[ p. 49 ]
ANCIENS SINGES ANTHROPOÏDES — CLIMAT, FORÊTS ET VIE DU PLIOCÈNE EN EUROPE OCCIDENTALE — TRANSITION VERS LE PLÉISTOCÈNE, OU ÂGE DE L’HOMME — LA PREMIÈRE GLACIATION, SES EFFETS SUR LE CLIMAT, LES FORÊTS ET LA VIE ANIMALE — LA RACE TRINIL PRÉHUMAINE DE JAVA — LES ÉOLITHES OU SILEX PRIMITIFS — LA SECONDE GLACIATION — L’HEIDELBERG, LA PLUS ANCIENNE RACE HUMAINE CONNUE — LA TROISIÈME GLACIATION
Les ancêtres partiellement connus des singes anthropoïdes et les ancêtres inconnus de l’homme sont probablement originaires des forêts et des plaines inondables de l’Asie du Sud et ont commencé très tôt à migrer vers l’ouest, en Afrique du Nord et en Europe occidentale.
Dès l’Oligocène, un précurseur des grands singes (Propliopithecus), très proche des gibbons, apparaît dans le désert bordant le Fayoum, au nord de l’Égypte. Au début du Miocène, de véritables gibbons arboricoles ont pénétré en Europe et ont persisté tout au long du Pliocène sous les formes connues sous les noms de Pliopithecus et Pliohylobates, ce dernier étant un véritable gibbon par ses proportions ; son aire de répartition s’étendait vers le nord jusqu’à l’actuelle Allemagne. Un autre singe arrivé très tôt en Europe est le Dryopithecus ; on le trouve au Miocène dans le sud de la France ; ses dents broyeuses évoquent celles de l’orang-outan, sa mâchoire est profonde et ressemble par certains aspects à celle de l’homme de Piltdown. Un troisième singe (Neopithecus) est présent au Pliocène inférieur, près d’Eppelsheim, en Allemagne. On ne le connaît que par une seule molaire inférieure, qui rappelle la dentition de Dryopithecus et, plus obscurément, celle d’Homo. Dans le Pliocène des collines de Siwalik, en Asie, on trouve Paleopithecus, une forme généralisée que l’on croit apparentée au chimpanzé, au gorille et au gibbon ; les prémolaires supérieures ressemblent à celles de l’homme.
Aucun de ces anthropoïdes fossiles, ni d’Europe ni d’Asie, ne peut être considéré comme l’ancêtre de l’homme, bien que Neopithecus [ p. 50 ] et Dryopithecus aient été placés dans ou près de la lignée des ancêtres humains par des autorités aussi prestigieuses que Branco et Gaudry. Lorsque Dryopithecus fut découvert pour la première fois par Lartet, Gaudry1 le considérait comme le singe le plus humanisé de tous, lui attribuant même une intelligence suffisante pour travailler le silex, mais une connaissance plus approfondie de cet animal a montré que certains anthropoïdes vivants sont plus humanisés que Dryopithecus. Cet animal est étroitement apparenté à la souche ancestrale du chimpanzé, du gorille et de l’orang-outan. La mâchoire, il est vrai, ressemble à celle de l’homme de Piltdown (Eoanthropus), mais les dents qui broient sont beaucoup plus primitives et il y a peu de raisons de penser qu’elles soient ancestrales à un type humain quelconque.[1]

[ p. 51 ]
Parmi ces anthropoïdes fossiles, ainsi que parmi les quatre formes vivantes, nous ne découvrons aucune preuve de parenté directe avec l’homme, mais de très fortes preuves de descendance du même ancêtre. Ces preuves d’ascendance commune, déjà observées chez les races humaines actuelles, deviennent bien plus évidentes chez les races paléolithiques anciennes ; en fait, nous ne pouvons interpréter l’anatomie des hommes de l’Âge de pierre sans examiner les principaux caractères des singes anthropoïdes actuels : le gibbon, l’orang-outan, le chimpanzé et le gorille.

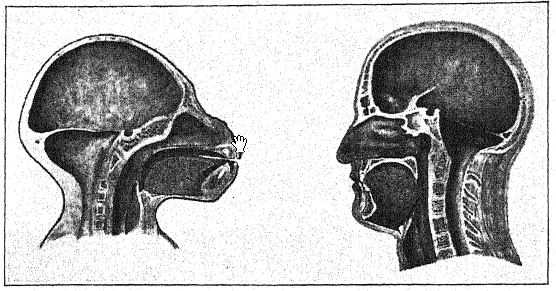
Le gibbon est le plus primitif des singes vivants par son crâne et sa dentition, mais le plus spécialisé par la longueur de ses bras et ses autres adaptations extrêmes à la vie arboricole. Comme chez les autres anthropoïdes, le visage est abrégé, la région narienne est étroite, c’est-à-dire catarrhine, et la boîte crânienne est élargie, mais le sommet du crâne est lisse et le front est dépourvu des crêtes proéminentes au-dessus des orbites ; ainsi, le profil du crâne du [ p. 52 ] gibbon (Fig. i6) est plus humain que celui des autres singes anthropoïdes. Au sol, le gibbon marche droit et peut ainsi utiliser librement ses bras et effectuer des mouvements indépendants de ses doigts. Dans le cerveau, on observe un développement remarquable des centres de la vue, du toucher et de l’ouïe. Ce sont ces caractéristiques du gibbon moderne qui préservent, avec des changements relativement légers, le type de l’ancêtre originel de l’homme, comme l’a noté Elliot Smith.2

Les membres de l’orang-outan sont moins allongés et moins spécialisés pour la vie arboricole que ceux du gibbon, mais davantage que ceux du chimpanzé et du gorille. Le crâne est arrondi et d’une grande hauteur verticale, avec de larges crêtes osseuses au-dessus des orbites et une grande crête médiane au sommet du crâne chez les mâles âgés. La mâchoire inférieure de l’orang-outan est robuste et profonde, et, bien qu’utilisées comme arme de combat, les défenses canines sont beaucoup moins proéminentes que celles du gibbon, du chimpanzé ou du gorille.
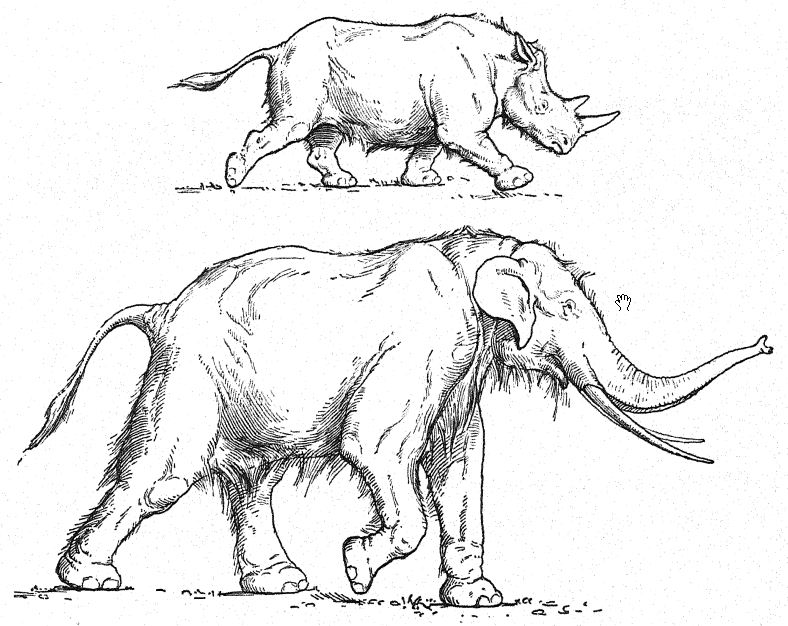
Chez le chimpanzé, on observe des crêtes osseuses très proéminentes au-dessus des yeux, semblables à celles des hommes de Trinil et de Néandertal. De tous les singes anthropoïdes, la mâchoire inférieure du chimpanzé est celle qui ressemble le plus à celle de l’homme de Piltdown. Les rangées de dents prognathes ou saillantes et le menton fuyant évoquent ceux des races de Heidelberg, de Piltdown et de Néandertal. Lorsque le chimpanzé marche (Fig. 18), ses bras descendent sous le niveau des genoux, alors que chez les hommes de races supérieures, ils n’atteignent que la moitié des cuisses.

[ p. 54 ]
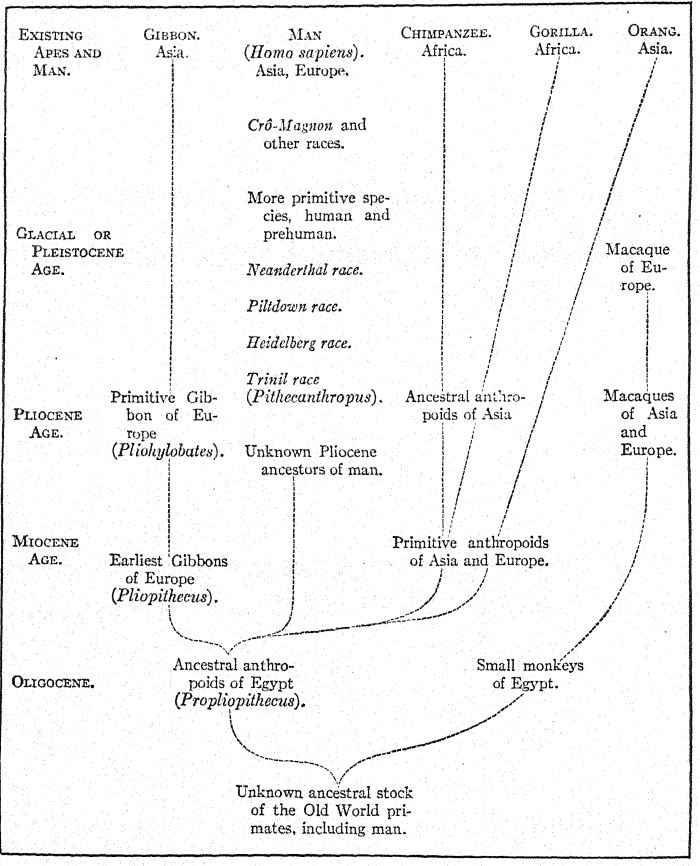
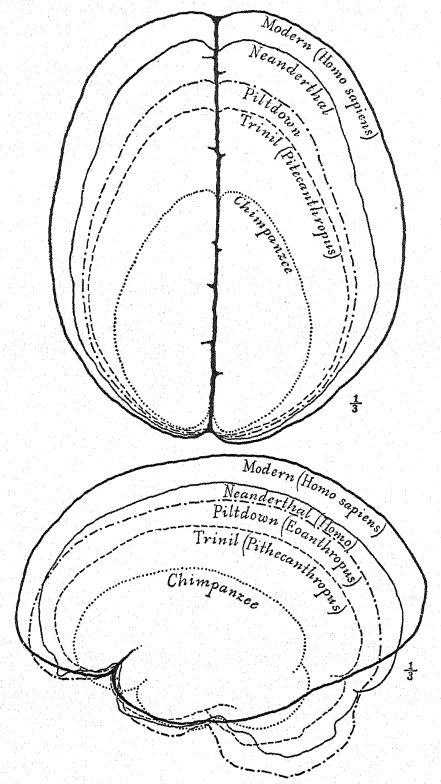
Issu de la lignée ancestrale inconnue des singes anthropoïdes et de l'homme, le gibbon fut le premier à se diviser à l'Oligocène ; l'orang-outan s'est ensuite diversifié dans une direction très différente. Le tronc du chimpanzé et du gorille s'est divisé plus récemment et est plus proche de celui de l'homme. Cinq races humaines primitives ont été découvertes en Europe à l'époque glaciaire ou pléistocène, mais aucune trace d'autres primates, à l'exception des macaques, apparentés à la division inférieure des babouins, n'a été trouvée en Europe au Pléistocène. Modifié d'après Grégoire. (Pour la dernière découverte, voir l'annexe, note VII.)
Français Ainsi, le membre antérieur, bien que beaucoup plus court que celui du gibbon, est relativement plus long que celui de toute race humaine, récente ou ancienne. Nous observons également chez le chimpanzé marcheur (Fig. 18) [ p. 55 ] que la partie supérieure de la jambe, le fémur, est relativement longue, tandis que la partie inférieure, le tibia, est relativement courte. En effet, tant dans le bras que dans la jambe, les os supérieurs sont relativement longs et les os inférieurs sont relativement courts. Ces proportions, qui sont héréditaires de la vie arboricole, contrastent fortement avec celles observées dans les bras et les jambes de la race des hommes de Néandertal, chez qui les membres sont de type terrestre ou marcheur.

On observe également chez le chimpanzé un contraste entre la capacité de préhension du gros orteil, qui est une sorte de pouce, et l’absence de cette capacité dans la main, où le pouce est presque inutile ; chez tous les singes, cette fonction est caractéristique du pied, chez l’homme, de la main seule. Le pouce opposable, avec sa capacité à le plaquer contre chacun des doigts, est le seul caractère absent chez tous les singes anthropoïdes et qui s’est développé très tôt chez les ancêtres de l’homme.
Le crâne du chimpanzé est plus long que celui de l’orang-outan. La caractéristique la plus marquante, vue de dessus, est l’extrême protubérance des orbites, entourées d’une crête osseuse supraorbitaire et ciréoorbitaire. Cette crête est également fortement développée dans le crâne de Néandertal, ainsi que dans celui du Pithécanthrope ou de Trinil, mais, à notre connaissance, elle est totalement absente dans celui de Piltdown. Comme chez l’orang-outan et le gorille, une crête se développe au milieu du sommet du crâne pour l’insertion des puissants muscles des mâchoires, crête totalement absente chez le gibbon et probablement chez tous les véritables ancêtres de l’homme.
Le gorille illustre à l’extrême les spécialisations amorcées chez le chimpanzé, attribuables à une vie mi-arboricole, mi-terrestre, où le crâne et les mâchoires sont utilisés comme de puissants organes de combat. La tête est allongée par la croissance du museau vers l’avant, ce qui entraîne un prognathisme extrême. Les membres et le corps du gorille s’éloignent du type primitif, arboricole et aux membres fins, des singes et sont en partie adaptés à un mode de vie bipède au sol.
En ce qui concerne l’évolution psychique3, Elliot Smith observe que le mode de vie arboricole des premiers ancêtres de l’homme a développé des mouvements rapides, alertes et agiles qui ont stimulé le développement progressif des parties postérieures et latérales du cerveau. L’odorat était bien développé lors d’une vie terrestre antérieure, mais une fois que ces créatures ont quitté la terre et se sont installées dans les arbres, le guidage par le sens olfactif est devenu moins essentiel, car la vie au milieu des branches des arbres est plus propice au développement élevé des sens de la vue, du toucher et de l’ouïe. De plus, elle exige une agilité et une rapidité de mouvement qui nécessitent des centres moteurs efficaces dans le cerveau pour coordonner et contrôler les actions que la vie dans les arbres exige. La spécialisation de la vue éveille la curiosité d’examiner les objets avec plus de minutie et guide les mains vers des mouvements plus précis et plus habiles.
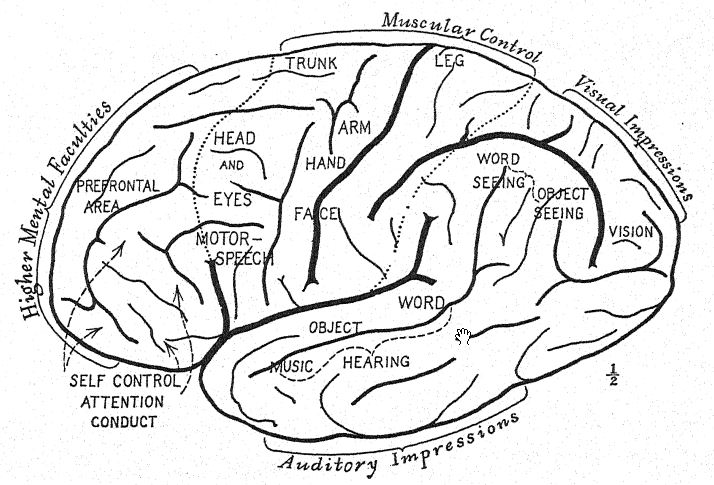
L’anatomie de l’homme est pleine de lointains rappels de cette existence arboricole originelle, ce qui explique également le développement très important et précoce des parties postérieures du cerveau, dans lesquelles se trouvent les différents sens de la vue, du toucher et de l’ouïe.
Le premier passage de la vie arboricole à la vie terrestre est marqué par la faculté de marcher plus ou moins droit sur les membres postérieurs, libérant ainsi les bras ; cette faculté est plus ou moins développée chez tous les singes anthropoïdes ; avec la pratique, ils deviennent des marcheurs experts. La liberté supplémentaire que l’attitude droite confère aux bras, aux mouvements des mains et aux mouvements séparés des doigts est particulièrement notable chez le gibbon. Le développement des facultés de la main réagit sur la croissance et la spécialisation ultérieures du cerveau ; ainsi, le cerveau et l’attitude droite réagissent l’un sur l’autre. Chez le gibbon, on observe une augmentation marquée de la taille des parties du cerveau qui innervent les centres du toucher, de la vision et de l’ouïe.
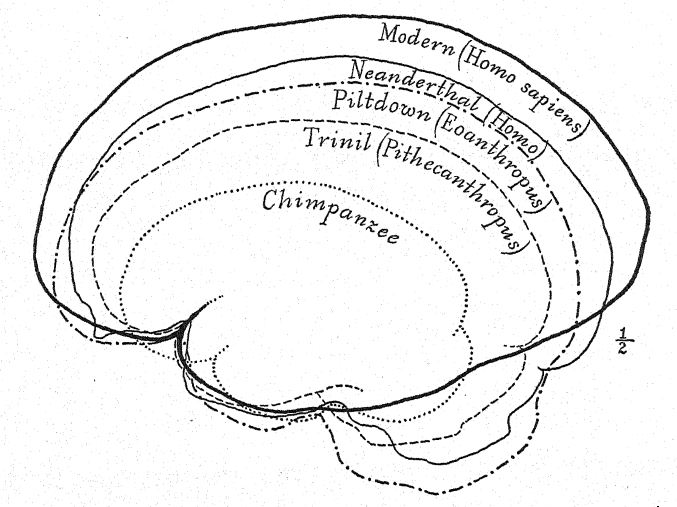
Français La discussion sur la façon dont les ancêtres de l’homme ont été façonnés a principalement porté sur les revendications rivales de quatre lignes d’évolution structurelle : premièrement, l’adoption de l’attitude droite ; deuxièmement, le développement du pouce opposable ; troisièmement, la croissance du cerveau ; et quatrièmement, l’acquisition de la parole. L’argument en faveur de l’attitude droite suggéré par Lamarck, et habilement exposé par Munro4 en 1893, indique que le développement de l’habileté [ p. 59 ] avec les mains et les doigts est à la racine de la suprématie mentale de l’homme. L’argument d’Elliot Smith selon lequel la croissance et la spécialisation constantes du cerveau lui-même ont été le facteur principal de l’ascension progressive des ancêtres de l’homme indique qu’une avancée telle que l’attitude droite a été provoquée parce que le cerveau avait rendu possibles les mouvements habiles des mains.
La véritable conception de l’évolution préhumaine, qui s’est produite au Miocène et au Pliocène, repose plutôt sur le développement simultané de ces quatre facultés spécifiquement humaines. Il ressort des proportions des membres de l’espèce néandertalienne que l’attitude partiellement dressée et la démarche étaient bien plus anciennes à l’époque géologique qu’on ne l’imaginait. Le lien étroit entre l’usage du pouce opposable et le développement des facultés mentales supérieures de l’homme est aujourd’hui confirmé par la découverte que l’une des meilleures méthodes pour développer l’esprit de l’enfant est d’insister sur l’usage constant des mains, car l’action et la réaction entre la main et le cerveau se révèlent être responsables du développement de l’esprit. Une action et une réaction similaires entre le pied et le cerveau ont développé la démarche droite qui a libéré la main de sa fonction de locomotion et de préhension des membres, et par le perfectionnement résultant du mouvement des pouces et des doigts a transformé la main en un organe prêt pour la spécialisation croissante exigée par la fabrication d’outils en silex.
C’est le stade atteint, pensons-nous, à la fin du Pliocène, où l’ancêtre humain émerge de l’âge des mammifères pour entrer dans l’âge de l’homme, période où commence véritablement la préhistoire de l’homme. L’attitude est droite, la main possède un pouce opposable bien développé, les centres cérébraux liés aux sens supérieurs et au contrôle de tous les mouvements des membres, des mains et des doigts sont bien développés. La parole est peut-être encore rudimentaire. Les centres cérébraux antérieurs, responsables de l’emmagasinage de l’expérience et du développement des idées, sont certainement très rudimentaires.
¶ Changement d’environnement en Europe
Considérant que l’origine et le développement de toute créature sont favorisés par une lutte pour l’existence suffisamment intense pour exiger l’exercice complet et fréquent de ses facultés mentales et physiques, il est intéressant de retracer la séquence des événements naturels qui ont préparé l’Europe occidentale à l’arrivée des premières branches de l’espèce humaine. Les forêts et les plantes illustrent de manière encore plus vivante que les animaux les changements de conditions environnementales et de température qui ont marqué l’approche et les diverses vicissitudes de la grande ère glaciaire.
[ p. 61 ]
Les forêts du centre de la France au Pliocène, ainsi que celles de la vallée de l’Arno en Italie du Nord, étaient très semblables à celles du centre des États-Unis aujourd’hui, comprenant des arbres tels que le sassafras, le robinier, le févier, le sumac, le cyprès chauve et le tulipier. Ainsi, les régions qui abritaient la riche faune forestière et préalpine du nord de l’Italie au Pliocène supérieur abondaient en arbres familiers aujourd’hui en Caroline du Nord et du Sud, y compris des espèces typiquement américaines comme le liquidambar (Liquidambar styraciflua), le gommier aigre (Nyssa sylvatica) et le laurier, outre ceux mentionnés ci-dessus. Au sud, le long de la Méditerranée, fleurissaient également des arbres propres à un climat plus tropical : le bambou, le sabal et le palmier nain ; la présence du sabal, qui prospère aujourd’hui dans les forêts tropicales subtropicales du centre de la Floride, est particulièrement intéressante. Le séquoia était également abondant. Vers la fin du Pliocène, les premiers signes de l’arrivée de l’époque glaciaire furent une baisse de la température et, peut-être dans les régions montagneuses les plus élevées, le début des étapes glaciaires.
Les ancêtres des forêts européennes modernes prédominaient dans le centre de la France : le chêne, le hêtre, le peuplier, le saule et le mélèze. Ce sont ces forêts, qui ont survécu aux vicissitudes des périodes glaciaires, qui ont donné naissance aux forêts de l’Europe postglaciaire, tandis que tous les types purement américains ont disparu d’Europe et ne se trouvent plus que dans les régions tempérées des États-Unis.
Nous avons vu que peu de singes anthropoïdes ont été découverts au Pliocène moyen ou supérieur en Europe ; la lignée gibbon-singe disparaît avec les Pliohylobates du Pliocène supérieur. Ces animaux sont cependant rarement retrouvés sous forme fossile, en raison de leur retraite vers les arbres en période d’inondation et de danger, de sorte que nous ne devons pas nécessairement supposer que les anthropoïdes se soient réellement éteints en France. Les primates que l’on trouve au Pliocène supérieur appartiennent aux types inférieurs des singes de l’Ancien Monde, apparentés au langur actuel de l’Inde, au macaque et au babouin. Les preuves, pour autant qu’elles soient disponibles, indiquent que les ancêtres de l’homme évoluaient à cette époque en Asie et non en Europe. Ces preuves seraient néanmoins complètement contredites si l’on pouvait prouver que les éolithes, ou silex primitifs, découverts dans diverses régions d’Europe de l’Oligocène au Pléistocène sont en réalité des artefacts d’origine humaine ou préhumaine.
Les mammifères d’Europe au Pliocène provenaient de migrations très lointaines en provenance d’Amérique du Nord et, plus directement, d’Asie du Sud. L’élément oriental est très fort, incluant des types de rhinocéros aujourd’hui spécifiques à Sumatra et à l’Asie du Sud, de nombreux mastodontes très semblables aux types sud-asiatiques de l’époque, des gazelles et des antilopes, y compris des types apparentés aux élans existants, et des types primitifs de chevaux et de tapirs. Parmi les carnivores d’Europe semblables aux espèces sud-asiatiques figuraient les hysènes, les ours-chiens (Hyaenarctos), les civettes et les pandas (Ailurus) ; on trouvait également les tigres à dents de sabre et de nombreux autres félins. Dans les arbres, on trouvait les singes d’Asie du Sud et d’Afrique du Nord ; et dans les forêts, le cerf axis, aujourd’hui restreint à l’Asie. Mais l’animal afro-asiatique le plus distinctif de cette période se trouvait dans les rivières ; Il s’agit de l’hippopotame, arrivé en Italie au début du Pliocène et qui s’est propagé vers le sud, via le pont terrestre sicilien, jusqu’en Afrique du Nord, et vers l’est, le long des rives méridionales de la mer Noire, jusqu’aux monts Siwalik en Inde. Ainsi, de nombreux ancêtres de ce que nous avons appelé le groupe de mammifères afro-asiatiques du Pléistocène avaient déjà pénétré en Europe au début du Pliocène. Au milieu et à la fin du Pliocène, trois types de mammifères très importants sont arrivés, jouant un rôle majeur au début du Pléistocène. Ce sont :
Les vrais chevaux (Equus stenonis) d’origine nord-américaine lointaine.
Le premier véritable bovin (Leptobos elatus), originaire du sud de l’Asie.
Les véritables éléphants, d’abord Elephas planifrons et plus tard E. meridionalis, mieux connu sous le nom de mammouth du sud, tous deux originaires d’Asie.
[ p. 63 ]
Les forêts et les rives fluviales de la vallée de l’Arno, près de Florence, abritaient tous ces animaux afro-asiatiques au Pliocène supérieur. C’est là qu’ils reçurent leurs noms qui rappellent l’Italie d’aujourd’hui, comme le rhinocéros étrusque (Dicerorhinus etruscus), le macaque florentin (Macacus florentinus), le cheval de Sténo (Equus stenonis), le bœuf étrusque (Leptobos etruscus), premier bœuf à avoir atteint l’Europe.
En Italie et en France, ces mammifères afro-asiatiques se mêlaient aux ancêtres du groupe eurasien plus rustique des forêts et prairies. Parmi ceux-ci, les plus gracieux étaient une variété de cerfs aux bois très élaborés ou à nombreuses branches, d’où leur surnom de cerfs « polycladins ». Dans les forêts erraient les sangliers d’Auvergne (Sus arvernensis), ainsi que les ours d’Auvergne (Ursus arvernensis), les lynx, les renards et les chats sauvages. Dans les rivières nageaient la loutre et le castor, étroitement apparentés aux formes existantes. Parmi les rochers des hautes collines se trouvaient les pikas ou lièvres sans queue (Lagomys), ainsi que les hamsters, les taupes et les musaraignes.
Nombre des animaux les plus caractéristiques des plateaux secs d’Afrique moderne avaient disparu d’Europe avant la fin du Pliocène, notamment des espèces de gazelles, d’antilopes et de chevaux hipparions, tous adaptés aux hautes terres sèches ou aux déserts d’Afrique. La faune pliocène récente, conservée par des auteurs français, témoigne de la fin du Pliocène dans toute l’Europe occidentale, marquée par un climat humide, chaud et tempéré, avec de vastes forêts et rivières entrecoupées de prairies propices à la vie d’une grande variété de cerfs brouteurs ainsi que d’éléphants, de chevaux et de bovins. La flore du Pliocène moyen, telle qu’elle a été découverte à Meximieux, indique une température annuelle moyenne de 16 à 17 °C.
L’une des preuves de la baisse progressive de la température vers la fin du Pliocène en Europe est le retrait vers le sud et la disparition des singes et des anthropoïdes ; le gibbon du Miocène supérieur se trouve aussi loin au nord qu’Eppelsheim, près de Worms, en Allemagne ; au Pliocène inférieur, les singes et les anthropoïdes ne se trouvent que dans les forêts du sud de la France ; au Pliocène supérieur, ils ne sont enregistrés que dans les forêts du nord de l’Italie ; les preuves, dans la mesure où elles vont, indiquent un retrait progressif vers le sud.
Enfin, à la fin du Pliocène, il existait des relations géographiques très étroites, vers l’est, avec les mammifères de l’Inde, via ce qui était alors l’isthme des Dardanelles, et vers le sud, avec les mammifères d’Afrique, via le pont terrestre sicilien. Cela indiquerait que les longues lignes de migration vers l’est et vers l’ouest étaient ouvertes et favorables à l’arrivée en Europe occidentale de nouveaux migrants venus d’Extrême-Orient, y compris peut-être les races humaines les plus primitives. Il n’existe pas la moindre preuve de l’existence de l’homme du Pliocène ou de ses ancêtres en Europe, à l’exception de celles que peuvent fournir les éolithes problématiques ou les silex les plus primitifs.
¶ La première glaciation
Au Pliocène supérieur, des courants marins froids6 venus du nord commencèrent à circuler le long de la côte sud-est de l’Angleterre, avec des signes d’une baisse progressive des températures culminant à une époque où la mer abondait en mollusques arctiques, préservés dans les « Weyboum Crags », une formation géologique le long de la côte du Norfolk. Ce courant arctique annonça la première étape glaciaire.
Il ne semble pas qu’une calotte glaciaire d’une étendue considérable se soit formée en Grande-Bretagne à cette époque, mais c’est à cette époque que la première grande calotte glaciaire s’est formée en Amérique du Nord britannique, à l’ouest de la baie d’Hudson, ce qui a étendu ses calottes glaciaires jusqu’en Iowa et au Nebraska. Dans ce dernier, les forêts domaniales d’épicéas et d’autres conifères témoignent de l’apparition d’une flore tempérée fraîche avant la glaciation. Dans les Alpes suisses, la neige descendait 1 200 mètres sous la limite des neiges actuelles, et en Scandinavie et dans le nord de l’Allemagne se sont formées les premières grandes calottes glaciaires d’où ont jailli les glaciers et les rivières, transportant le « Vieux Diluvium », ou « plus ancien dépôt de glace ». Accompagnant la vague de froid le long de la côte est de l’Angleterre, on remarque, dans les célèbres gisements fossilifères connus sous le nom de « Forest Bed of Cromer », [ p. 65 ] qui surplombent les Weybourn Crags, l’arrivée du sapin (Abies) venu du nord. Ceci est très significatif, car il n’était jusqu’alors connu que dans la région arctique de Crinnell Land, et c’était sa première apparition en Europe centrale. Un autre signe annonciateur des conditions nordiques fut la première apparition du bœuf musqué en Angleterre, attribuée aux dépôts du « Forest Bed ».
Bien que la Grande-Bretagne ait été moins touchée à cette époque que d’autres régions, il ne fait aucun doute que la Première Étape glaciaire s’est étendue à l’échelle de l’Amérique britannique, de la Scandinavie et des Alpes ; dans cette dernière région, elle a été appelée « l’étape de Günz » par Penck et Bruckner. Les dépôts de « drift » ont une épaisseur générale de 98 pieds (30 mètres), mais ils sont en grande partie recouverts et enfouis par ceux de la Deuxième Étape glaciaire, beaucoup plus étendue. La calotte glaciaire scandinave8 occupait non seulement le bassin de la Baltique, mais débordait sur la Scanie (la partie sud de la Suède) et s’étendait jusqu’à Hambourg et Berlin. Dans les Alpes, les glaciers descendaient toutes les grandes vallées montagneuses jusqu’aux basses terres de l’avant-pays, ce qui implique un abaissement de la limite des neiges jusqu’à 4 000 pieds sous son niveau actuel.

¶ La première étape interglaciaire. Les éolithes
Les preuves qu’une vague de froid prolongée a traversé la Grande-Bretagne pendant la première glaciation se manifestent dans ses conséquences, à savoir la modernisation des forêts et la disparition, en Grande-Bretagne et en France, d’un nombre considérable d’animaux, abondants au Pliocène supérieur. Pourtant, la majeure partie des mammifères du Pliocène a survécu, ce qui tend à démontrer que, si des conditions climatiques très froides et de fortes précipitations d’humidité ont pu caractériser les régions entourant immédiatement les champs de glace, le reste de l’Europe occidentale a tout au plus traversé une période de froid prolongée pendant [ p. 67 ] l’apogée de la première étape glaciaire. Cette période a été suivie, pendant le premier interglaciaire, par le retour d’une période légèrement plus chaude que la période actuelle.
Français Ce premier stade interglaciaire est connu sous le nom de Norfolkien, du fait qu’il a été reconnu pour la première fois en Europe dans les dépôts connus sous le nom de « Forest Bed of Cromer », Norfolk, qui contiennent de riches archives non seulement sur les forêts de cette période, mais aussi sur les nobles formes de mammifères qui parcouraient la Grande-Bretagne et la France à l’époque Norfolkienne. Les forêts du Norfolk, à la latitude 52° 40 N., abondaient principalement en arbres encore indigènes à cette région, tels que l’érable, l’orme, le bouleau, le saule, l’aulne, le chêne, le hêtre, le pin et l’épicéa, une flore forestière correspondant étroitement à celle des côtes du Norfolk et du Suffolk en Angleterre à l’heure actuelle, bien que nous trouvions dans cette flore fossile plusieurs espèces exotiques qui lui confèrent un caractère légèrement différent.9 De cette flore arborée, Reid conclut que le climat du sud-est de l’Angleterre était presque le même qu’aujourd’hui, mais légèrement plus chaud.
Nous remarquons surtout qu’un très grand changement s’est produit dans la disparition complète dans ces forêts des arbres qui, à l’époque du Pliocène, étaient communs à l’Europe et à l’Amérique, comme décrit plus haut ; en d’autres termes, la flore de l’Europe s’est fortement appauvrie pendant la première vague de froid.
Dans le sud de la France, comme aujourd’hui, les conditions climatiques interglaciaires étaient plus douces, car on y trouve de nombreuses espèces végétales, aujourd’hui représentées dans le Caucase, en Perse, dans le sud de l’Italie, au Portugal et au Japon. Ainsi, le premier interglaciaire, relativement court, bénéficiait d’une température qui se situe aujourd’hui à environ 4° de latitude plus au sud.
Ce premier étage interglaciaire est également connu sous le nom de Saint-Prestien, car parmi les nombreuses localités de France et d’Italie qui conservent la flore et la faune de l’époque, celle de Saint-Prest, dans le bassin parisien, est la plus célèbre. C’est là qu’en 1863, Desnoyers10 fut le premier à signaler la découverte d’un certain nombre d’ossements de mammifères portant des lignes d’incision, qu’il considérait comme l’œuvre de l’homme. Ces dépôts étaient alors considérés comme datant du Pliocène, ce qui donna immédiatement naissance à une théorie largement répandue [ p. 68 ] de l’apparition de l’homme dès le Pliocène. L’origine humaine des incisions découvertes par Desnoyers a longtemps été sujette à controverse et est aujourd’hui considérée comme très improbable. Des lignes similaires pourraient être d’origine animale, à savoir des marques de griffes ou de dents, ou dues à la pression accidentelle de surfaces coupantes. Cependant, nous ne prétendons pas exprimer une opinion valable quant à la cause de ces incisions. La prétendue confirmation des preuves de Desnoyers concernant l’existence de l’homme du Pliocène serait la découverte par Abbott de plusieurs silex taillés, dont deux in situ, dans la « Forest Bed of Cromer », dans le Norfolk. Bien des années plus tard, dans des gisements similaires à Saint-Prest, furent découverts les prétendus « éolithes » que Rutot a associés à l’Étage Prestien. L’âge des gisements de Saint-Prest est donc une question de la plus haute importance.

[ p. 69 ]
Saint-Prest n’est pas pliocène ; il s’agit plutôt du gisement pléistocène le plus ancien du bassin parisien,11 et ces ossements de mammifères incisés datent probablement du premier interglaciaire. Le lit qui a livré les ossements incisés et la riche série de fossiles est constitué de sables et graviers de rivière grossiers, formant une « haute terrasse », située à 30 mètres au-dessus du niveau actuel de l’Eure. Celle-ci, comme d’autres « hautes terrasses », abrite une faune caractéristique du premier interglaciaire, notamment le mammouth méridional (E. meridionalis) et le cheval de Sténo (E. stenonis). On y trouve aussi d’autres mammifères très caractéristiques du Pléistocène ancien, tels que le rhinocéros étrusque (D. etruscus), l’hippopotame géant du Pléistocène ancien (H. major), le castor géant du Pléistocène ancien (Trogontherium), trois formes du castor commun (Castor) et une du bison (Bison antiquus). La vie mammalienne de Saint-Prest est très semblable à celle du Norfolk, en Angleterre ; à celle de Malbattu dans le centre de la France, dans le Puy-de-Dôme ; de Peyrolles, près de l’embouchure du Rhône, dans le sud de la France ; de Solilhac près du Puy ; de Durfort, dans le Gard ; de Cajarc, dans le Lot-et-Garonne ; et enfin à celle de la vallée de l’Arno, dans le nord de l’Italie.
L’une des raisons pour lesquelles certains auteurs, comme Boule et Depéret, ont situé cette étape au Pliocène supérieur est que les mammifères comprennent de nombreuses formes pliocènes survivantes, telles que les tigres à dents de sabre (Machaerodus), le cerf polycladin aux bois élaborés (C. sedgwicki), le rhinocéros étrusque et le cheval de Sténon primitif. Or, nous avons récemment découvert qu’à l’exception du cerf polycladin, ces mammifères ont certainement survécu en Europe jusqu’au deuxième interglaciaire, et il existerait des preuves que certains ont même persisté jusqu’au troisième interglaciaire.
Français C’est donc l’extinction ou la disparition d’Europe de nombreux animaux très abondants même à la fin du Pliocène qui marque cette faune comme datant du début du Pléistocène. On ne trouve plus de singes anthropoïdes ; en effet, il n’y a aucune preuve de la survie d’aucun des primates, à l’exception des macaques, qui survivent dans les Pyrénées jusqu’à la fin du Pléistocène ; le tapir a entièrement disparu des forêts d’Europe ; mais le départ le plus significatif [ p. 70 ] est celui du mastodonte, dont on pense qu’il a persisté en Afrique du Nord et qui a certainement survécu en Amérique jusqu’à la fin du Pléistocène. La vie animale de l’Europe occidentale, comme la vie végétale, a perdu une partie de son aspect pliocène tout en conservant une autre partie, tant dans sa faune mammalienne que dans sa flore forestière.
Français Le milieu vivant dans son ensemble prend en outre un aspect nouveau avec l’arrivée, principalement du nord, d’animaux et de plantes plus robustes qui évoluaient depuis très longtemps dans les forêts et prairies tempérées d’Eurasie, au nord-est et au nord-ouest. De cette région eurasiatique sont venus le cerf élaphe (Cervus elaphus), ainsi que le cerf géant (Megaceros), et des marais du nord, l’élan à tête large (Alces latifrons). La présence de membres de la famille des cervidés (Cervidae) en grand nombre et représentant de nombreuses lignées différentes est l’un des traits les plus distinctifs du premier interglaciaire. Outre les nouvelles formes nordiques mentionnées ci-dessus, il y avait le chevreuil (Capreolus), qui survit encore en Europe, mais il n’existe plus aucune trace du [ p. 71 ] magnifique cerf axis (Axis), qui s’est maintenant retiré en Asie du Sud. Le cerf « polycladine », observé pour la première fois dans la vallée de l’Arno, est représenté au premier interglaciaire par le cerf de Sedgwick (C. sedgwicki), dans le Norfolk, et par l’espèce C. dicranius du nord de l’Italie, « où l’on trouve également le cerf des Carnutes » (C. carnutorum).

Nous observons que les types brouteurs, forestiers et riverains prédominent. Parmi les carnivores forestiers figuraient le carcajou, la loutre, deux espèces d’ours, le loup, le renard et la martre ; un autre habitant de la forêt était un sanglier, apparenté au Sus scrofa européen.
Ainsi, au tout début du Pléistocène, les forêts d’Europe regorgeaient d’une vie sauvage très semblable à celle de la préhistoire, à laquelle se mêlait l’élément oriental : les grands éléphants, les rhinocéros et les hippopotames, reliant l’Europe à l’Extrême-Orient. Parmi ces migrants orientaux du début du Pléistocène, on compte deux nouveaux arrivants : le bétail sauvage primitif (Bos primigenius) et le premier bison (Bison priscus).
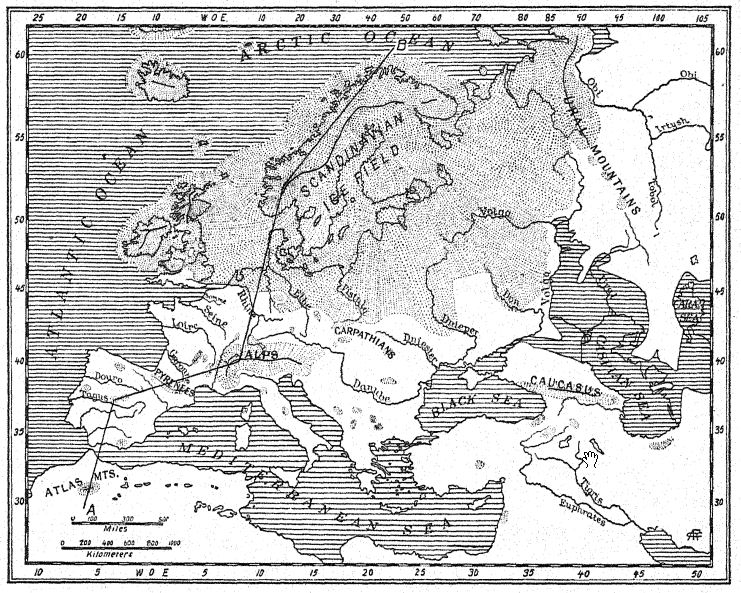
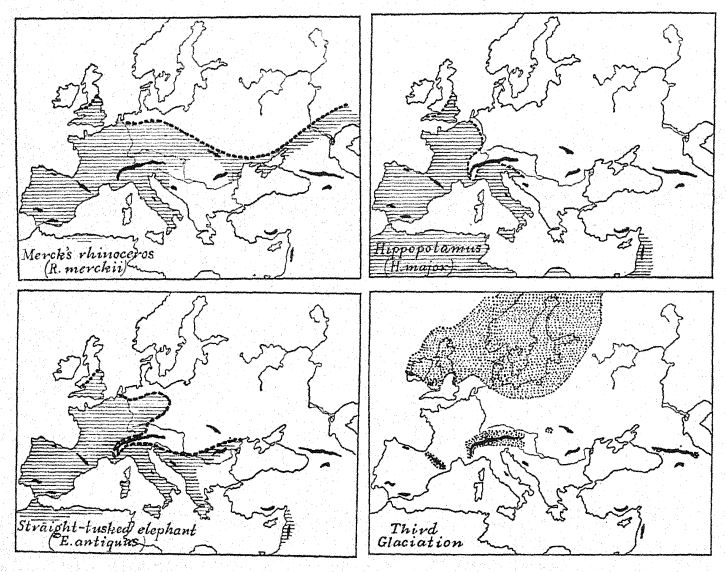
La carte théorique de l’Europe occidentale durant le premier interglaciaire (fig. 12, également fig. 56) permet de comprendre ces migrations depuis le nord-est et l’Orient. Comme l’indiquent les chenaux fluviaux submergés découverts sur l’ancien plateau continental, le littoral s’étendait loin à l’ouest jusqu’aux limites du plateau continental, aujourd’hui profondément enfoui sous l’océan ; les îles Britanniques étaient séparées de la France non par la mer, mais par une large vallée, tandis que le Rhin, avec la Tamise comme affluent occidental, coulait vers le nord sur une vaste plaine inondable, qui constitue le fond actuel du bassin de la mer du Nord. Il n’est pas improbable que les riches gisements de vie mammifère du « lit forestier de Cromer », dans le Norfolk, aient été charriés par les affluents de cet ancien Rhin.
Dans tous les grands fleuves de cette Europe occidentale élargie, on trouvait des hippopotames, et le long des rives et dans les forêts, le rhinocéros étrusque broutait. Parmi les formes broutant et vivant dans les prairies du Norfolk, en Grande-Bretagne, figuraient des espèces de bovins sauvages (Bos, Leptohos), ainsi que deux espèces de chevaux, dont une forme plus légère ressemblant au cheval de Sténo (E. stenonis cocchi) du Val d’Arno et un type plus lourd appartenant probablement aux forêts. L’éléphant géant de cette période est le mammouth méridional (E. meridionalis trogontherii), un descendant quelque peu spécialisé du mammouth méridional pliocène de la vallée de l’Arno ; cet animal est surtout connu grâce à un superbe spécimen découvert à Durfort (Fig. 42) et conservé au Muséum de Paris. On dit qu’il atteignait une hauteur de plus de 3,6 mètres, contre 3,4 mètres, la taille des plus grands éléphants d’Afrique actuels. Il est probable que tous ces migrants sud-asiatiques en Europe étaient partiellement ou entièrement couverts de poils, pour s’adapter au climat chaud et tempéré des étés et aux hivers frais. Plus au sud, dans le climat encore plus doux de l’Italie, l’arrivée d’une autre grande espèce, connue sous le nom d’« éléphant ancien » ou « éléphant à défenses droites » (E. antiquus), est attestée. Cet animal n’avait pas encore atteint la France ou la Grande-Bretagne.
Les grands machasrodontes, ou tigres à dents de sabre, s’attaquaient aux membres sans défense de cette faune hétérogène et parcouraient l’Europe, l’Afrique du Nord et l’Asie. Il ne semble pas que les véritables lions (Felis leo) aient encore pénétré l’Europe.
L’intercommunication des formes de vie sur une vaste zone s’étendant sur 9 600 kilomètres, de la vallée de la Tamise à l’ouest jusqu’à l’Inde au sud-est, est attestée par la présence d’au moins six espèces d’éléphants et de rhinocéros, semblables ou apparentées. À quatre mille cinq cents kilomètres au sud-est des contreforts de l’Himalaya, des troupeaux de mammifères similaires, mais à un stade d’évolution plus précoce, parcouraient l’île de Java, qui faisait alors partie du continent asiatique.
¶ La race Trinil de Java
L’intérêt humain pour cette grande faune réside dans le fait que les voies migratoires ouvertes par ces grandes races animales ont peut-être aussi ouvert la voie aux premières races humaines. Ainsi, la découverte de la race Trinil dans le centre de Java, au sein d’une faune étroitement apparentée à celle des contreforts de l’Himalaya et plus éloignée à celle du sud de l’Europe, a un rapport plus direct avec notre sujet qu’il n’y paraît à première vue.
Sur la rivière Bengawan, dans le centre de Java, un chirurgien militaire néerlandais, Eugen Dubois, effectuait des fouilles à la recherche de fossiles dans l’espoir de découvrir des restes préhumains. En 1891, il découvrit près de Trinil un dépôt de nombreux ossements de mammifères, dont une molaire supérieure unique qu’il considérait comme celle d’une nouvelle espèce de singe. En dégageant soigneusement la roche, le sommet d’un crâne apparut à environ un mètre de distance de la dent. Des fouilles plus poussées, à la fin de la saison des pluies, mirent au jour une deuxième molaire et un fémur gauche à environ 15 mètres de l’endroit où le crâne avait été trouvé, enchâssés et fossilisés de la même manière. Ces fragments épars furent décrits par Dubois13 en 1894 comme le type de Pithecanthropus erectus,[2] terme désignant l’[ p. 74 ] homme-singe debout. Français Le terme spécifique erectus fait référence au fémur, dont l’auteur observe : « Nous devons donc conclure que le fémur du Pithécanthrope a été conçu pour les mêmes fonctions mécaniques que celui de l’homme. Les deux articulations et l’axe mécanique correspondent si exactement aux mêmes parties chez l’homme que la loi de parfaite harmonie entre la forme et la fonction d’un os nécessitera la conclusion que cette créature fossile avait la même posture verticale que l’homme et marchait également sur deux jambes. … De là, il s’ensuit nécessairement que la créature avait le libre usage des membres supérieurs - maintenant superflus pour la marche - et que ces derniers étaient sans doute déjà bien avancés dans cette ligne de différenciation qui les a développés chez l’humanité en outils et organes du toucher. . . . De l’étude du fémur et du crâne, il résulte avec certitude que ce fossile ne peut pas être classé comme simien. . . . Et, comme pour le crâne, de même pour le fémur, les différences qui séparent le Pithécanthrope de l’homme sont moindres que celles qui le distinguent de l’anthropoïde le plus élevé. … Bien que très avancée dans le cours de la différenciation, cette forme pléistocène n’avait pas encore atteint le type humain [ p. 75 ]. Pithecanthropus erectus est la forme de transition entre l’homme et les anthropoïdes dont les lois de l’évolution nous enseignent l’existence. Il est l’ancêtre de l’homme.


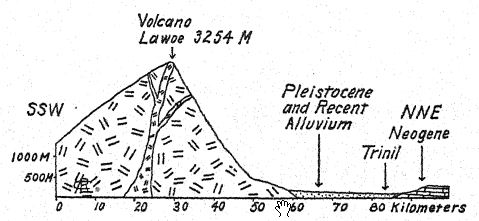
Ainsi l’auteur a placé Pithecanthropus dans une nouvelle famille, de l’ordre des Primates, qu’il a nommé les Pithecanthropidae.
L’âge géologique des ossements mentionnés est primordial. Les restes de Pithécanthrope reposaient dans un dépôt d’environ un mètre d’épaisseur, constitué de grès tufacés meubles, grossiers, sous lequel se trouvait une strate d’argile dure bleu-gris, et sous cette brèche marine. Au-dessus de la strate de Pithécanthrope se trouvait la strate « Kendeng », un grès tufacé multicouche, d’environ 15 mètres d’épaisseur. Cette série géologique était considérée par Dubois et d’autres comme datant de la fin du Tertiaire ou du Pliocène ; Pithécanthrope fut ainsi surnommé « l’homme-singe du Pliocène » tant attendu. Français Les recherches ultérieures menées par des géologues experts ont eu tendance à situer l’âge au début du Pléistocène17. Selon Elbert18, les strates de Kendeng recouvrant la couche de Pithecanthropus correspondent à une période pluviale précoce de basse température et, en termes de temps, à l’[ p. 76 ] Âge glaciaire de l’Europe. Car même à Java, on peut distinguer trois divisions de la période du Pléistocène, y compris la première période de basse température à laquelle la couche de Pithecanthropus est attribuée.
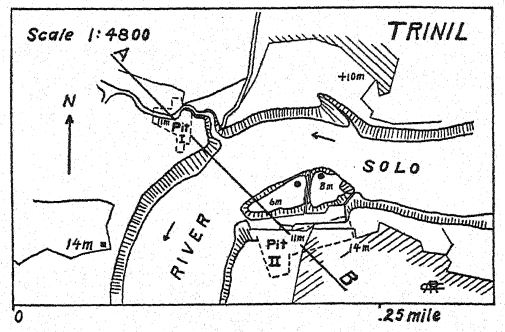
Français Les mammifères fossiles contenus dans la couche Pithecanthropus ont également été étudiés en profondeur,19 et ils tendent à confirmer la référence originale au Pliocène supérieur. Ils fournissent une faune très riche semblable à celle des collines de Siwalik en Inde, comprenant le porc-épic, le pangolin, plusieurs félins, la hyène et la loutre. Parmi les primates, outre Pithecanthropus, on trouve un macaque. Parmi les plus grands ongulés, on trouve deux espèces de rhinocéros apparentées aux formes indiennes existantes : le tapir, le sanglier, l’hippopotame, le cerf axis et le cerf rusa, le buffle indien et le bétail sauvage. Il est à noter que trois espèces d’éléphants du Pliocène tardif, toutes connues sous le nom de Stegodon, et en particulier l’espèce Stegodon ganeza, sont présentes, ainsi que Elephas hysudricus, une espèce apparentée à E. antiquus, ou l’éléphant aux défenses droites, qui est arrivé en Europe au début du Pléistocène. Des fossiles du même animal ont été découverts dans les contreforts de l’Himalaya, en Inde, à environ 4 000 kilomètres au nord-ouest. Les gisements indiens sont considérés comme datant du Pliocène supérieur,20, car il s’agit de la fin de la vie des Siwaliks supérieurs de l’Inde.
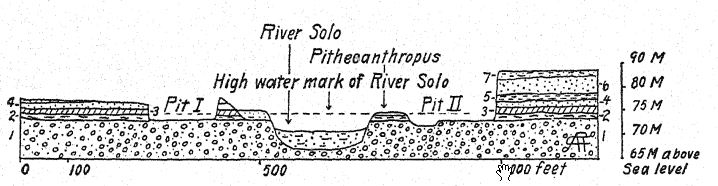
Récent : 7. Lavoir fluvial, argile bleu-noir.
Pléistocène : 6. Grès clair, semblable au tuf.
5. Tuf gris avec boules d'argile, coquillages d'eau douce.
4. Grès blanc strié ressemblant au tuf.
3. Argile bleu-noir avec restes végétaux.
2. Strate osseuse. Pithecanthropus.
I. Conglomérat de lahar.
[ p. 77 ]

Java faisait alors certainement partie du continent asiatique, et des troupeaux similaires de grands mammifères parcouraient librement les plaines, depuis les contreforts de l’Himalaya jusqu’aux rives de l’ancien fleuve Trinil, tandis que des singes similaires peuplaient les forêts. À cette époque, l’orang-outan a peut-être pénétré les forêts de Bornéo, qui sont aujourd’hui son territoire ; c’est le seul singe découvert jusqu’à présent dans le Pliocène supérieur de l’Inde. On peut donc s’attendre à la découverte, à tout moment, en Inde, d’une race semblable au Pithécanthrope.
L’âge géologique de la race Trinil doit donc être considéré comme se situant à la fin du Pliocène ou au début du Pléistocène.
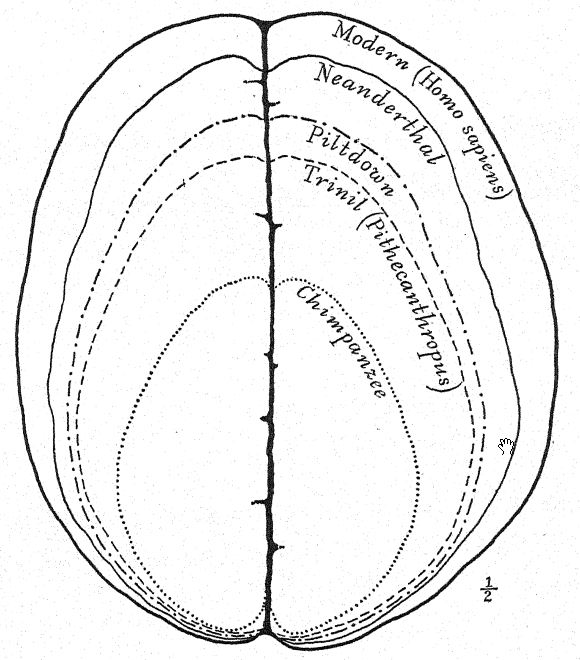
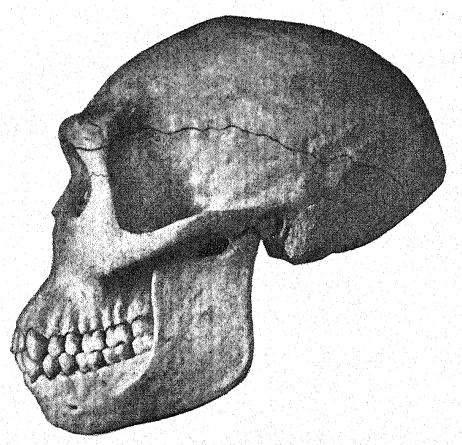
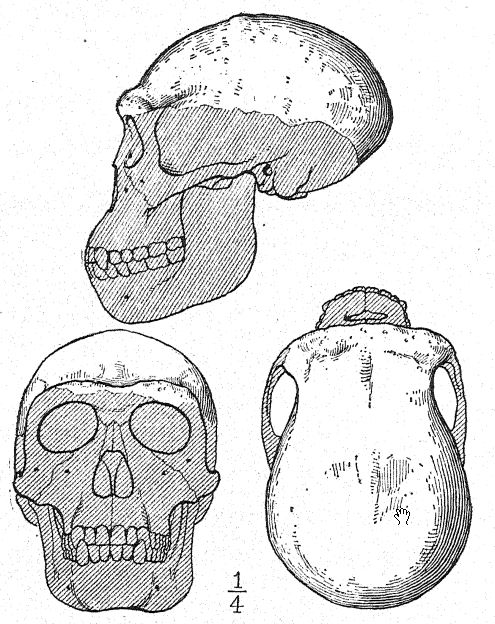
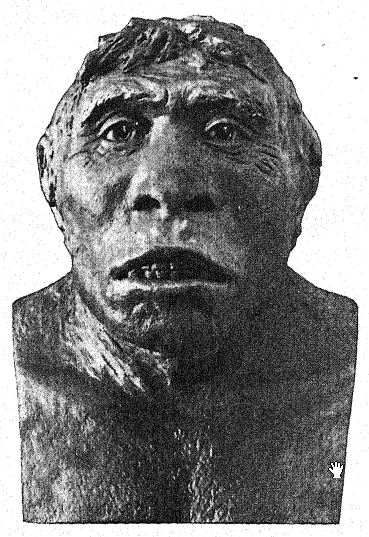
Français Cette grande découverte de Dubois a suscité une discussion large et passionnée, à laquelle ont pris part les plus grands anatomistes et paléontologues du monde. Certains considéraient le crâne comme celui d’un gibbon géant, d’autres comme préhumain, et d’autres encore comme une forme de transition. Nous pouvons cependant nous forger notre propre opinion à partir d’une compréhension plus complète des spécimens eux-mêmes, en gardant toujours à l’esprit qu’il s’agit de savoir si le fémur et le crâne appartiennent au même individu ou même à la même race. Tout d’abord, nous sommes frappés par la ressemblance marquée que le sommet du crâne présente, tant en le regardant de côté que de dessus, avec celui de la race néandertalienne. Cela justifie pleinement l’opinion de l’anatomiste Schwalbe21 selon laquelle le crâne du Pithécanthrope est plus proche de celui de l’homme de Néandertal que de celui même du [ p. 78 ] plus grand des singes anthropoïdes. Tel que mesuré par Schwalbe, l’indice de la hauteur du crâne (Kalottenhöheindex) peut être comparé à d’autres comme suit :
| La race humaine la plus basse | 52 pour cent. |
| Homme de Néandertal | 40,4 pour cent. |
| Pithecanthropus, ou race Trinil | 34,2 pour cent. |
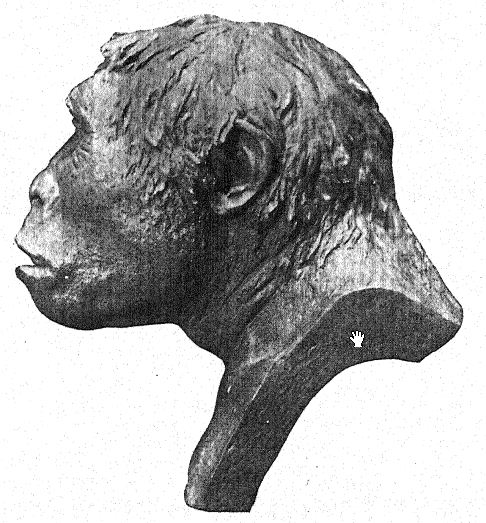
Français Cela concorde avec l’estimation de la capacité cérébrale[3] de 855 cm³ (Dubois) contre 1 230 cm³, la plus petite capacité cérébrale trouvée chez un membre de la race néandertalienne. Deuxièmement, vu de dessus, nous sommes frappés par la grande longueur de la calotte crânienne par rapport à sa largeur, l’indice céphalique ou rapport largeur/longueur étant de 73,4 % (Schwalbe) contre 73,9 % dans le crâne de type néandertalien ; cette dolichocéphalie concorde avec le fait que toutes les premières races humaines découvertes jusqu’à présent ont une tête longue, bien que selon Schwalbe22 tous les anthropoïdes aient une tête large. Il s’agit d’une distinction très importante. La troisième caractéristique est la proéminence et la largeur des crêtes sourcilières osseuses au-dessus des orbites, qui sont presque aussi grandes que chez le chimpanzé et dépassent largement celles [ p. 79 ] de la race néandertalienne et de l’Australien moderne. Le profil de la tête de Trinil restauré par McGregor (Fig. 38) présente cette crête osseuse proéminente et le front bas et fuyant. Selon l’opinion la plus récente de Schwalbe23, le Pithécanthrope peut être considéré comme l’un des ancêtres directs de l’homme de Néandertal et même de l’espèce humaine la plus élevée, l’Homo sapiens. Il considère également que lorsque la mâchoire inférieure de la race Trinil sera connue, elle se révélera très similaire à celle de l’homme de Heidelberg, la conclusion finale étant que le Pithécanthrope et l’homme de Heidelberg, qui lui est presque apparenté, peuvent être considérés comme les ancêtres communs de la race néandertalienne, d’une part, et des races supérieures, d’autre part. Il existe cependant des raisons d’exclure le Pithécanthrope de la lignée ancestrale directe des races supérieures de l’homme.

Ce stade préhumain a néanmoins une très grande importance dans l’histoire du développement de l’homme. À notre avis, c’est précisément ce stade que nous devrions théoriquement retrouver à l’aube du Pléistocène. Buchner adopte un point de vue similaire, en présentant dans un admirable diagramme (Fig. 117) le résultat de sa comparaison de douze caractères différents dans les crânes de Pithécanthropes, de Néandertaliens, d’Australiens et de Tasmaniens. L’un des principaux objectifs des recherches de Buchner était une comparaison très détaillée du crâne de Trinil avec celui de la race tasmanienne, aujourd’hui éteinte, qui, comme nous le constatons sur le diagramme, occupe une position à peine supérieure à celle de la race néandertalienne Spy.
Si le fémur appartient au crâne, les Trinils étaient une race de grande taille, atteignant une hauteur de 5 pieds 7 pouces contre 5 pieds 3 pouces chez les Néandertaliens. Le fémur (Fig. 122) présente une très légère courbure par rapport à celui des singes ou des lémuriens, et à cet égard est plus humain ; il est remarquablement allongé (455 mm), surpassant celui des Néandertaliens ; le tibia était probablement proportionnellement court. Les deux dents broyeuses supérieures préservées sont beaucoup plus humaines que celles du gibbon, mais elles ne ressemblent pas suffisamment à celles de l’homme pour confirmer positivement la théorie préhumaine. Dubois observe :25 « L’appartenance de la dent à une forme hominidé n’est plus à démontrer. Outre sa taille et la plus grande rugosité de sa surface broyeuse, elle diffère de la dent broyeuse humaine en ce que la cuspide la moins développée du Pithécanthrope est la cuspide postérieure, près de la joue, alors que chez l’homme, il s’agit généralement de la cuspide postérieure, près de la langue. La simplification de la couronne et de la racine de la dent broyeuse de Trinil est tout aussi importante que chez l’homme. »
Divers efforts ont été déployés pour compléter les rares et épars matériaux collectés par Dubois. L’expédition Selenka de 1907-1908 a rapporté une molaire inférieure gauche humaine, seul résultat d’une recherche expresse de restes de Pithécanthrope. [ p. 82 ] Dubois posséderait également un fragment de mâchoire inférieure d’apparence primitive provenant de la chaîne connue sous le nom de Kendeng Hills, à la base sud de laquelle se trouve le village de Trinil.
Il nous reste à considérer le stade d’évolution psychique atteint par la race Trinil, et cela dépend naturellement de l’attitude droite et du peu que l’on sait de la taille et des proportions du cerveau.
Adopter une attitude droite ne se résume pas à apprendre à équilibrer le corps sur les membres postérieurs.26 Cela implique des changements à l’intérieur du corps, la perte de la queue, la libération des bras et l’établissement du diaphragme comme principal muscle de la respiration. Le fémur du Pithécanthrope ressemble tellement à celui de l’homme qu’il étaye la théorie selon laquelle la position droite aurait été adoptée par les ancêtres de l’homme dès l’Oligocène. Il semblerait que le Pithécanthrope ait eu libre usage de ses bras et il est possible que le contrôle du pouce et des doigts ait été cultivé, peut-être lors de la fabrication d’outils primitifs en bois et en pierre.
La découverte de l’utilisation du bois comme outil et comme arme a probablement précédé celle de l’utilisation de la pierre.
Elliot Smith décrit ainsi ce stade de développement27 : « … L’affranchissement des mains de la progression a fait reposer toute la responsabilité sur les jambes, qui sont devenues plus efficaces pour leur fonction de soutien, une fois qu’elles ont perdu leur pouvoir préhensile et se sont allongées et spécialisées pour une progression rapide. Ainsi, l’attitude droite est devenue stéréotypée et figée, les membres se sont spécialisés, et ces simiens dressés ont émergé de leurs forêts ancestrales en sociétés, armés de bâtons et de pierres et des rudiments de tous les pouvoirs qui leur ont finalement permis de conquérir le monde. L’exposition accrue au danger que ces esprits plus aventureux ont rencontré une fois qu’ils ont émergé à découvert, et les luttes constantes que ces premières créatures semi-humaines ont dû mener face à des ennemis bien définis, tout autant qu’aux forces de la nature, ont fourni les facteurs qui ont rapidement éliminé ceux qui n’étaient pas adaptés aux nouvelles conditions et, par sélection naturelle, ont fait des survivants de véritables hommes. »
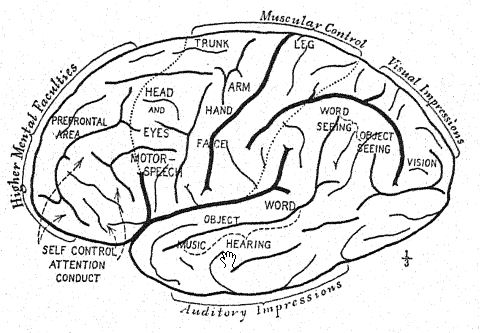
Le front peu développé du Pithécanthrope et la petite zone frontale du cerveau indiquent que la race Trinil avait une capacité limitée à tirer profit de l’expérience et des traditions accumulées, car c’est dans cette zone préfrontale du cerveau que se situent les pouvoirs d’attention et de contrôle des activités de toutes les autres parties du cerveau. Dans le cerveau du singe, les aires sensorielles du toucher, du goût et de la vision prédominent, et elles sont bien développées chez le Pithécanthrope. Français La zone centrale du cerveau, qui est le magasin des souvenirs des actions et des sentiments qui leur sont associés, est également bien développée, mais la zone préfrontale, qui est le siège de la faculté de profiter de l’expérience ou de rappeler les conséquences des réponses antérieures à l’expérience, est développée à un degré très limité.28 Ainsi, alors que le cerveau du Pithécanthrope est estimé à 855-900 cm3, comparé aux 600 cm3 du plus grand cerveau simien et aux 930 cm3 du plus petit cerveau enregistré dans la race humaine, cela indique un stade d’intelligence très bas.
¶ Absence de paléolithes et présence d’éolithes en Europe occidentale
En revenant aux conditions du premier interglaciaire en Europe, nous observons que les cours d’eau traversaient les mêmes vallées qu’aujourd’hui, mais qu’au début des périodes glaciaires, les chenaux étaient beaucoup plus larges et s’élevaient de 100 à 150 pieds au-dessus du niveau actuel relativement étroit des rivières. Les vastes crues de la glaciation suivante ont comblé ces vallées, mais certaines des « hautes terrasses » étaient déjà formées. Il est extrêmement important de noter que des silex pré-chelléens ou de véritables paléolithes n’ont jamais été découverts dans les sables ou les graviers de ces « hautes terrasses ».
Les éolithes découverts sur ce niveau de « haute terrasse » à Saint-Prest appartiennent à la culture Prestien de Rutot29, qui considère cette station comme datant du Pliocène supérieur. Ceux-ci, comme d’autres silex éolithiques supposés, sont très bruts, mais, malgré leur rudimentarité, ils présentent généralement une partie façonnée comme pour être saisie par la main, tandis que l’autre partie est tranchante ou pointue comme pour être taillée. Il est généralement admis que ces silex sont pour la plupart de formes accidentelles, et il n’existe que peu ou pas de preuves de leur façonnage par la main de l’homme. À ce sujet, Boule30 observe : « Quant aux éolithes, j’ai combattu cette théorie non seulement parce qu’elle me paraît improbable, mais parce qu’une longue expérience géologique m’a montré qu’il est souvent impossible de distinguer les pierres fendues, taillées ou retouchées par des agents purement physiques de certains produits de fabrication rudimentaire. »
D’un autre côté, il est intéressant à ce stade de citer les mots de MacCurdy31 : « Mon opinion, fondée sur mon expérience personnelle, … est que l’existence d’une industrie primitive, antérieure à ce qui est communément accepté comme le Paléolithique, a été établie. Cette industrie remonte au Miocène supérieur et se poursuit tout au long du Tertiaire supérieur jusqu’au Quaternaire inférieur inclus. Les caractères distinctifs de l’industrie restent peu modifiés tout au long de la période, la subdivision de la période en époques étant basée sur la stratigraphie [étages géologiques] et non sur les caractères industriels. Les besoins en outils étant très simples et l’approvisionnement en matériaux sous forme d’éclats naturels et de fragments de silex étant très abondant, les pouvoirs inventifs de la population sont restés en sommeil pendant des siècles. Le marteau et le couteau étaient les outils d’origine. Tous deux étaient récupérés prêts à l’emploi. Un éclat naturel tranchant servait pour l’un, et un nodule ou un fragment pour l’autre. » Lorsque le tranchant de l’éclat s’émousse à l’usage, la pièce est soit jetée, soit retouchée pour une utilisation ultérieure. Si le marteau ou l’éclat ne permet pas une prise en main confortable, les pointes ou les bords gênants sont retirés ou réduits par ébréchure. Le stock d’outils s’accroît lentement, parallèlement à la croissance des besoins. À mesure que ceux-ci se multiplient et que l’approvisionnement naturel en matière première diminue, celui-ci est complété par la fabrication d’éclats artificiels. Lorsque la leçon d’associer des formes d’outils précises à des usages précis est apprise, des types particuliers apparaissent, notamment l’outil amygdaloïde et le poignard. Puis vient la transition de l’Éolithique au Paléolithique, une étape qui a été si minutieusement étudiée par Rutot.
Il n’est pas improbable que la race Trinil ait été à un stade de culture éolithique ; il est hautement probable que les races préhumaines de cet âge géologique très reculé aient utilisé plus d’une arme de bois et de pierre.
¶ La Grande Seconde Glaciation (Fig. 25, p. 65)
Au début du Pléistocène, une élévation générale de l’Europe méridionale unissait les îles de la Méditerranée à l’Europe au nord et à l’Afrique au sud, formant de larges liaisons terrestres entre les deux continents, offrant des voies de migration vers le nord et le sud. À cette époque, certains mammifères typiquement africains, comme l’éléphant à défenses droites et le lion, se dirigeaient probablement vers le nord ; la Sicile acquit alors sa riche faune d’éléphants et d’hippopotames, et l’île de Malte fut reliée au continent, tout comme les îles orientales de Chypre et de Crète. Il semble probable que la connexion entre l’Italie continentale et Malte ait été renouvelée à plusieurs reprises.
L’approche de la deuxième glaciation est indiquée le long de la côte sud-est de la Grande-Bretagne par l’affaissement des terres et la montée de la mer, accompagnés d’un nouveau courant arctique, apportant avec eux une invasion de coquillages arctiques qui se sont déposés dans une couche de lits marins directement au-dessus de ceux qui contiennent les [ p. 87 ] [ p. 88 ] [ p. 89 ] riche faune et flore chaudes du « lit forestier de Cromer », Norfolk.®^ Il semble également probable qu’un courant froid du nord ait balayé les côtes occidentales de l’Europe, et Geikie estime qu’une baisse de température d’au moins 20° Fahr. s’est produite, un changement aussi important que celui que l’on observe actuellement en passant du sud de l’Angleterre au Cap Nord.

La deuxième glaciation fut de loin la plus importante d’Europe et d’Amérique. Dans la région des Pyrénées, qui, bien plus tard lors du troisième interglaciaire, devint un territoire privilégié de l’homme paléolithique, on trouvait des glaciers d’une immense étendue. La comparaison avec les conditions actuelles le confirme. Le plus grand des glaciers actuels des Pyrénées ne mesure que 3 kilomètres de long et se termine à 2 200 mètres d’altitude. Durant la plus grande glaciation, la neige semble être descendue de 1 300 mètres sous son niveau actuel. Des Pyrénées, par la vallée de Gallego jusqu’en Espagne, coulait un glacier de 61 kilomètres de long, tandis qu’au nord, le glacier de la vallée de la Garonne coulait sur 72 kilomètres jusqu’à un point proche de Montrejeau. Même dans sa partie inférieure, ce glacier avait plus d’un kilomètre d’épaisseur. À l’est se trouvait un glacier de 61 kilomètres de long, remplissant la vallée de l’Ariège et recouvrant les sites de grandes cavernes paléolithiques comme celle de Niaux ; il est probable que la formation de cette caverne ait commencé à cette époque. Que ces glaciers soient tous antérieurs à la période acheuléenne du Paléolithique inférieur est prouvé par le fait que l’on trouve fréquemment des outils acheuléens gisant à la surface des moraines déposées par ces anciennes banquises33.
Au nord s’étendait l’immense champ de glace scandinave, qui s’étendait sur la Grande-Bretagne et au-delà des vallées du Rhin, de l’Elbe et de la Vistule, atteignant presque les Carpates. Même les plus petites chaînes de montagnes étaient couvertes de glaciers, notamment l’Atlas en Afrique du Nord.
En Amérique du Nord, à partir du grand centre à l’ouest de la baie d’Hudson, la calotte glaciaire a étendu sa dérive vers le sud jusqu’au Missouri, à l’Iowa, au Kansas et au Nebraska, au-delà des limites des glaciations antérieures et ultérieures.
[ p. 90 ]
Les matériaux des principales « hautes terrasses » des grandes vallées fluviales d’Europe occidentale ont été déposés à cette époque.
¶ La vie du deuxième étage interglaciaire chaud
La longue période chaude qui a suivi la grande glaciation est remarquable en ce qu’elle présente les premières preuves de la présence humaine en Europe occidentale. C’est l’époque de l’espèce humaine de Heidelberg (Homo heidelbergensis), connue uniquement par une mâchoire unique découverte par Schoetensack dans les sables de Mauer, près de Heidelberg, en 1907. Aucune autre preuve de l’existence humaine n’a été trouvée dans les gisements formés durant ce vaste intervalle de temps géologique, à moins d’accepter la théorie de Penck et de Geikie selon laquelle les carrières pré-chelléennes et chelléennes de la Somme appartiennent au deuxième stade interglaciaire.
La longue durée de cette période interglaciaire est attestée, tant en Europe qu’en Amérique, par la profonde érosion des « congères » provoquées par la deuxième glaciation. Penck estime que cette « longue période chaude » représente une période plus longue que l’intervalle entre la troisième glaciation et l’époque actuelle. Le climat qui a immédiatement suivi le retrait des glaciers était frais et humide dans les régions glaciaires, mais il a été suivi d’une période de chaleur et de sécheresse si prolongée que les glaciers des Alpes se sont retirés bien au-delà de leurs limites actuelles.
Sur l’une des anciennes « hautes terrasses » de l’Inn, dans le nord du Tyrol, se trouve un dépôt contenant la flore forestière dominante de l’époque, ce qui permet à Penck de conclure que le climat d’Innsbruck était supérieur de 2 °C à son niveau actuel. Par conséquent, la limite des neiges éternelles se situait à 300 mètres au-dessus de son niveau actuel, et les Alpes, à l’exception des plus hauts sommets, étaient presque entièrement dépourvues de glace et de neige. Une plante caractéristique est le rhododendron pontique (Rhododendron ponticum), qui prospère aujourd’hui à une température ambiante de 14 °C à 18 °C, ce qui indique que le climat d’Innsbruck était aussi clément que celui des versants italiens des Alpes aujourd’hui. Ce rhododendron se trouve aujourd’hui dans le Caucase. Parmi les autres espèces méridionales de l’époque, on trouve le nerprun, [ p. 91 ] apparenté à une espèce vivant aujourd’hui aux îles Canaries, et au buis. On y trouvait également des plantes plus rustiques, notamment le sapin (Pinus sylvestris), l’épicéa, l’érable, le saule, l’if, l’orme, le hêtre et le sorbier. Les forêts de la même époque en Provence étaient, pour la plupart, semblables à celles que l’on trouve aujourd’hui dans cette région ; sur trente-sept espèces, vingt-neuf sont encore présentes dans cette partie du sud de la France. Dans l’ensemble, l’aspect du sud de la France à cette époque était étonnamment moderne. Les forêts comprenaient des chênes, des ormes, des peupliers, des saules, des tilleuls, des érables, des sumacs, des cornouillers et des aubépines. Parmi les plantes grimpantes se trouvaient la vigne et la clématite. On y trouvait également quelques formes qui ont depuis reculé vers le sud, comme des espèces de laurier-rose et de laurier, aujourd’hui confinées aux îles Canaries. La forte humidité de l’époque est indiquée par la présence de certaines espèces de conifères, exigeant une humidité considérable. Comme au premier interglaciaire, la présence de figuiers indique des hivers doux.
Il est difficile d’imaginer des forêts de ce caractère moderne, qui plus au nord abritaient un certain nombre d’espèces encore plus tempérées et rustiques, comme le cadre de la grande vie africaine et asiatique qui régnait alors dans toute l’Europe occidentale. C’est la présence d’hippopotames, d’éléphants et de rhinocéros qui a donné à Lyell, Evans et d’autres observateurs primitifs l’impression qu’une température et une végétation tropicales étaient caractéristiques de cette longue période de vie. Ces animaux étaient autrefois considérés comme la preuve d’un climat quasi tropical, mais les preuves plus fiables des forêts, renforcées par la présence de très nombreux types robustes d’animaux des forêts et des prairies, ont infirmé toutes les théories anciennes sur les températures extrêmement chaudes du deuxième interglaciaire.
Les vestiges de ce que l’on appelle encore communément la « faune chaude » se trouvent principalement dans les sables et graviers des anciens lits du Neckar, de la Garonne et de la Tamise, ainsi que d’autres fleuves du nord et du sud, ainsi que dans l’Essex, en Angleterre. Le fait le plus surprenant est que la vie des mammifères d’Europe occidentale est restée entièrement inchangée lors de la vaste deuxième glaciation que nous venons de décrire ; les quelques extinctions qui ont eu lieu, ainsi que le nombre de nouveaux arrivants, peuvent être attribués à de nouvelles connexions géographiques avec l’Afrique au sud et à la progression constante des migrations en provenance d’Extrême-Orient.
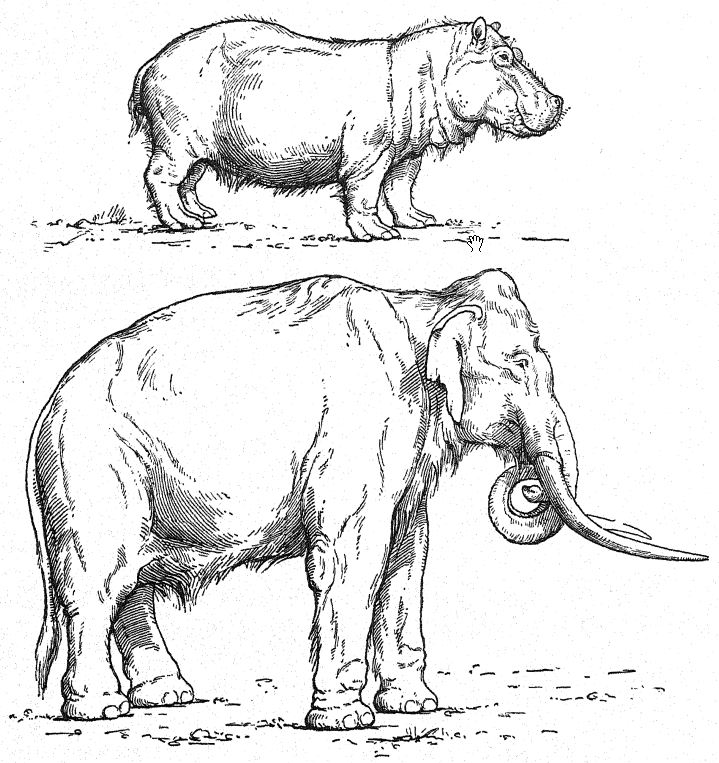
Il y eut quatre nouveaux arrivants très importants et distinctifs du monde afro-asiatique, à savoir l’éléphant à défenses droites ou ancien (E. antiquus), le rhinocéros à nez large (D. merckii), le lion d’Afrique (Felis leo) et l’hycène d’Afrique (H. striata), qui témoignent de liens géographiques étroits avec l’Afrique du Nord. Parmi ceux-ci, l’éléphant ancien et le rhinocéros à nez large étaient de proches compagnons ; ils jouissaient des mêmes régions et des mêmes températures, leurs restes sont très fréquemment retrouvés ensemble, et ils ont survécu jusqu’à la toute fin de la grande période de vie de l’Europe occidentale, qui s’est terminée avec l’avènement de la quatrième glaciation. Ils contrastent avec l’autre paire de grands mammifères déjà présente en Europe au Pliocène et au Premier Interglaciaire, à savoir le mammouth du sud, connu à ce stade sous le nom de trogontJierii, qui avait une préférence pour la compagnie de l’hippopotame (H. major) ; il semblerait que ces animaux étaient moins robustes car tous deux [ p. 94 ] ont disparu d’Europe un peu plus tôt que l’éléphant ancien et le rhinocéros de Merck.
Le lion d’Afrique semble avoir été un concurrent du tigre à dents de sabre, car ce dernier est aujourd’hui moins abondant, bien qu’il y ait des raisons de croire qu’il a survécu jusqu’au troisième interglaciaire. Avec l’ancien type pliocène du tigre à dents de sabre, on a également trouvé le rhinocéros étrusque, l’ours primitif d’Auvergne (Ursus arvernensis) et le rhinocéros géant (Trogontherium cuvieri).
Français Les forêts du nord de l’époque étaient fréquentées par l’élan à face large, le cerf géant et le chevreuil, ainsi que par de nobles spécimens de cerf (Cervus elaphus). Dans les forêts ouvertes et les prairies, le bétail sauvage (Bos primigenius) commença à être plus nombreux et le bison (Bison priscus) était également présent. Parmi les formes fréquentant les prairies ou les forêts se trouvaient des chevaux de plus grande taille, tels que les chevaux de Mosbach et de Süssenborn. Dans cet assemblage de types septentrionaux et méridionaux, il est à noter que les types de mammifères des forêts et des prairies eurasiennes prédominent largement en nombre et en variété sur les types afro-asiatiques ; ceci, avec la flore, est une indication que le climat était de caractère tempéré ; Il est donc probable que tous les mammifères étaient bien protégés par une couverture velue et adaptés à un climat tempéré. Le fait que la faune dans son ensemble soit restée pratiquement inchangée tout au long de la deuxième glaciation prouve non pas qu’elle a migré vers le sud puis est revenue, mais que les régions non glaciaires d’Europe occidentale étaient tempérées plutôt que froides.
¶ La course d’Heidelberg
Homme de Heidelberg. Éléphant antique. Rhinocéros étrusque. Cheval de Mosbach. Sanglier. Élan à face large. Cerf élaphe. Chevreuil. Bison primitif (bison des marais). Aurochs primitif (urus). Ours d’Auvergne. Ours de Deninger. Lion. Chat sauvage. Loup. Castor.
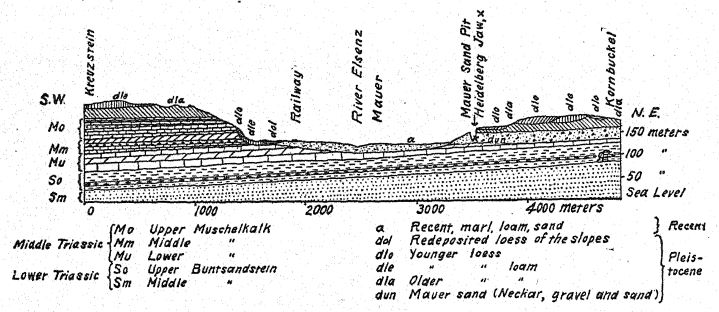
Français Pour nous, la vie mammalienne de loin la plus intéressante est celle découverte au sud de l’embouchure du Neckar, le long de l’ancien ruisseau Elsenz, où se sont déposés les « sables de Mauer » inférieurs, contenant la mâchoire inférieure de l’Homme de Heidelberg et les restes de nombreux animaux de cette période. Le dénombrement de toute cette faune est très important, car il indique les conditions climatiques tempérées qui entouraient la première véritable espèce humaine découverte jusqu’à présent en Europe. Le découvreur, Schoetensack,35, a attribué ces mammifères et l’Homme de Heidelberg à la Première Étape Interglaciaire, et une opinion similaire a récemment été exprimée par Geikie. La présence du rhinocéros étrusque semble indiquer une telle antiquité, mais les preuves fournies par cet animal primitif sont éclipsées par celles de trois mammifères très caractéristiques de la Deuxième Étape Interglaciaire : le rhinocéros à défenses droites ou [ p. 96 ] l’éléphant ancien (E. antiquus), le lion et le cheval de Mosbach. À l’exception du rhinocéros étrusque, toutes ces espèces fréquentant l’ancien ruisseau Elsenz et déposées avec les « sables de Mauer » se trouvaient également dans les forêts et les prairies de la région maintenant connue sous le nom de Bade, où se trouvent les gisements de mammifères fossiles de Mosbach près du Neckar. Une vie mammalienne similaire d’une époque un peu plus récente se rencontre dans les graviers fluviaux de Süssenborn, près de Weimar. Les chevaux de Mauer, de Mosbach et de Süssenborn[4] étaient de taille beaucoup plus grande et d’un caractère plus spécialisé que le cheval de Steno de la première période interglaciaire.
Français Ainsi, les Heidelberg, la première race humaine recensée en Europe occidentale, apparaissent dans le sud de l’Allemagne au début du deuxième interglaciaire, au milieu d’une faune mammalienne très imposante d’aspect nordique et contenant de nombreuses espèces forestières, telles que l’ours, le cerf et l’élan ; dans les prairies et les forêts broutées, l’éléphant géant aux défenses droites (E. antiquus), de par la structure simple de ses dents brouteuses, est considéré comme similaire dans ses habitudes à l’éléphant d’Afrique qui habite maintenant les forêts d’Afrique centrale ; la présence de cet animal indique un climat relativement humide et un pays bien boisé. Le rhinocéros étrusque [ p. 97 ] différait de la forme plus grande de Merck par la possession de dents brouteuses à couronne relativement courte, adaptées aux habitudes de broutage et à un pays forestier ; sur la tête portaient deux cornes ; c’était un type à longs membres, se déplaçant rapidement ; les troupeaux [ p. 98 ] de bisons et de bovins sauvages (urus) qui parcouraient les plaines étaient désormais sujets à l’attaque du lion.

ab. « Lœss récent », soit du Troisième Interglaciaire, soit du Postglaciaire.
av. J.-C. « Lœss ancien » (lœss sableux) de la fin du Deuxième Interglaciaire.
cf. Les « sables de Mauer ».
de. Couche intermédiaire d'argile.
La croix blanche (X) indique l'endroit, à la base des « sables de Mauer », où la mâchoire de Heidelberg a été découverte.
La découverte en 1907 d’une mâchoire inférieure humaine au fond des « sables de Mauer » est l’une des plus importantes de toute l’histoire de l’anthropologie. La découverte a été faite à une profondeur de 24,10 m (79 pieds) au-dessus d’une haute falaise (fig. 46), dans d’anciens sables fluviaux connus depuis longtemps pour abriter la très ancienne faune mammalienne décrite ci-dessus. Pendant des années, les ouvriers avaient reçu pour instruction de surveiller attentivement les restes humains. La mâchoire avait manifestement dérivé avec les sables fluviaux et s’était détachée du crâne, mais elle était restée parfaitement préservée. La description de l’auteur peut être citée en premier lieu.36 La mandibule présente une combinaison de caractéristiques jamais observées auparavant chez un fossile ou un homme récent. La protubérance de la mâchoire inférieure, juste sous les dents de devant, qui donne forme au menton humain, est totalement absente. Sans les dents, il aurait été impossible de diagnostiquer l’humain. D’après un fragment de la symphyse de la mâchoire, on pourrait bien le classer comme un anthropoïde ressemblant à un gorille, tandis que la branche ascendante ressemble à celle d’une grande variété de gibbon. La certitude absolue que ces restes soient humains repose sur la forme des dents : molaires, prémolaires, canines et incisives sont toutes essentiellement humaines et, bien que de forme quelque peu primitive, ne présentent aucune trace d’intermédiaire entre l’homme et les singes anthropoïdes, mais plutôt de descendance d’un ancêtre commun plus ancien. Les dents, cependant, sont plutôt petites pour la mâchoire ; la taille du bord permettrait le développement de dents beaucoup plus grandes ; nous ne pouvons que conclure qu’aucune contrainte importante n’a été exercée sur les dents et que, par conséquent, le puissant développement des os de la mâchoire n’était pas conçu à leur avantage. La conclusion est que la mâchoire, considérée comme incontestablement humaine de par la nature de ses dents, se situe à proximité du point de séparation entre l’homme et les singes anthropoïdes. Comparée aux mâchoires des races néandertaliennes, telles que celles découvertes à Spy, en Belgique, et à Krapina, en Croatie, on peut considérer la mâchoire de Heidelberg comme pré-néandertalienne ; il s’agit en fait d’un type généralisé.
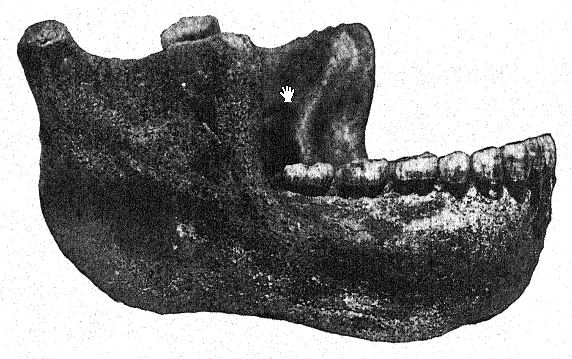
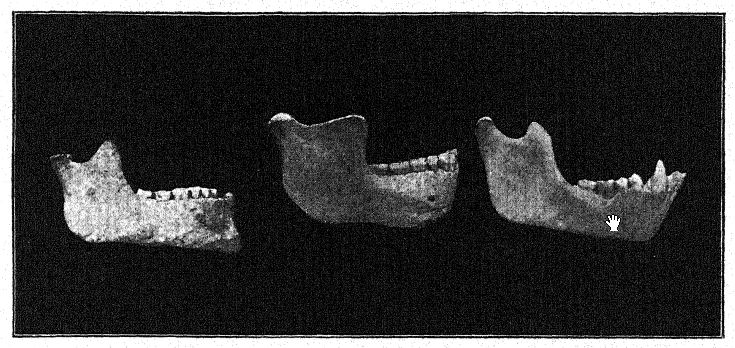
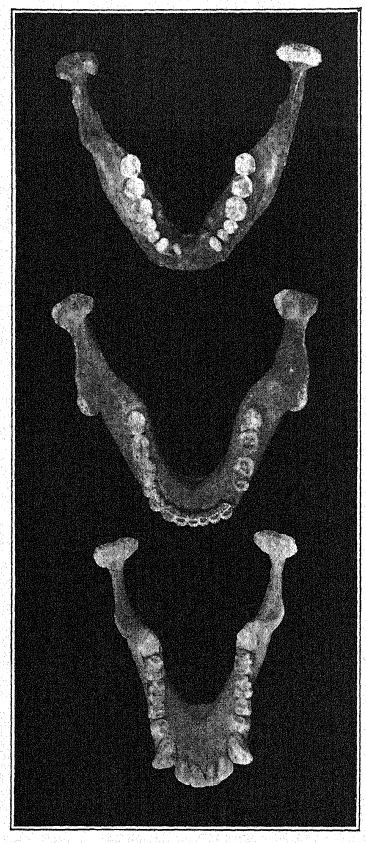
Dans un esprit conservateur, Schoetensack a nommé le type représenté par cette mâchoire Homo heidelbergensis. D’autres auteurs l’ont considéré comme d’un rang générique distinct ; ainsi, Bonarelli l’a appelé Palaoanthropus heidelbergensis.37 La mâchoire elle-même est extrêmement massive ; les canines, contrairement à celles des singes anthropoïdes et de la race de Piltdown, ne dépassent pas la ligne des autres dents et n’étaient donc pas utilisées comme armes d’attaque et de défense comme chez les anthropoïdes, chez qui ces dents [ p. 100 ] sont largement développées en défenses. Comme l’a noté Schoetensack, les dents ne sont pas très massives par rapport à la mâchoire elle-même, qui est la mâchoire humaine la plus puissante connue, dépassant même la plus grande mâchoire esquimaude et indiquant un crâne de caractère très massif et primitif. Elle ressemble à celle du singe par la récession du menton, d’où son nom d’amentalis. Le processus coronoïde de la mandibule est fortement développé pour l’insertion du muscle temporal. Cette mâchoire pourrait bien avoir servi d’outil lors des dernières étapes de la préparation des peaux, comme le font les Esquimaux. On observe que les puissantes branches osseuses de la mâchoire, vues de dessus, se referment sur l’espace laissé à la langue ; en fait, l’os se referme à tel point qu’il gêne sérieusement le libre usage de la langue dans la parole articulée.
Il semblerait que, dans la mâchoire, et probablement dans tous les autres caractères du crâne, à mesure qu’ils seront connus, la race de Heidelberg se révèle être un Néandertalien en devenir, c’est-à-dire une forme ancestrale primitive, plus puissante et plus simiesque. En ce qui concerne le menton fuyant, les véritables Néandertaliens de Spy, Malarnaud, Krapina, [ p. 101 ] et La Chapelle se situent exactement à mi-chemin entre les races les plus inférieures de l’homme moderne et les singes anthropoïdes.
Non seulement chez les Esquimaux, mais généralement chez les races sauvages d’Australie et d’autres pays, les mâchoires sont utilisées comme outils ; chez les Australiens, les dents sont très usées, mais dans un état de conservation admirable. Vues d’en haut, on observe que les dents de Heidelberg forment un arc parfait, ou une disposition en fer à cheval, alors que chez tous les singes, les deux rangées de dents sont presque parallèles. Ainsi, malgré de grandes divergences d’opinions quant aux liens de parenté de l’homme de Heidelberg, tous s’accordent à dire que la découverte de Schoetensack nous offre l’un des grands chaînons manquants de la chaîne du développement humain.

Français La vie mammifère typique du Second Interglaciaire, telle qu’on la trouve à Mosbach et Sussenborn, appartient peut-être à une étape un peu plus récente du Second Interglaciaire que celle des « sables de Mauer », car dans ces localités, le rhinocéros étrusque [ p. 102 ] fait défaut et le rhinocéros à nez large, plus spécialisé, est abondant ; cet animal diffère de la forme étrusque par la possession de dents broyeuses à couronne relativement longue, mieux adaptées aux habitudes de pâturage. Sur la tête portaient deux cornes. Une variété de mammouth du sud (E. trogontherii) est si hautement caractéristique du Second Interglaciaire que Pohlig fait référence à cette période de vie comme le stade E. trogontherii. De par la structure de ses dents broyeuses, on le considère comme semblable à l’éléphant d’Asie, qui peuple aujourd’hui les forêts de l’Inde, mais il possède le front concave particulier, caractéristique du mammouth, et très différent du front convexe de l’éléphant indien. Les ours de cette période appartiennent aux espèces primitives U. deningeri et U. arvernensis, car il n’existe jusqu’à présent aucune trace certaine de la présence du véritable ours brun d’Europe (U. arctos). Le tigre à dents de sabre de cette époque est préservé dans les cavernes des Pyrénées, près de Montmaurin, associé aux restes de la hyène rayée (H. striata), une espèce largement répandue en Europe occidentale au début du Pléistocène. Cette espèce était contemporaine de la hyène tachetée (H. crocuta), puis remplacée par elle, dont descend la très robuste hyène des cavernes (H. crocuta spelcea) de la « Période du Renne ». Nous constatons que le cerf polycladin du Pliocène supérieur et du Premier Glaciaire a disparu d’Europe occidentale ; il n’existe pas non plus de traces du cerf axis. L’hippopotame est rigide, représenté par l’espèce géante, H. major.
¶ Premières migrations des rennes vers le nord
Les animaux que nous avons décrits appartiennent aux régions chaudes et tempérées d’Europe. Le renne était déjà présent dans les régions proches des glaciers ; cet animal typiquement nordique est d’ailleurs signalé dans les graviers de Süssenborn, près de Weimar.
Il existe des preuves d’une succession de changements climatiques dans la région de Heidelberg. La mâchoire de Heidelberg, avec sa faune mammalienne tempérée, se trouvait à la base même de la falaise de Mauer. [ p. 103 ] mais plus haut sur la falaise (Fig. 46), à un niveau correspondant, on trouve des restes de mammifères qui indiquent une baisse marquée de la température et que certains experts attribuent à la période de climat froid qui a caractérisé l’Europe du Nord vers la fin du deuxième interglaciaire. Le renne est également présent dans les graviers des « hautes terrasses » de la rivière Murr, près de Steinheim ; ainsi, à Mauer, à Sussenbom et à Steinheim, nous trouvons la preuve que le renne avait commencé à se répandre dans les régions les plus froides d’Europe, et il y a des raisons de croire qu’il a trouvé son chemin jusqu’aux Pyrénées.
Les traces de la première période froide et aride, qui a profondément influencé le climat de l’Europe occidentale à cette époque, se trouvent également dans la couche de loess ancien, située sur la falaise au-dessus des sables de Mauer. Ce loess recouvre les dépôts mammifères chauds des sables de Mosbach, ainsi que les hautes terrasses de nombreuses vallées fluviales anciennes. En Europe comme en Amérique, la séquence climatique du deuxième interglaciaire, de l’humide au sec, semble avoir été la même.
Ainsi, après le recul des glaciers de la deuxième glaciation, le climat fut d’abord froid et humide ; suivit ensuite une longue période chaude, favorable à l’extension des forêts ; celle-ci fut finalement suivie d’une période d’aridité au cours de laquelle se sont produits les plus anciens dépôts de « lœss ». En Russie également, la troisième glaciation fut précédée d’un climat aride et steppique, avec des vents violents favorables au transport du « lœss ».
Aucun palésolithe ni autre preuve d’occupation humaine n’a été découvert durant cette période froide et sèche, car il n’existe aucune trace, en Europe, de campements dans ce « lœss ancien », comme on en trouve dans le « lœss » déposé durant la même période aride, vers la fin du troisième interglaciaire, puis durant la période postglaciaire. Nous n’avons pas non plus trace de vie mammalienne dans ce « lœss ancien » d’Europe.
[ p. 104 ]
¶ La troisième glaciation
Cette période aride en Europe du Nord et en Amérique du Nord fut suivie par le climat humide et frais de la troisième glaciation. Penck estime que l’avancée de ces nouveaux champs de glace a commencé il y a 120 000 ans et que la période d’avancée et de recul des glaciers n’a pas duré moins de 20 000 ans. Dans les Alpes, la limite des neiges s’est abaissée de 1 250 mètres sous le niveau actuel ; par conséquent, cette glaciation fut plus sévère que la première, mais légèrement moins sévère que la seconde. En Europe du Nord, le champ de glace scandinave ne couvrait pas une zone aussi étendue que lors de la seconde glaciation, bien que la Grande-Bretagne et la Scandinavie aient été à nouveau profondément ensevelies sous la glace ; la calotte glaciaire et les glaciers s’écoulaient vers l’ouest et le sud-ouest à travers le Danemark et la partie sud du bassin de la Baltique jusqu’en Hollande et dans le nord de l’Allemagne. Dans les Alpes, la troisième glaciation a déplacé d’immenses plaques de glace le long de la vallée du Rhin, jusqu’à l’est de la France et dans la vallée du Pô, où elle a été encore plus étendue que la seconde. Mais le plus grand glacier de cette époque était celui de l’Isar, un affluent méridional du Danube, qui prend sa source dans les Alpes bavaroises38.
Au cours de la troisième période glaciaire, certaines des « terrasses moyennes » le long du Rhin et d’autres fleuves descendant des Alpes se sont formées. En Grande-Bretagne39, alors que pendant la deuxième glaciation les champs de glace s’étendaient jusqu’à la Tamise au sud, pendant la troisième glaciation ils ne dépassaient pas les Midlands ; pourtant, un climat arctique prévalait dans le sud de l’Angleterre, avec des conditions et une température de toundra, comme l’indiquent les dépôts végétaux de Hoxne40 dans le Suffolk. Avant même le début de la troisième glaciation en Europe, une grande calotte glaciaire s’était formée au-dessus du Labrador, sur la côte est de l’Amérique du Nord, et les calottes glaciaires s’écoulant vers le sud et le sud-ouest s’étendaient jusqu’à l’Illinois, déposant les grandes « congères » illinoises.
[ p. 105 ]
Français Le long des frontières de ces grands champs de glace, dans les deux pays, régnait un climat froid et humide, car l’une des principales conditions de la glaciation est la forte précipitation de neige. En Europe du Nord, entre les grands champs de glace alpins et scandinaves de la troisième glaciation, un climat froid régnait sans aucun doute ; dans la région du Neckar, près de Cannstatt, se trouve un gisement connu sous le nom de « loam à mammouths », que Koken croyait contemporain de la période de la troisième glaciation, bien que les preuves ne soient certainement pas convaincantes41. On y trouve des restes fossiles de rennes b^dinaviens, ainsi que de deux nouveaux arrivants très importants en Europe en provenance des régions de toundra de l’extrême nord-est, ar,imglg [ p. 106 ] qui avait erré le long des frontières méridionales de la calotte glaciaire scandinave depuis les toundras du nord de la Russie et de la Sibérie* C’est la première apparition en Europe occidentale du mammouth laineux (E. primigenius) et du rhinocéros laineux (D. antiquitatis). Dans ce « terreau à mammouths », on trouve également deux espèces de chevaux, le cerf géant (Megaceros), le cerf, le bison et l’aurochs. Si le mammouth laineux et le rhinocéros laineux sont effectivement entrés dans l’est de l’Allemagne à cette époque, ils se sont certainement retirés vers le nord à l’approche du climat tempéré chaud du troisième stade interglaciaire, car aucune trace de ces animaux n’a été retrouvée en Europe avant l’avènement de la quatrième glaciation.
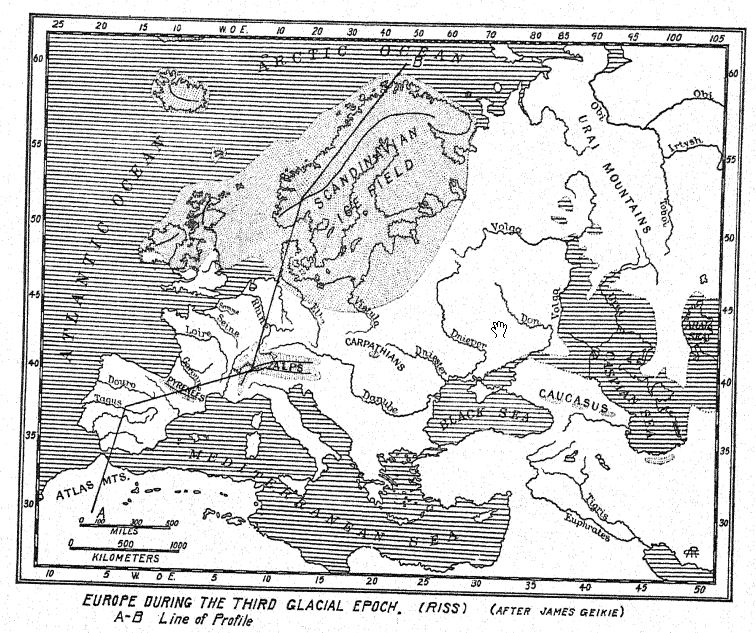
¶ Bibliographie
(1) Gaudry, 1890.1.
(2) Smith, GE, 1912.1, p. 582.
(3) Op. cit.
(4) Munro, 1893.1.
(5) Osborn, 1910.1, pp. 306, 307.
(6) Geikie, J., 1894.1, pp. 329-336; 1914.1, p. 227.
(7) Dawkins, 1883.1, pp. 576-579.
(8) Geikie, J., 1914.1, p. 248.
(9) Reid, C., 1908.1.
(10) Desnoyers, 1863.1.
(11) Haug, 1911.1, p. 1S07.
(12) Geikie, J., 1894.1, p. 682 ; 1914.1, p. 250.
(13) Dubois, 1894.1.
(14) Fischer, 1 913. 1.
(15) Schwalbe, 1899.1; 1914.1.
(16) Büchner, 1914.1.
(17) Vol., 1907. 1.
(18) Elbert, 1908.1.
(19) Sélénka, 1911.1.
(20) Pèlerin, 1913.1.
(21) Schwalbe, 1899.1, p. 227, 228. (22) Op. cit., p. 223.
(23) Schwalbe, 1914. i, pp. 601-606.
(24) Buchner, 1 914.1, p. 129.
(25) Dubois, 1894,1, p. 14.
(26) Keith, 1912. 1.
(27) Smith, GE, 1912. 1, p. 595.
(28) Op. cit.
(29) Rutot, 1907.1.
(30) Boule, 1913. 1, pp.'266, 267.
(31) MacCurdy, 1 905.1, pp. 468, 469.
(32) Geikie, J., 1914.1, p. 251.
(33) Op. cit. p. 255.
(34) Op. cit. p. 238.
(35) Schoetensack, 1908.1.
(36) Op. cit. pp. 25-43.
(37) Bonarelli, 1909.1.
(38) Penck, 1909. 1.
(39) Geikie, J., 1914.1, p. 258.
(40) Op. cit., pp. 257-262.
(41) Schmidt, 1912.1, p. 181
¶ Notes de bas de page
Un article récent de A. Smith Woodward décrit le quatrième spécimen connu de Dryopithecus, récemment découvert dans le nord de l’Espagne (voir Woodward, 1914.2). ↩︎
Il existe une vaste littérature sur le Pithécanthrope. Celle principalement utilisée dans la présente description comprend Dubois,13 Fischer14 Schwalbe,15 Buchnerd16. ↩︎
Dans le crâne de Trinil tel que restauré par McGregor (Fig. 36), la capacité crânienne est de 900 cm³. ↩︎
Ces chevaux sont désormais identifiés respectivement comme E. mauerensis, E. moshachensis et E. süssenbornensis. ↩︎
Cette glaciation telle qu’elle se produit en Europe du Nord a été appelée Polandienne par Geikie ; dans les Alpes, Penck l’a appelée l’Illinoisienne ; en Amérique, elle est connue sous le nom d’Illinoisien en raison des grandes dérives qu’elle a déposées sur l’État de l’Illinois. ↩︎