur moyenne se situe entre 180 et 210 mètres. Toutes les strates présentent un léger pendage vers le sud-est, d’environ 12 mètres par kilomètre. Les roches affleurent à l’est des strates triasiques surélevées et, plus à l’est, elles passent progressivement sous les formations du Crétacé et du Cénozoïque. Les dépôts sont principalement composés d’argiles sableuses non consolidées, qui, à la base et à l’ouest, contiennent davantage de sable, voire des conglomérats. Les matériaux y sont généralement arkosiques ou riches en feldspaths fragmentés. On trouve également des quantités importantes de sidérite dans la partie inférieure (formation d’Arundel), exploitée dans le Maryland depuis plus d’un siècle. La présence de ces gisements de fer confère aux formations une couleur rougeâtre sous l’effet de l’altération, et dans le Haut-Potomac, les dépôts présentent une coloration naturellement variée (Patapsco). Le Bas-Potomac débute par des matériaux grossiers très variables d’un endroit à l’autre (formation de Patuxent), et finit par laisser place, dans la région du Maryland, à une zone marécageuse restreinte, comme en témoignent les dépôts sombres et carbonés, parfois même lignitiques (Arundel). De plus, les racines des forêts sont encore à leur point d’origine, et c’est dans ces marécages que sept espèces de dinosaures ont été découvertes.
La formation du Potomac se divise clairement en une série inférieure et une série supérieure par une discordance érosive marquée. De plus, les flores sont très différentes entre les deux parties : le Potomac inférieur (Patuxent-Arundel) conserve des caractéristiques du Jurassique, tandis que le Potomac supérieur (Patapsco) présente une flore caractéristique du Crétacé supérieur, composée de plantes à fleurs ou d’angiospermes. Le Potomac inférieur est entièrement d’origine duntivira et s’est déposé sur les plaines alluviales de la pénéplaine ondulée de Weverton, dont le relief atteignait environ 90 mètres. En revanche, le Potomac supérieur, bien que totalement dépourvu de fossiles marins, pourrait être d’origine estuarienne ; il pourrait représenter la partie d’eau douce ou la partie continentale des différents deltas donnant sur l’océan Atlantique.
Groenland occidental. — Dans le centre-ouest du Groenland, on trouve des grès du Crétacé inférieur moyen alternant avec des schistes argileux sombres présentant localement de très minces et pauvres [ p. 546 ] couches de charbon. Cette formation, appelée formation Home, semble être d’origine hydrothermale et aucun fossile marin n’y a été découvert. L’ensemble de la série atteint environ 213 mètres d’épaisseur et repose sur un substratum vallonné de roches cristallines anciennes (Protérozoïques ?) (voir pl., p. 539). La flore est riche en fougères (quarante espèces), cycadées (onze) et conifères (dix-huit). On y dénombre sept espèces de plantes à fleurs.
¶ Mouvements vers la fin du Crétacé inférieur
Perturbation de la Cordillère centrale (voir fig. ci-dessous). — Le long de la côte Pacifique, du comté de San Luis Obispo en Californie, jusqu’au nord de l’Oregon (chaîne côtière au sens large), on observe des traces de mouvements crustaux au Crétacé inférieur. Anderson indique que la formation de Knoxville est partout dans cette zone traversée et perturbée par des dykes et des masses de serpentine et de péridotites associées. De plus, dans la chaîne côtière du sud de la Californie, où l’on trouve ces intrusions rocheuses, les strates de Horsetown sont également absentes, tandis que la formation de Chico (Crétacé supérieur) repose en discordance sur les formations plus anciennes.
[ p. 547 ]
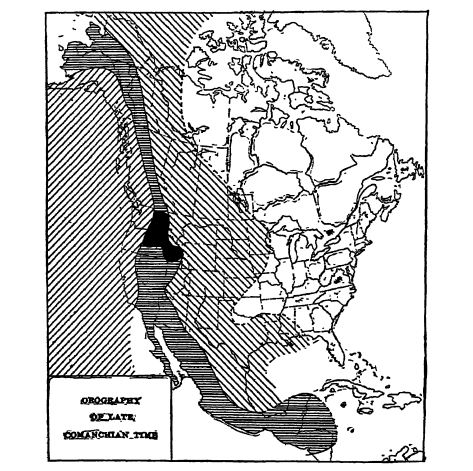
Nous avons vu dans les chapitres précédents que les eaux du Pacifique ont transgressé largement les deux géosynclinaux occidentaux du continent nord-américain au Trias, puis au Jurassique, et que ces inondations se sont produites principalement dans la fosse de Colombie-Britannique. Aux États-Unis, cependant, un soulèvement s’est amorcé dès le Trias moyen, formant apparemment un arc peu prononcé dans l’ouest de l’Utah, l’est du Nevada et l’Idaho, constituant ainsi le plateau du fleuve Columbia (zone noire de la figure, p. 546). Cet arc a persisté jusqu’à la fin du Jurassique. À la fin du Jurassique, les plis de la Sierra Nevada se sont formés au sud-ouest de ce soulèvement, qui a ensuite été intégré à la zone de la perturbation nevadienne. Ce mouvement du Jurassique supérieur a réduit l’ancienne extension des géosynclinaux californien et de Colombie-Britannique.
L’étude de la paléogéographie du Crétacé supérieur révèle rapidement que les conditions d’expansion océanique ont été profondément modifiées vers la fin du Crétacé inférieur. En effet, après cette période, les géosynclinaux du Pacifique étaient étroits et, sur le site actuel des montagnes Rocheuses et loin à l’est, au Canada et aux États-Unis, une nouvelle mer intérieure est apparue : la mer des montagnes Rocheuses ou mer coloradique du Crétacé supérieur, s’étendant du golfe du Mexique à l’océan Arctique (Fig., p. 546). La barrière qui séparait ces eaux était la formation continentale nouvellement bombée à l’ouest des crêtes principales actuelles des montagnes Rocheuses, la Cordillère centrale de Ransome, et cette barrière a continué de s’élever tout au long du Crétacé supérieur. Nous voyons donc ici le début du processus qui a donné naissance à la ceinture cordillère centrale de plateaux élevés s’étendant de l’Arctique AJaska jusqu’en Amérique centrale, et c’est pour cette raison que le mouvement est appelé la perturbation de la Cordillère centrale (perturbation oregonienne de Blackwelder).
Cette perturbation s’est également manifestée au Mexique, car à la fin du Crétacé inférieur, l’ouest du Mexique et l’Amérique centrale jusqu’au Nicaragua étaient surélevés, excluant plus ou moins ici le chevauchement antérieur du golfe du Mexique.
Dislocation du Gondwana (voir fig., pp. 431, 555). — La grande terre équatoriale qui s’étendait de part et d’autre de l’Atlantique, et qui avait si longtemps uni l’Afrique du Nord au Brésil, s’est fragmentée au Crétacé inférieur et a disparu sous les flots au Crétacé supérieur. On en trouve des traces vers la fin du Crétacé inférieur (époque de Gault), lorsque l’Atlantique a commencé à gagner du terrain sur le Brésil et l’Afrique [ p. 648 ] de l’Ouest équatoriale. D’autres chevauchements, plus importants, datent de l’Éocène. On peut donc affirmer que la configuration actuelle de l’océan Atlantique trouve son origine au début du Crétacé. Pour plus de détails, voir le chapitre sur le Jurassique.
¶ Les îles hawaïennes au Crétacé
Dans la première partie de cet ouvrage (p. 116), les îles océaniques sont décrites et comparées aux îles continentales. Il y est démontré que ces îles sont d’origine volcanique et qu’elles sont principalement constituées de laves extrudées provenant du fond océanique. Dès lors, une question se pose : quel est l’âge des îles océaniques ? Les îles Hawaïennes émergent d’une profondeur de 4 572 mètres (15 000 pieds) jusqu’à la surface de l’océan, certaines s’élevant même à plus de 4 267 mètres (14 000 pieds) au-dessus du niveau de la mer. Cette altitude élevée est probablement d’origine très récente (Pléistocène), et l’on considère que l’action des vagues et le ruissellement des eaux de pluie les ramèneront presque au niveau de la mer dans un laps de temps géologiquement court. Nul ne sait combien de fois de tels cycles se sont répétés.
Les îles Hawaïennes ne possèdent pratiquement aucun dépôt marin, hormis ceux des rivages actuels. Par conséquent, pour déterminer leur âge géologique, les naturalistes se basent sur la vie insulaire. Cette vie, principalement composée de plantes, d’insectes et d’oiseaux, est très particulière car elle se compose de nombreuses populations non apparentées, manifestement d’origine diverse, transportées ici par les courants océaniques et atmosphériques. L’origine des plantes a été étudiée par F. B. H. Brown (1921), qui montre qu’elles sont arrivées en trois vagues migratoires. La plus ancienne, aujourd’hui confinée aux hautes terres, semble dater du Crétacé inférieur et supérieur, depuis les hautes terres d’Amérique centrale et du Sud. On peut donc affirmer que les îles Hawaïennes existent certainement depuis le début du Crétacé inférieur et qu’elles ont probablement constitué, à plusieurs reprises, des régions montagneuses. La deuxième vague d’organismes a perduré durant l’Éocène supérieur et l’Oligocène, et ses représentants se trouvent principalement dans les basses terres actuelles des îles. La troisième vague de vie a été introduite par l’homme et n’est présente que depuis quelques milliers d’années tout au plus. Les insectes ne contredisent pas ces conclusions et il est probable que l’on finisse par trouver des escargots terrestres qui corroborent les données relatives aux plantes terrestres.
De tous ces faits, il ressort que les îles océaniques du Pacifique sont pour la plupart d’anciennes structures géologiques, dont certaines remontent certainement au début du Crétacé. Les océans ont probablement toujours abrité des îles et un volcan sous-marin ne peut être détruit car il n’est soumis à aucun agent [ p. 549 ] érosif majeur tel que les vagues ou le sable. Au contraire, même un volcan sous-marin inactif continue de croître en altitude et en circonférence grâce aux organismes qui y vivent. Les Bermudes, dans l’Atlantique, sont une île calcaire et ces dépôts organiques, épais de plusieurs centaines de mètres, recouvrent un volcan éteint, comme le montre un forage en eau profonde qui a atteint les roches ignées sous-jacentes.
¶ La vie au Crétacé inférieur
Flores. — Les flores du Crétacé inférieur se divisent en deux phases de développement : une phase ancienne et une phase récente. Les plus anciennes, comme l’explique Berry, sont celles du Jurassique qui ont persisté jusqu’au Crétacé. Elles se composent de fougères, de cycadées et de conifères (voir fig., pp. 468 et 507). Les joncs, quant à eux, ont considérablement diminué et ne sont plus représentés qu’aujourd’hui. Les fougères du Mésozoïque, plus anciennes, cèdent la place aux fougères modernes. Enfin, à la fin du Crétacé inférieur, les cycadées commencent également à décliner et sont remplacées par les plantes à fleurs, ou angiospermes, dont l’importance croît rapidement. Un tiers de ces flores étaient en effet composées de ce type d’ancêtres. Ces ancêtres des flores modernes annoncent l’avènement des forêts actuelles. En réalité, au moins trois de ces genres existent encore (Sassafras, Populus, Celastrophyllum). On connaît entre trois et quatre cents espèces de plantes du Crétacé inférieur en Amérique, dont environ les deux tiers appartiennent à la forme la plus ancienne. Avant la fin du Crétacé inférieur, cette forêt primitive de feuillus, semblable à celle que nous connaissons aujourd’hui, s’était étendue à l’Alaska, au Groenland et au Portugal, où poussaient des palmiers, des chênes, des érables et des magnolias. Plus tard, au début du Crétacé supérieur, elle a conquis le monde entier.
Les angiospermes, ou plantes modernes, dominent le règne végétal depuis le début du Crétacé supérieur. Leur apparition fut aussi importante dans le monde végétal que celle de l’homme dans le monde animal. Leur seconde modernisation survint au début de l’Éocène, et une troisième dans les climats refroidis du Miocène. On compte aujourd’hui plus de 100 000 espèces vivantes, soit plus que toutes les autres plantes réunies. Les angiospermes sont non seulement les plantes les plus complexes du point de vue structural, mais aussi les plus diversifiées, les plus répandues et les plus adaptables à tous les milieux. Elles vivent sous tous les climats et à toutes les altitudes où les plantes peuvent exister, et forment forêts, prairies et pâturages. « Allant de minuscules organismes aquatiques à des arbres géants de plusieurs centaines de mètres de [ p. 550 ] haut, et ayant une durée de vie allant d’une saison à plusieurs milliers d’années, elles sont les membres les plus impressionnants du règne végétal » (Berry). Au début, les Angiospermes étaient tous arborescents ou ligneux, les herbacés étant d’origine relativement moderne.
Les fruits sont presque exclusivement l’apanage des angiospermes, et leur diversité est presque aussi grande que celle des fleurs. Ces fruits sont d’une valeur inestimable pour de nombreux animaux, et notamment pour l’humanité. Il semble plus qu’une simple coïncidence que les angiospermes soient apparues et se soient répandues à travers le monde avant même le déploiement le plus important et l’évolution la plus significative des mammifères. Berry a raison de penser que la civilisation humaine n’aurait pu se développer sans la présence de ce groupe de plantes.
La structure la plus importante des angiospermes est leur organe reproducteur : la fleur. L’éclat de leurs couleurs et la douceur de leur parfum attirent insectes et oiseaux qui viennent s’y nourrir de leur nectar et, par la même occasion, assurer la fécondation croisée. La structure des fleurs d’angiospermes est remarquablement constante. À l’extérieur se trouve l’enveloppe florale composée du calice et de la corolle, cette dernière étant généralement constituée de pétales. À l’intérieur se trouvent les étamines et, au centre de la fleur, le pistil qui renferme les graines.
Le terme « angiosperme » fait référence à la fermeture des ovaires, une caractéristique unique aux plantes. Cette fermeture protège les graines pendant leur maturation et facilite leur dissémination et leur germination ultérieures. De plus, les graines d’angiospermes contiennent d’importantes réserves nutritives, assurant ainsi le développement de l’embryon. Pour féconder les graines dans les ovaires, les grains de pollen, retenus par le stigmate collant qui recouvre le pistil, doivent le traverser pour pénétrer dans la graine.
Chez les angiospermes, le système ligneux ou vasculaire constitue le tissu conducteur et le squelette de soutien les plus développés de toutes les plantes, leur permettant ainsi de mieux stocker les nutriments. De plus, la grande surface foliaire favorise une production accrue de ces nutriments.
Origine des plantes à fleurs. — L’origine des angiospermes, ou plantes à fleurs, demeure inconnue. Arrivées en Amérique du Nord, au Groenland et au Portugal au Crétacé inférieur, elles y sont apparues « prêtes à l’emploi », sans que l’on puisse encore établir avec certitude leurs origines.
En 1917, la découverte surprenante d’une véritable plante à fleurs, et même d’une espèce distincte (Artocarpidium), dans la partie la plus ancienne du Crétacé inférieur de Nouvelle-Zélande (E. A. N. Arber) fit grand bruit. Ceci indique clairement que ces plantes s’étaient répandues dans le monde entier durant la première moitié du Crétacé, que [ p. 552 ] le genre est plus ancien qu’on ne le [ p. 551 ] pensait généralement, et qu’il pourrait provenir d’un climat froid du Jurassique, voire du Trias.

Sinnott et Bailey (1914-1915) soutiennent que les angiospermes sont apparues dans les flores des hautes terres vivant dans des climats froids, certainement dès le Jurassique et peut-être même au Permien ; et qu’elles proviennent d’un ancêtre conifère palmé plutôt que d’un ancêtre cycadéen penné. Wieland, en revanche, soutient que les angiospermes proviennent d’un ancêtre cycadéen. En 1923, J.H. Hoskins annonça la découverte de bois de monocotylédones dans le Pennsylvanien (Conemaughan) de l’Illinois, mais d’autres botanistes rejettent sa conclusion.
Les angiospermes ligneuses furent les premières plantes à fleurs à apparaître. Parmi ces arbres et arbustes, on compte aujourd’hui environ 4 200 genres, et seulement 2 600 genres d’herbes. Les herbes annuelles, explique Sinnott, descendent de formes ligneuses vivaces, et leur réduction est due soit aux hivers rigoureux, soit aux sécheresses, ce qui a entraîné le développement de rhizomes dans le sol, ou mieux encore, la production annuelle de graines. Les herbes s’adaptent le mieux aux climats froids en survivant sous terre pendant les périodes de basses températures, ou sous forme de graines.
Il est également probable que les fleurs des angiospermes soient apparues indépendamment de la pollinisation par les insectes et que, pendant longtemps, la fructification se soit faite grâce au vent. Il est probable que les fleurs aient été visitées par des insectes au Crétacé inférieur et qu’ils se soient ensuite nourris du pollen, mais l’interdépendance entre les plantes à fleurs et les insectes, si poussée aujourd’hui, est un développement qui a probablement eu lieu au Crétacé (J. J. Lovell 1917).
Dinosaures. — On ne sait presque rien des dinosaures du Crétacé inférieur d’Amérique du Nord, si ce n’est les squelettes fragmentaires de la formation de Potomac, décrits par Lull, mais en Europe, ils sont souvent présents dans les formations de Weald et équivalentes.
Une sépulture des plus intéressantes, abritant des dinosaures bipèdes herbivores du Crétacé inférieur, a été découverte dans les mines de charbon près de Bemissart, en Belgique. Il s’agit d’Iguanodon, un genre connu depuis longtemps grâce à des ossements isolés trouvés dans le Weald, en Europe. Lors de l’extraction du charbon, les mineurs ont mis au jour un dépôt de chenal fluvial recoupant la mine. À plus de 300 mètres de profondeur, ils ont découvert dans ce dépôt vingt-deux squelettes complets et sept incomplets (Fig., p. 551).
Vie marine. — Les ammonites étaient encore nombreuses, quoique moins qu’au Jurassique, mais commençaient à décliner fortement, car très peu de nouvelles populations apparaissaient. Pour plus de détails, voir le chapitre XXXVII. Les bélemnites étaient encore abondantes et florissantes.
La faune invertébrée marine, dans ses autres eaux, ne différait guère de celle du Jurassique ; seuls quelques changements notables méritent d’être soulignés. Les oursins étaient très variés et prolifiques dans les mers plus chaudes, et les oursins à cœur (types irréguliers) atteignirent ici, comme au Crétacé supérieur, leur apogée évolutif (voir fig., p. 347). Parmi les bivalves, les huîtres côtelées (fig., p. 520) et les Gryphaeas (fig., p. 541), ressemblant à des huîtres, étaient très abondantes, notamment dans la Téthys.
[ p. 553 ]
Dans les mers boréales, les aucellidés (Fig., p. 504) étaient encore abondants, tandis que dans les eaux équatoriales d’Europe, du Texas et d’Amérique centrale, apparurent des populations de bivalves remarquablement aberrantes. Chez ces dernières, une valve était cimentée à un objet, la coquille se développant vers le haut en un cône épais, court ou long, torsadé, tandis que la valve de recouvrement était soit torsadée, soit un capuchon simple et épaissi (Fig., p. 542). Il s’agissait des chamidés et des rudistidés, qui persistèrent jusqu’au Crétacé supérieur et donnèrent naissance aux caprinidés et aux rudistidés de plus grande taille, des coquilles qui contribuèrent largement à la construction des récifs.
¶ Lecture collatérale
E. W. Berry, Les flores du Crétacé inférieur du monde. Marjdand Geological Survey, Crétacé inférieur, 1911, pp. 99-151.
E. W. Berry, Paléobotanique ; Esquisse de l’origine et de l’évolution des flores. Rapport annuel de la Smithsonian Institution pour 1915, 1920, pp. 289-407.
Forest B. H. Brown, Origine de la flore hawaïenne. Actes du premier congrès scientifique pan-pacifique, 1921, pp. 131-142.
C. W. Gilmore, La faune de la formation d’Arundel du Maryland. Actes du Musée national des États-Unis, vol. 59, 1921, pp. 581-594.
R. S. Lull, Paléontologie systématique des dépôts du Crétacé inférieur du Maryland : Vertébrés. Maryland Geological Survey, Crétacé inférieur, 1911, pp. 183-211.
Charles Schuchert, Âge des formations américaines de Morrison et est-africaines de Tendaguru. Bulletin de la Société géologique d’Amérique, Vol. 29, 1918, pp. 245-280.
D. H. Scott, L’évolution des plantes. New York (Holt), 1912.
E. W. Sinnott et I. W. Bailey, L’origine et la dispersion des angiospermes herbacées. Annals of Botany, Vol. 28, 1914, pp. 547-600.
E. W. Sinnott et I. W. Bailey, Preuves foliaires concernant l’ascendance et l’environnement climatique précoce des angiospermes. American Journal of Botany, Vol. 1, 1915, pp. 1-22.
E. W. Sinnott et I. W. Bailey, L’évolution des plantes herbacées et son incidence sur certains problèmes de géologie et de climatologie. Journal of Geologj, Vol. 23, 1915, pp. 289-306.
T. W. Stanton, Étude comparative des formations et faunes du Crétacé inférieur des États-Unis. Ibid., Vol. 5, 1897, pp. 579-624.
T. W. Stanton, La formation Morrison et ses relations avec la série Comanche et la formation Dakota. Ibid., Vol. 13, 1905, pp. 657-669.
| XXXVII. Ammonites et calmars | Page de titre | XXXIX. Le Crétacé supérieur et la naissance des montagnes Rocheuses |