[ p. 186 ]
FIN DU TROISIÈME INTERGLACIAIRE. CLIMAT TEMPÉRÉ ET ARIDE. INDUSTRIE ACHÉULÉENNE — AVENUE DE LA QUATRIÈME GLACIATION, PROFONDEURS CHANGEMENTS DANS LA VIE ANIMALE ET VÉGÉTALE — LA TOUNDRA ARCTIQUE, PÉRIODE DE LA VIE MAMMIFÈRE ET VÉGÉTALE — CARACTÈRES DE LA RACE NÉANDERTALIENNE, DE SON INDUSTRIE DU SILEX MOUSTÉRIEN — CAUSES SUPPOSÉES D’EXTINCTION OU DE DISPERSION
Nous arrivons maintenant à une étape prolongée et importante de la préhistoire de l’Europe, à savoir la période de la quatrième glaciation, du développement final de la race humaine de Néandertal, de l’industrie moustérienne, des débuts de la vie dans les cavernes, de la chasse au renne et de son utilisation pour l’alimentation et l’habillement.
Dans toute l’Europe, l’industrie acheuléenne semble avoir pris fin durant une période de climat aride, chaud dans certaines régions d’Europe occidentale et frais, voire froid, dans d’autres. Les variations saisonnières ont pu être extrêmes, comme dans les steppes du sud de la Russie, où des étés extrêmement chauds peuvent être suivis d’hivers extrêmement froids, avec des vents violents et des tempêtes de neige destructrices de vie.
C’est cette alternance saisonnière, ainsi que la récurrence, saisonnière ou séculaire, du climat plus froid, qui explique la survie ou le retour de la faune asiatique même après la fin de l’industrie acheuléenne et lorsque l’industrie moustérienne était bien avancée.
Français D’après les dépôts découverts à Grimaldi, dans la grotte des Enfants et dans la grotte du Prince, on a longtemps affirmé que les hommes du Moustérien ancien vivaient contemporains de l’hippopotame, de l’éléphant à défenses droites et du rhinocéros de Merck dans le climat clément de la Riviera méditerranéenne. Plus récemment, les mêmes animaux ont été retrouvés aussi loin au nord que la vallée de la Somme, dans les « river-drifts » de Montières-les-Amiens.1 Ici encore, nous trouvons des restes 186 [ p. 187 ] de l’hippopotame, de l’éléphant à défenses droites et de son compagnon, le rhinocéros de Merck, dans des dépôts moustériens, une découverte surprenante, car on avait toujours supposé qu’une période climatique froide s’était installée dans toute l’Europe occidentale, même avant la fin de la culture acheuléenne. Mais il existe aussi des preuves d’un climat tempéré qui prévalait encore dans la vallée de la Tamise à l’époque des « planchers » moustériens. 2 De même, le long de la vallée de la Vézère, en Dordogne, nous constatons qu’à la station de La Micoque, où l’industrie marque la transition entre l’Acheuléen tardif et le Moustérien précoce, le rhinocéros de Merck se trouve dans les couches les plus basses associées aux restes de l’élan (Alces).
Il existe des preuves que le rhinocéros de Merck et l’éléphant à défenses droites ont persisté en Europe occidentale pendant toute la période du développement précoce de l’industrie moustérienne. Comme observé précédemment, ces animaux étaient plus robustes que le mammouth du sud, qui fut le premier mammifère asiatique à disparaître, bientôt suivi par son compagnon, l’hippopotame. Même après l’apparition du couple étroitement associé de la toundra, le mammouth laineux et le rhinocéros laineux, le rhinocéros de Merck persiste, comme par exemple dans les gisements de Rixdorf, près de Berlin, où ce type ancien est présent dans les mêmes gisements que le mammouth laineux, le rhinocéros laineux, le renne et le bœuf musqué, ainsi que chez les formes forestières, l’élan, le cerf, le loup et le cheval des forêts. La latitude extrême nord de ce gisement explique l’absence de l’éléphant à défenses droites, qui vivait peut-être à cette époque plus au sud. Le même mélange de mammifères d’Asie du Sud et d’Asie du Nord se retrouve à Steinheim, dans la vallée de la Murr, à quelques degrés à l’ouest et au sud de Rixdorf, non loin de Göttingen, où l’on trouve le rhinocéros de Merck® et l’éléphant à défenses droites en association avec le mammouth laineux, le rhinocéros laineux, le cerf géant et le renne.
Français Ainsi, les races néandertaliennes entraient dans le stade culturel moustérien à la fin de la troisième période interglaciaire et au début de la progression des champs de glace depuis les grands centres de Scandinavie et des Alpes. Alors que ces champs de glace [ p. 188 ] se rapprochaient lentement l’un de l’autre du nord et du sud, une très longue période de temps a dû s’écouler pendant laquelle tous les mammifères d’Asie du Sud ont abandonné l’Europe occidentale ou ont disparu, à l’exception des lions et des hyènes, qui se sont bien adaptés au climat très rigoureux qui a prévalu sur l’Europe pendant la quatrième glaciation, et même pendant la longue période postglaciaire qui a suivi. Les grands carnivores s’adaptent facilement et complètement aux climats froids, car ils se nourrissent de la vie animale où qu’elle se trouve ; Des tigres de la même souche que ceux de l’Inde ont été trouvés aussi loin au nord que la Léna, à 52° 25’ de latitude, où le climat est plus froid que celui de Petrograd ou de Stockholm, tandis que le lion prospérait dans l’atmosphère froide du haut Atlas. Ainsi, le lion des cavernes (Felis leo spelaea) et l’hysène des cavernes (H. crocuta spelaea) ont sans doute développé une sous-couche de fourrure ainsi qu’une couverture de poils longs, comme les mammifères de la toundra. En taille, le lion de cette période en France égalait souvent, et parfois surpassait, ses parents actuels, le lion d’Afrique et d’Asie occidentale ; il figure fréquemment dans l’art des artistes du Paléolithique supérieur et a survécu en Europe occidentale jusqu’à la fin du Paléolithique supérieur.
¶ La quatrième glaciation
Penck4 a estimé que le premier maximum de la quatrième glaciation dans les Alpes a été atteint il y a 40 000 ans, et qu’après la période de récession, le second maximum s’est terminé il y a au moins 20 000 ans. Cela prolongerait l’industrie moustérienne sur une très longue période, car il ne fait aucun doute que la culture moustérienne était pratiquement contemporaine de la quatrième glaciation, même si une période plus brève devait être attribuée à ce grand événement naturel.
La quatrième glaciation, comme la première, aurait eu lieu simultanément en Europe et en Amérique du Nord5, un fait particulièrement important pour les anthropologues américains concernant la date d’arrivée de l’homme primitif en Amérique. Dans les deux pays, la glaciation a atteint un maximum précoce, suivi d’une période de recul des glaciers, période durant laquelle un climat légèrement plus tempéré a prévalu, mais celle-ci a ensuite cédé la place à une seconde avancée, aussi sévère que la première[1].
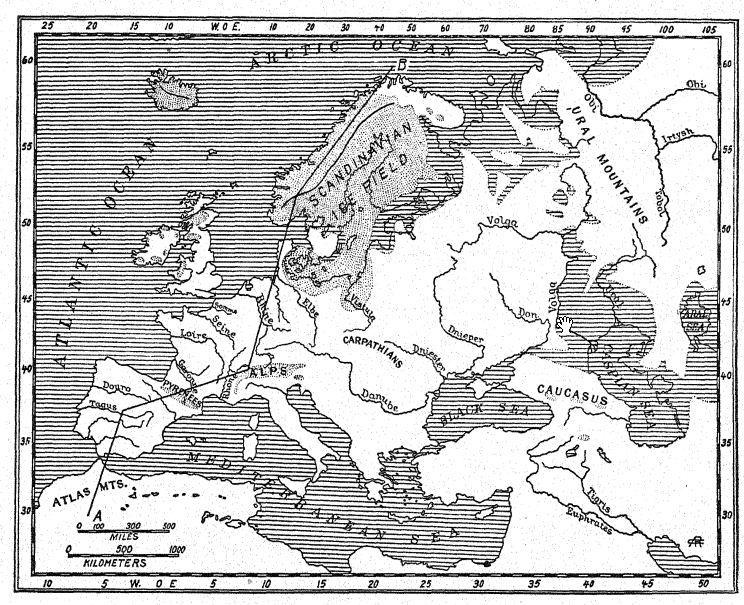
Français Au nord, la Scandinavie et la Finlande furent à nouveau englouties par les glaces, et une grande mer de glace occupa le bassin de la mer Baltique, envoyant ses moraines terminales au Danemark et au Schleswig-Holstein [ p. 190 ] et sur les provinces du nord de l’Allemagne, mais ce grand champ de glace ne conflua plus avec celui de la Grande-Bretagne.7 Au début de la quatrième glaciation, de grands glaciers descendirent sur les vallées des montagnes écossaises et en remplirent beaucoup jusqu’à la mer ; la côte s’affaissa d’au moins 40 mètres dans cette région. Dans le sud de la Grande-Bretagne, le long de la vallée de la Tamise, se répandit une flore arctique, avec le saule polaire (Salix polaris) et le bouleau nain (Betula glandulosa) ; une plante arctique [ p. 191 ] Un lit de toundra a également été découvert dans la vallée de la Lea. Ainsi, le climat de la toundra s’étendait des basses terres écossaises au sud de l’Angleterre, la terre étant aride et presque dépourvue d’arbres.8 C’était, pensons-nous, également la période de la flore arctique à Hoxne, dans le Suffolk, et de la couche végétale arctique dans la vallée de la Tamise. À cette époque, la vallée était fréquentée par le renne, le rhinocéros laineux et le mammouth, dont les restes sont enfouis dans les alluvions basses balayées par les flancs de la vallée, de sorte que les restes de cette faune arctique peuvent par endroits dépasser ceux de la faune asiatique chaude, plus profondément enfouie et beaucoup plus ancienne, de l’époque cheléenne. Comme la Somme, la Tamise® était alors de 10 à 25 pieds sous son niveau actuel, le fond s’étant depuis envasé par des alluvions.

C’est à cette époque que se déposèrent les « dépôts supérieurs » sur les plaines du nord de l’Allemagne, les Alpes et le nord de l’Angleterre, ainsi que les dépôts du Wisconsin ancien et récent, ou « dépôts supérieurs », qui s’étendent très largement sur les États de l’Est, du Wisconsin vers le sud et l’est jusqu’à la latitude de New York. Les graviers et les sables de certaines des « terrasses les plus basses » furent également déposés.
¶ La vie des mammifères à l’époque moustérienne
Les trois phases successives de climat et d’environnement entourant les hommes de Néandertal pendant la période de développement de l’industrie moustérienne, étaient dans l’ordre décroissant comme suit :
3. Climat extrêmement froid de la dernière grande avancée glaciaire. Période d’industrie du Moustérien tardif de La Quina. Propagation de tous les mammifères arctiques et de la toundra en Europe occidentale, y compris le bœuf niusk ; migrations de l’obi et du lemming rayé de l’extrême nord. Vie et industrie des Néandertaliens, principalement dans les abris, les grottes et les entrées des cavernes.
2. Climat froid et humide. Période de l’industrie du Moustérien moyen ou « plein Moustérien » des races néandertaliennes. Apparition de la vie de la toundra, incluant des mammifères et des oiseaux bien protégés de la région arctique, ainsi que la descendance des types alpins vers les contreforts et les bords des rivières. Premiers précurseurs de la vie de la steppe ; la vie des forêts et des champs eurasiens est largement répandue dans toute l’Europe. Vie et industrie principalement dans les abris, les grottes et les entrées des cavernes. Les rennes sont très abondants.
1. Climat aride chaud ou froid. Transition de la culture acheuléenne au Moustérien ancien, observée dans les stations de La Micoque et de Combe-Capelle. La faune dite « moustérienne chaude », comprenant l’hippopotame survivant, le rhinocéros de Merck et l’éléphant à défenses droites dans le nord et le sud de la France ; troupeaux de bisons, de bovins et de chevaux sauvages dans le sud-ouest de la France. Vie tribale, avec une activité industrielle en partie dans des stations ouvertes, en partie sous des falaises abritées.
C’est le début de la « période du renne », car ce migrateur venu de Scandinavie, accompagné de ses compagnons des toundras du nord-est, le mammouth laineux et le rhinocéros, erra lentement vers le sud avant l’avancée des glaciers scandinaves, considérablement accrus par le climat de plus en plus froid et humide. Ainsi, ces animaux se trouvent dans le nord avec des silex de la culture moustérienne avant d’apparaître dans la région plus clémente de la Dordogne. Dans la station acheuléenne-moustérienne un peu plus ancienne de La Micoque, le long de la Vistule, les foyers contiennent presque exclusivement des restes de chevaux et relativement peu de restes de bisons et de bovins sauvages, mais aucun renne. Un foyer près de la station de Combe-Capelle livre de nombreux restes de bisons, seulement quelques restes de chevaux et les premiers de rennes. Avant l’apparition des rennes dans la vallée de la Vézère, on peut se représenter les prairies couvertes de bisons et de chevaux sauvages, ces derniers du type qui caractérise aujourd’hui les hauts plateaux de l’Asie centrale, tandis que le bison de l’époque paraît plus proche du buffle américain que de la forme européenne survivante.
Progressivement, les animaux de la toundra se sont répandus vers le sud, avec le climat froid qui, pour la première fois, a balayé toute l’Europe occidentale. L’aspect général du pays a lentement changé avec l’arrivée des rennes, et la flore nordique, composée d’épicéas, de sapins et de saules arctiques, a recouvert les vallées fluviales et les coteaux les plus abrités, tandis que les plateaux et les champs étaient partiellement ou totalement déboisés.
Français Ainsi, le pays s’est adapté principalement aux types de mammifères de la toundra ; et dans les strates du Moustérien moyen, ces troupeaux, [ p. 193 ] [ p. 194 ] nouvellement migrés de l’extrême nord et des steppes du nord-est bordant la rivière Obi, sont largement plus nombreux que les formes steppiques, qui sont limitées à deux ou trois espèces. Parmi celles-ci, les principaux types sont le cheval des steppes, apparenté au cheval de Przewalski vivant maintenant dans le désert de Gobi, le suslik des steppes (Spermophilus rupescens), et le tétras des steppes, ou poule d’eau. Les formes les plus caractéristiques de la vie steppique, telles que l’antilope saïga, la gerboise et le kiang, sont toutes arrivées plus tard et ne sont apparues qu’après la fin de l’industrie moustérienne et la disparition de la race néandertalienne.

Cela était dû au fait que le climat entourant la race néandertalienne à l’époque moustérienne était froid et humide, avec de fortes pluies en été et des tempêtes de neige en hiver, un climat parfaitement adapté aux mammifères timdra arctiques avec leur épaisse couverture de poils agissant comme un abri contre la pluie et la sous-couche de laine les protégeant des conditions météorologiques les plus extrêmes.
La vie des mammifères durant la quatrième glaciation, telle qu’elle s’est étendue au Rhin moyen et à la région westphalienne, est pleinement documentée dans les gisements de Toess d’Achenheim et dans la célèbre grotte de Sirgenstein, sur le Danube supérieur, au nord-ouest de Munich. On y trouve, outre des traces de l’industrie moustérienne la plus primitive, des restes de mammouth, de bison, de renne, d’une espèce de cheval sauvage et d’ours des cavernes. Après ces mammifères, on trouve, dans le même gisement, une trace de l’arrivée du lemming Obi, venu du nord de la Russie.
Le fait que seules sept stations moustériennes soient connues dans toute l’Allemagne, ou huit si l’on inclut le site de la sépulture néandertalienne, peut s’expliquer par la proximité relative du grand glacier scandinave au nord, distant de seulement 560 kilomètres du grand glacier alpin au sud. À l’est s’étendaient les plaines de Bohême et la vaste région de plaine s’étendant vers le nord-est jusqu’aux toundras et vers l’est jusqu’aux steppes, traversées par les grandes migrations de la vie dans la toundra et la steppe.
[ p. 195 ]
[ p. 196 ]
¶ Environnement géographique et climatique de la race néandertalienne
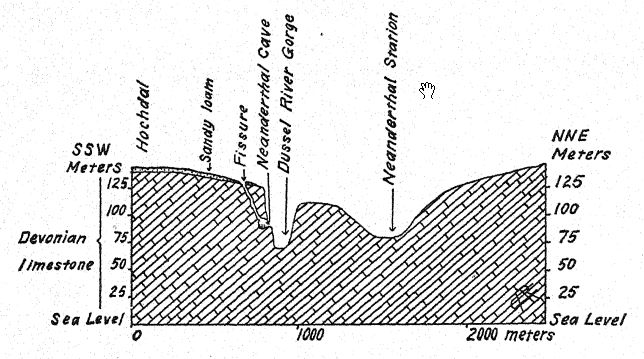
Jetons d’abord un coup d’œil sur la Dordogne. Parmi les stations de l’industrie moustérienne ancienne, nous avons vu que les Néandertaliens de la vallée de la Vézère, à La Micoque, étaient au milieu d’une faune composée principalement de bisons et de chevaux sauvages, les restes trouvés dans les foyers étant presque exclusivement de ce dernier animal[2]. Dans la station moustérienne primitive de Combe-Capelle, proche, les foyers livrent des restes de bisons, mais seulement quelques restes de chevaux.
Parmi les plus anciennes grottes habitées par l’homme10 figure celle du Moustier, située sur la rive droite de la Vézère, à environ 27 mètres au-dessus. Cet abri et cette grotte ont été examinés dès 1860-1863 par Lartet11 et Christy, puis par de Vibraye12, Massenat13 et d’autres. Outre les dépôts présents au fond de la grotte, une profonde couche de culture moustérienne a été découverte sous la falaise en face, et elle a été sélectionnée pour notre représentation de la vie des hommes de l’époque moustérienne et de la flore de la Vézère à cette époque ancienne (voir frontispice). Peyrony observe qu’ici comme ailleurs, les camps industriels les plus anciens et les plus bas étaient plus éloignés des abris ; en effet, dans cette même région, il existe des preuves que les silexiers chelléens et acheuléens visitaient occasionnellement les plateaux situés au-dessus ; mais à mesure que le temps passait et que le climat devenait plus rigoureux, les Néandertaliens commencèrent à travailler plus près des falaises surplombantes, puis directement en dessous. À cette station classique du Moustier, l’un des squelettes les plus complets d’homme de Néandertal fut mis au jour par Hauser en 1908. Il y eut une résidence continue ici au Moustérien moyen et supérieur, s’étendant jusqu’à l’Aurignacien inférieur du Paléolithique supérieur. La faune contemporaine de ces gisements comprenait le mammouth, le renne, le cerf géant (Megaceros), le cheval, le bison, le rhinocéros laineux et l’ours des cavernes. À l’époque où l’homme occupait cette station typique, le climat était très froid et humide.
Dans cette région se trouve le témoignage complet de l’évolution moustérienne, tant au niveau des outils que de l’apparition de nouvelles formes de vie ; le nombre de rennes augmente progressivement dans les couches ascendantes, parallèlement au développement de l’industrie moustérienne. On observe une gradation constante des types industriels acheiléens aux types industriels moustériens ; selon Gartailhac, cette industrie est entièrement l’œuvre du même peuple, sans distinction nette.

Ainsi, à Combe-Capelle, où la véritable culture moustérienne a débuté, on trouve un certain nombre de coups de poing importants, remontant aux premiers outils acheuléens. Les gradations observées ici dans ces couches successives contrastent fortement avec l’essor de l’industrie à la fin du Moustérien dans la même localité, où la transition culturelle vers l’Aurignacien est abrupte.
Le sud de la Grande-Bretagne témoigne d’une séquence similaire, que nous pouvons interpréter comme suit. Appartenant soit au climat tempéré [ p. 198 ] du Moustérien ancien, soit à la période de récession de la quatrième glaciation, connue dans les Alpes sous le nom de Laufenschwankung, se trouvent les stations moustériennes le long de la Lea et près de l’embouchure de la Tamise à Crayford (Worthington Smith,14 Geikie15). Ces « sols » paléolithiques du Moustérien sont enfouis sous 1,20 à 1,50 mètre de sable et de limon et reposent sur la surface de graviers fluviaux plus anciens. Parmi les dépôts fluviaux plus récents, plusieurs surfaces terrestres anciennes ont été découvertes ; elles consistent en quelques centimètres de gravier anguleux, encombrés par endroits d’outils et d’éclats non abrasés qui se trouvent manifestement là où ils ont été laissés par les ouvriers pakolithiques. À un moment donné, on constate que le fabricant de silex était accroupi au-dessus de son ouvrage, les genoux légèrement écartés, car les éclats sont projetés à droite et à gauche en petits tas. Çà et là, réalisées avec ces outils moustériens, on trouve des formes plus archaïques, peut-être issues des terrains plus anciens.

[ p. 199 ]
Un tel sol a été repéré par Worthington Smith16 à travers le Middlesex et des deux côtés de la Tamise. Les vestiges végétaux abondent sur cette ancienne surface terrestre, notamment des empreintes de portions de feuilles, de tiges d’herbe, de joncs et de carex. Le bouleau, l’aulne, le pin, l’if, l’orme et le noisetier ont été reconnus. La fougère mâle commune est fréquente, tandis que la fougère royale (Osmunda regalis) est présente en abondance. Dans l’ensemble, cet assemblage de plantes indique un climat tempéré. Les silex décrits et figurés par Worthington Smith sont soit du type « éclat Levallois » de l’Acheuléen tardif, soit du Moustérien précoce. Cet auteur note le grand nombre d’instruments connus sous le nom d’éclats taillés, que l’on trouve sur le « sol » paléolithique ; ce sont des éclats de grande taille, taillés en forme d’outil sur une face, tandis que l’autre face est laissée parfaitement lisse ; Les exemples sont remarquablement constants quant à leur forme. Le type d’outil décrit ici ressemble aux éclats de Levallois ou de Combe-Capelle, voire à la pointe typique du Moustier. De tels éclats, façonnés en forme de racloir moustérien, sont très courants dans les graviers de la Lea et de la Tamise.
Bien que l’on y trouve des restes de mammouth laineux, on y trouve également des indices de la présence d’une flore tempérée bien marquée. Ces « drifts fluviaux » de haut niveau le long de la Tamise18 se sont certainement déposés lorsque les conditions climatiques étaient tempérées, mais ils sont suivis de dépôts indiquant une nouvelle période froide, qui pourrait correspondre aux périodes froides du « plein Moustérien » de l’occupation de la Tamise au Paléolithique inférieur. On y trouve les remarquables nappes de « drifts » tortueuses attribuables aux mouvements du sol et du sous-sol gelés lorsqu’ils sont exposés à la chaleur du soleil d’été. À la même époque, des loams et graviers alluviaux, contenant parfois des pierres et des roches, ont pu se déposer le long de la Tamise, qui ont été apportés par des radeaux de glace ; ces graviers de bas niveau ne doivent pas être confondus avec les « anciens graviers fluviaux » sous-jacents qui contiennent la faune d’hippopotames tempérée chaude, car ils se sont accumulés dans des conditions très froides ; ils livrent des restes de rhinocéros laineux et [ p. 200 ] de mammouth. Ainsi, sur les niveaux élevés de la Tamise comme sur les niveaux bas, nous trouvons des témoignages de la culture humaine et de la faune éteinte de la période de la quatrième grande glaciation.
Les eaux supérieures du Rhin et du Danube étaient également fréquentées par les silexiers de l’Acheuléen tardif et du Moustérien précoce. À une distance considérable du sud de l’Angleterre se trouve la grotte de Wildkirchli, sur les monts Santis, près d’Appenzell, en Suisse ; à l’époque moustérienne, elle se trouvait au cœur même du champ de glace des Alpes du Nord. La vie animale qui y règne pourrait indiquer que cette grotte était ouverte pendant la période de recul entre les deux grandes avancées de la quatrième glaciation. Ici, à une altitude de 1 370 mètres, Bachler19 a découvert, entre 1903 et 1906, des preuves d’occupation par l’homme de Néandertal à l’époque moustérienne ; les silex ne sont pas bien formés ; la présence d’outils osseux bruts pourrait indiquer une époque moustérienne tardive ; mais Bachler considère que les silex sont du même stade que ceux du Moustier. On affirme que lorsque les Néandertaliens ont suivi la chasse ici [ p. 201 ] le climat était plus clément, car les animaux trouvés comprennent le cerf, le loup alpin (Cyon alpinum fossilis), l’ours des cavernes, le lion des cavernes, le léopard des cavernes (Felis pardus spelaea), le blaireau, la martre et la loutre, ainsi que les formes alpines typiques, le bouquetin, le chamois et la marmotte. Mais cette faune à elle seule peut difficilement être considérée comme la preuve d’un climat tempéré, car à cette altitude alpine, nous ne devrions pas nous attendre à découvrir la vie de la toundra de l’époque ; en fait, elle est totalement absente.


De toutes les stations le long du Danube, la plus importante est de loin celle de Sirgenstein, située entre les villes modernes de Nuremberg et d’Augsbourg. Elle fut d’abord occupée par les Néandertaliens au début du Moustérien et continua d’être visitée par les hommes du Paléolithique inférieur et supérieur jusqu’à la fin du Paléolithique supérieur. La section continue de vie animale et de culture humaine que livre cette remarquable caverne a fourni à Schmidt,20 qui a commencé ses recherches ici au printemps 1906, une clé de la préhistoire des dix-huit cavernes de la région du Haut-Danube et du Haut-Rhin. À Sirgenstein, la culture moustérienne primitive des premiers Néandertaliens a été découverte, ainsi que des restes de mammouth, de bison, de renne, d’une espèce de cheval sauvage et d’ours des cavernes ; Cette industrie moustérienne s’est achevée avec l’enregistrement de l’arrivée dans cette région du lemming Obi en provenance du nord de la Russie. Plus tard, la race Crô-Magnon de l’Aurignacien a laissé sur le sol de la caverne des vestiges de son industrie du silex et de ses festins, notamment des os de rhinocéros laineux, de mammouth, de cerf et de renne. Au cours du Paléolithique supérieur, au Solutréen, la caverne n’était pas occupée ; mais au début du Magdalénien, elle fut de nouveau habitée par l’homme, et coïncidant avec son retour, on assiste à l’arrivée d’une grande migration du lemming rayé (Myodes torquatus) en provenance des toundras arctiques du nord. Enfin, vers la fin du Paléolithique supérieur, à la fin du Magdalénien, une autre transition climatique est indiquée par l’apparition du pika, ou lièvre sans queue (Lagomys pusillus). Durant l’âge du bronze, cette grotte prisée fut à nouveau visitée et habitée pendant une partie de l’âge du fer. Les débris de ces diverses cultures, foyers et dépôts de limon cavernicole atteignent une épaisseur totale de 2,4 mètres et font de Sirgenstein la première station paléolithique d’Allemagne.
¶ Types et migrations des mammifères chassés par les Néandertaliens
Voici la vie à l’époque de la quatrième glaciation, lorsqu’un climat très froid et humide régnait dans toute l’Europe occidentale, jusqu’au nord de l’Espagne et de l’Italie. Bien que les champs glaciaires n’étaient pas aussi étendus que lors de la troisième ou de la deuxième glaciation, le climat était très rigoureux, comme l’indique la migration vers le sud non seulement de la flore arctique, mais aussi des mammifères et des oiseaux de la toundra bordant les rives sud de l’océan Arctique. Deux ou trois formes du froid [ p. 203 ] [ p. 204 ] [ p. 205 ] Les steppes du nord de la Russie ont également trouvé leur chemin vers l’Europe occidentale, mais ce n’était clairement pas une période de steppe en raison de l’humidité dominante du climat ; à la place des vents d’ouest et des grands nuages de poussière de la fin de l’Acheuléen, des brumes froides et des nuages lourds d’humidité ont balayé le pays, qui pendant les hivers était parfois enseveli sous la neige et soumis à de rapides changements de température. Ces conditions climatiques[3] semblent être démontrées par la prédominance de la vie de la toundra arctique, des mammifères qui n’étaient adaptés qu’aux conditions climatiques rigoureuses et attirés par la flore nordique.

Les étés étaient sans aucun doute chauds, comme les étés arctiques actuels, mais beaucoup plus longs sous ces latitudes méridionales. Il n’est pas improbable que des migrations saisonnières, vers le nord et le sud, aient eu lieu pour les mammouths, les rhinocéros et les rennes, et que les carrières de silex du nord, le long de la Somme et de la Marne, aient été principalement fréquentées pendant la saison chaude de l’été. Les mammifères asiatiques avaient entièrement disparu des régions de France et d’Allemagne lors du premier pic de la quatrième glaciation, mais certains soutiennent que, lors de l’amélioration climatique qui a suivi, une période dans la région alpine appelée Laufenschwankung par Penck, l’éléphant à défenses droites et le rhinocéros de Merck ont de nouveau migré vers le nord de la France. Il est vrai que l’on trouve parfois des os de ces animaux en étroite association avec ceux du mammouth laineux et du rhinocéros laineux. On peut expliquer ce mélange soit par l’avancée de la quatrième glaciation, soit par la migration vers le nord et vers le sud des troupeaux respectifs de mammifères en été et en hiver. Avec la poursuite de la quatrième glaciation, il est certain que ces mammifères asiatiques ont entièrement disparu.
Au même moment, les Néandertaliens avaient franchi le premier stade de développement de l’industrie moustérienne et avaient atteint ce qu’on appelle le Moustérien « Tull » ou « haut » Moustérien, qui, à quelques exceptions près, se déroulait à l’abri des falaises surplombantes ou à l’intérieur des grottes.
La vie mammifère de ces périodes « moustériennes » « complètes », telle qu’on la trouve le long des sources du Danube, du Rhin et des bras de la Dordogne et de la Vère, est divisée entre les différents groupes fauniques comme suit :
La vie à l’époque du Moustérien moyen
- La vie dans la toundra.
- Mammouth laineux.
- Rhinocéros laineux.
- Renne scandinave.
- Renard arctique.
- Lièvre arctique.
- Lemming à bandes.
- Lagopède arctique.
- La vie alpine.
- Marmotte des Alpes.
- Bouquetin.
- Lagopède alpin.
- La vie des steppes.
- Cheval des steppes.
- Souslik des steppes.
- Poule d’eau.
- La vie asiatique.
- Lion des cavernes.
- Hyène des cavernes.
- Léopard des cavernes.
- La vie en forêt.
- Cerf, lynx, loup, renard, campagnol, ours brun, cerf géant.
- Ours des cavernes.
- La vie dans les prés.
- Bison.
- Bovins sauvages.
Français Il semblerait que le renne, le mammouth laineux et le rhinocéros laineux étaient déjà largement répandus en Europe occidentale, accompagnés du renard arctique (Cams lagopus), du lièvre arctique (Lepus variabilis) et du lemming rayé (Myodes torquatus). Il n’y a aucune preuve que le bœuf musqué ait atteint à cette époque son aire de répartition extrême méridionale, et il semblerait que l’arrivée du deuxième type de lemming nordique de la région de la rivière Obi (Myodes obensis) n’ait eu lieu qu’à la fin de l’époque moustérienne,21 car la grande migration de ces animaux est attestée par leurs abondants restes dans ce qu’on appelle la « couche inférieure des rongeurs » de toutes les stations le long du Rhin et du Danube, telles que Sirgenstein, Wildscheuer et Ofnet, après le stade final de l’industrie moustérienne. Français En fait, ce remarquable petit rongeur semble marquer le deuxième maximum ou la fin de la quatrième glaciation par sa migration dans toute l’Europe occidentale, et partout où ses restes sont trouvés dans les dépôts des grottes, ils fournissent l’une des dates préhistoriques les plus importantes et les plus positives, à savoir celle de la [ p. 207 ] « couche inférieure des rongeurs ». Les lemmings surpassent tous les autres mammifères par les grandes distances parcourues par leurs migrations, et il semblerait que cette espèce nordique ait balayé toute l’Europe occidentale à la même époque, laissant ses restes non seulement dans les cavernes le long du Danube, mais aussi dans celles de Belgique et de Thiede, près de Brunswick. Cette dernière station, Thiede, n’était pas loin de la frontière sud du glacier scandinave ; il était soumis à un climat arctique très rigoureux, car les seuls associés du lemming Obi étaient le lemming à bandes, le renard arctique, le lièvre arctique, le renne, le mammouth et le bœuf musqué.
Le mammouth laineux atteint maintenant le sommet de son évolution et de sa spécialisation ; tel qu’il est préservé dans les toundras gelées du nord de la Sibérie, et tel que représenté dans de très nombreux dessins et gravures par les artistes du Paléolithique supérieur, il est le plus complètement connu de tous les mammifères fossiles22. Ses proportions, comme le montre la figure ci-jointe, qui représente les informations recueillies auprès de toutes les sources, sont entièrement différentes de celles de l’éléphant indien ou africain. La tête est très haute et surmontée d’une grande masse de poils et de laine ; derrière celle-ci, une dépression prononcée sépare l’arrière de la tête de la grande bosse sur le dos ; la partie postérieure du dos tombe très rapidement et la queue est courte ; le pelage de longs poils atteint presque le sol, et en dessous se trouve une chaude sous-couche de laine. Il n’est pas improbable que les bosses sur la tête et le dos aient été des réservoirs de graisse. La couleur des poils était d’un brun jaunâtre, variant du brun clair au brun pur ; des poils laineux, d’un pouce à un pouce et demi de long, couvraient tout le corps ; Entre les poils plus courts se trouvait un grand nombre de poils plus longs et plus épais, qui formaient des plaques semblables à une crinière sur les joues, le menton, les épaules, les flancs et l’abdomen. Une large frange de ces longs poils s’étendait le long des côtés du corps, comme le montrent les œuvres des artistes du Paléolithique supérieur dans la grotte des Combarelles. La nourriture trouvée dans l’estomac et la bouche des mammouths sibériens congelés est particulièrement intéressante pour nous. Elle consiste principalement en une flore de prairie telle que celle qui fleurit aujourd’hui en été dans le nord de la Sibérie, notamment des graminées et des carex, du thym sauvage, des haricots de l’oxytropis sauvage, ainsi que la variété arctique de la renoncule acer (Ranunculus acer). C’était la nourriture d’été. La nourriture d’hiver comprenait sans aucun doute les feuilles et les tiges du saule, du genévrier et d’autres plantes hivernales.
La vie à l’époque du Moustérien tardif
Deuxième maximum de la quatrième glaciation
- Vie de toundra, de steppe, alpine, asiatique et de prairie, comme ci-dessus.
- Obi-lemming.
- Bœuf musqué.
- L’hermine.
- Lagopède arctique.
- Belette d’Eversmann (Belette des steppes).

Le rhinocéros laineux était le compagnon invariable du mammouth, tout comme le rhinocéros de Merck s’est toujours associé à l’éléphant aux défenses droites. Cet animal remarquable est apparenté au groupe nord-africain des rhinocéros blancs, dont il est issu à une époque très reculée. Le profil de sa tête très longue et étroite, de son énorme corne antérieure et de sa petite corne postérieure, ainsi que de son dos bossu, ressemble à celui de la forme africaine actuelle, mais sa protection contre le climat arctique lui a donné une apparence extérieure totalement différente ; les poils de la face, d’une couleur brun doré, recouverts d’une sous-couche de laine, sont conservés au Musée de Petrograd. Grâce à une découverte à Starunia, en Galicie orientale, en 1911, cet animal nous est maintenant complètement connu, à l’exception de la queue ; Français ses restes ont été trouvés ici à une profondeur de 30 pieds, et comprenaient la tête, la patte avant gauche et la peau du côté gauche du corps. Le spécimen de Starunia a une lèvre supérieure large et tronquée adaptée aux habitudes de pâturage, de petits yeux obliques, des oreilles longues, étroites et pointues, une longue corne antérieure à base ovale et une corne postérieure plus courte, un cou court, à l’arrière duquel se trouve une petite bosse charnue, tout à fait indépendante du squelette ; les pattes sont relativement courtes. Il diffère de la forme africaine actuelle par son museau un peu plus étroit, ses petites oreilles pointues et la présence d’une épaisse couche de poils. Comme le rhinocéros blanc, la forme laineuse était un habitant des plaines, se nourrissant d’herbe et de petites herbes.23 Ce rhinocéros restait plus près des frontières des grandes calottes glaciaires que le mammouth, arrêtant sa migration en Allemagne et en France ; c’est-à-dire qu’il n’a pas migré aussi loin vers le sud que le mammouth, qui a erré en Italie jusqu’à Rome.
Le renne fut le précurseur de toute la faune de la toundra arctique ; il atteignit la vallée de la Vézère au début de la véritable culture moustérienne et avait déjà pénétré beaucoup plus au sud pendant la troisième période glaciaire, migrant probablement le long des frontières des champs de glace ; en fait, on le retrouve en Europe du Nord même pendant la deuxième glaciation. C’est la véritable espèce scandinave ou de terrains nus, qui est aujourd’hui typifiée par deux formes du renne de l’Ancien Monde (R. tarandus, R. spitzbergensis) et par les formes américaines actuelles de terrains nus. Les bois sont ronds, fins et longs par rapport à la taille relativement petite de l’animal ; les dents frontales sont palmées. Il existe peu de preuves que les Néandertaliens aient fait un usage important des os de renne, mais il y a tout lieu de supposer qu’ils utilisaient les peaux, pour la préparation desquelles les grattoirs et rabots moustériens étaient particulièrement bien adaptés.
[ p. 210 ]
Français Dans la péninsule Ibérique, la faune de la toundra ne pénétrait pas aussi loin au sud que le Portugal, bien que le lemming norvégien (Myodes lemmus) ait atteint les environs de Lisbonne. Le mammouth laineux, accompagné du rhinocéros laineux, a été découvert dans deux localités de l’extrême côte nord de l’Espagne, dans la province de Santander, en bordure du golfe de Gascogne. Le renne (Rangifer tarandus) est présent dans la caverne de Seriha, au sud des Pyrénées ; dès l’époque acheuléenne, il atteignait la région d’Altamira, près de Santander. Ainsi, Harlé24 conclut qu’il est certain que la faune de la toundra s’est répandue de la France vers l’ouest en Catalogne, le long de la côte nord de l’Espagne, en bordure des Pyrénées. On pense généralement que l’ours des cavernes (Ursus spelaus) occupait de nombreuses cavernes avant leur occupation par l’homme et y a développé certaines particularités de structure. Français Ainsi, les phalanges portant les griffes sont faiblement développées, ce qui indique que les griffes ont partiellement perdu leur fonction préhensile ; les dents broyeuses antérieures sont très réduites et les cuspides des dents broyeuses postérieures sont émoussées d’une manière qui indique un régime omnivore ; pourtant les pattes avant étaient d’une taille énorme, le corps était trapu et de proportions plus lourdes que celui des plus grands ours récents (Ursus arctos) d’Europe. Il semblerait donc que les Néandertaliens aient chassé des grottes un type d’ours moins redoutable que l’espèce actuelle, mais néanmoins un adversaire sérieux pour les hommes armés des armes légères de la période moustérienne.

¶ Coutumes de la chasse et de la vie dans les cavernes
Nous ne disposons que de moyens indirects pour connaître le courage et l’activité des Néandertaliens à la chasse, grâce aux ossements d’animaux chassés, retrouvés mêlés aux silex autour de leurs anciens foyers. On y trouve, comme nous l’avons vu, dans les foyers du Moustérien ancien, des ossements de bison, de bovin sauvage et de cheval, suivis à Combe-Capelle par la première apparition d’ossements de renne. Les os de bison et de cheval sauvage sont tous deux utilisés dans les enclumes osseuses de la culture moustérienne finale de La Quina. Ce que nous pensons être la période de la grande vie mammalienne de la région du Haut-Danube se trouve dans les niveaux moustériens de la grotte de Sirgenstein, d’où il ressort que les Néandertaliens chassaient le mammouth, le rhinocéros, le cheval sauvage, le bison et le bovin, ainsi que le cerf géant et le renne. Il faut cependant garder à l’esprit que lorsque ces grottes étaient désertées pendant un certain temps, les bêtes de proie revenaient, et il arrive donc souvent que les couches suivantes offrent des preuves d’une occupation alternée par l’homme et par des bêtes de proie de taille suffisante pour attirer les plus gros gibiers, tandis que les hiboux peuvent être responsables des dépôts de lemmings, comme dans la « couche inférieure des rongeurs ».
Obermaier25 a étudié attentivement les vicissitudes de la vie dans les cavernes à l’époque moustérienne. Bien avant que ces grottes ne soient habitées par l’homme, elles servaient de repaires ou de refuges à l’ours des cavernes [ p. 212 ] et à la hyène des cavernes, ainsi qu’à de nombreux oiseaux de proie. Par exemple, la grotte d’Échenoz-la-Moline, sur les eaux supérieures de la Saône, contenait les restes de plus de huit cents squelettes d’ours des cavernes, et il ne fait aucun doute que les Néandertaliens ont dû livrer de nombreuses batailles acharnées avant que les animaux ne soient chassés et que l’homme ne s’empare de la grotte. Le feu a peut-être été le moyen employé. On s’est demandé si les grottes n’étaient pas des lieux d’habitation insalubres, mais il faut se rappeler que, sauf dans certaines cavernes dotées d’ouvertures naturelles au plafond pour l’évacuation de la fumée, il n’y avait pas de véritable vie cavernicole, mais plutôt une vie de grotte, centrée autour de l’entrée de la grotte. La plus petite grotte, observe l’auteur, était considérablement plus grande et mieux ventilée que les petites cabanes enfumées de certains paysans européens ou que les huttes de neige des Esquimaux. L’obstacle le plus sérieux était l’humidité ambiante, qui variait périodiquement dans les cavernes, de sorte qu’aux saisons sèches succédaient une humidité abondante s’infiltrant à travers le plafond calcaire et le long des parois latérales. À ces périodes, les cavernes étaient probablement inhabitables, et on a observé de nombreux cas de gonflements pathologiques et d’inflammation des vertèbres, causés par une humidité extrême, dans les os des hommes comme des animaux. Les avantages compensatoires résidaient dans l’abri offert contre la pluie et le froid, une température constante à une distance modérée de l’entrée, et aussi dans le fait que les grottes étaient très faciles à défendre, car l’entrée était généralement étroite et l’accès souvent escarpé et difficile ; un haut mur de pierre en travers de l’ouverture aurait facilité la défense, et un tison enflammé aurait empêché l’approche des ours et autres bêtes de proie. Grâce à cet abri contre les intempéries et les bêtes sauvages, les grottes et les plus grandes ouvertures des cavernes étaient certainement fréquentées par les tailleurs de silex moustériens pendant les saisons les plus rudes de l’année.
Pourtant, la plus grande partie de la vie des Néandertaliens se déroulait sans aucun doute en plein air et à la chasse. Durant toute l’époque moustérienne, le gibier le plus courant était le cheval sauvage, le bœuf sauvage et le renne. La chair et les peaux étaient utilisées, [ p. 213 ] et la moelle était extraite en fendant tous les os les plus gros. Ainsi, on trouve fréquemment dans les foyers les restes de mammouth, de rhinocéros laineux, de cerf géant, d’ours des cavernes et d’ours brun. De ces bêtes de proie, les chasseurs néandertaliens obtenaient des peaux et peut-être aussi de la graisse pour les torches utilisées pour éclairer les cavernes ; il n’existe aucune preuve de l’invention de la lampe à cette époque.
Le travail des femmes consistait sans aucun doute à préparer les repas et à transformer les peaux en couvertures et vêtements. Chaque fois que possible, cela se faisait à la lumière du jour, à l’extérieur des grottes. Mais par temps froid et pluvieux, ou par grand froid hivernal, toute la tribu se réfugiait dans la grotte, se rassemblant autour des foyers alimentés en bois ; quelques recoins servaient de réserves de combustible ou de viande séchée, conservées pour les jours où le froid extrême et la neige aveuglante empêchaient les chasseurs de s’aventurer.
Il semble que le gibier ait été démembré sur place et que les meilleures parties aient été retirées. Le crâne a été ouvert pour récupérer le cerveau ; les os longs ont été conservés pour la moelle ; ainsi, les os du flanc et de l’épaule du gibier sont fréquents dans les dépôts des grottes, tandis que les côtes et les vertèbres sont rares.
Le piège faisait peut-être partie de l’art de la chasse connu des Néandertaliens. La chasse se faisait à l’aide de lances ou de fléchettes munies de pointes de silex, ainsi qu’au moyen de « pierres à lancer », que l’on trouve en grand nombre dans les niveaux moustériens supérieurs de La Quina, dans la grotte du Loup de l’Yonne, aux Cottes et en divers endroits d’Espagne. Si l’on imagine, comme c’est fort possible, que la pierre à lancer était placée dans une fronde de cuir, dans l’extrémité fendue d’un bâton, ou encore attachée à une longue lanière de cuir, on comprend aisément qu’elle devait s’avérer une arme très efficace.
Les méthodes de chasse des Néandertaliens demeurent néanmoins un mystère. Il existait une disparité marquée entre la taille et l’efficacité de leurs armes et la force et la résistance des animaux qu’ils poursuivaient. Aucun des outils très lourds de l’époque acheuléenne n’a été conservé ; les pointes de fléchettes et de lances ne sont guère améliorées ; elles ne pouvaient certainement pas pénétrer l’épaisse peau des grands mammifères de la toundra arctique, lourdement protégés par des poils et de la laine ; la chasse même des chevaux, du bétail sauvage et des rennes se faisait apparemment sans arc ni flèche, et ce, avant l’invention de la flèche barbelée ou de la pointe de lance.
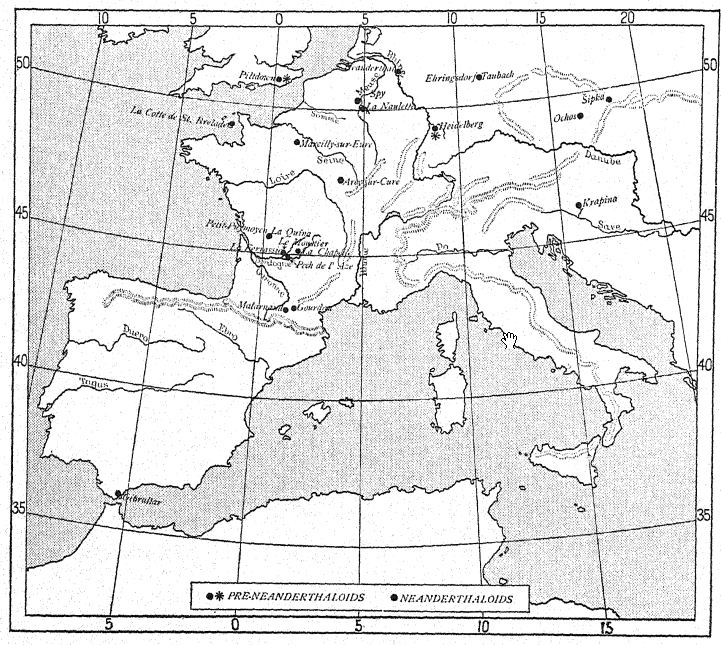
¶ Découverte des races néandertaliennes
La vie en plein air ou nomade de toutes les tribus d’Europe occidentale, depuis l’époque pré-chelléenne jusqu’à la fin de l’Acheuléen, était très défavorable à la préservation des restes humains. Il est possible que les corps des morts et des vieillards aient été jetés aux hyènes qui entouraient les stations, comme c’est le cas chez certaines tribus d’Afrique aujourd’hui, mais il est également possible qu’ils aient été enterrés de la même manière. Les squelettes enfouis près de la surface dans les sables ou graviers fluviaux des « terrasses » n’auraient pas été préservés. Nous avons vu que la préservation des restes de Heidelberg et de Piltdowm était entièrement due au hasard, les ossements ayant été lavés et mélangés à ceux des animaux ; Aucune trace d’enterrement cérémoniel ou de respect des morts n’a été trouvée dans la grotte d’Ehapina, mais au contraire, on y trouve des traces de coutumes cannibales. Même avant la fin du Moustérien ancien, tout cela avait changé. Peut-être l’association plus étroite imposée par le climat plus rigoureux a-t-elle indirectement produit un plus grand respect des morts et conduit à la coutume d’enterrer ou de disposer soigneusement les restes des morts au sol des grottes et cavernes partiellement protégées, coutume à laquelle nous devons notre connaissance actuelle de la structure de l’homme de Néandertal à l’époque moustérienne.
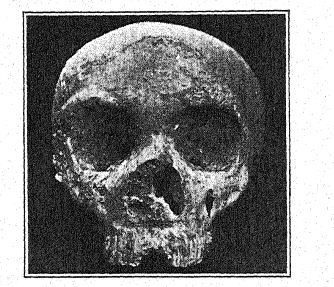
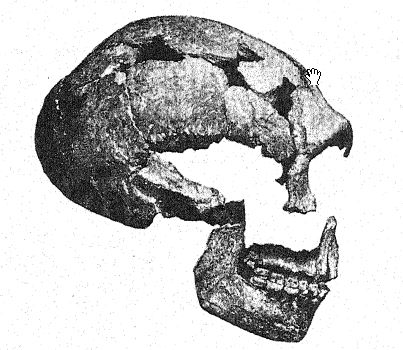
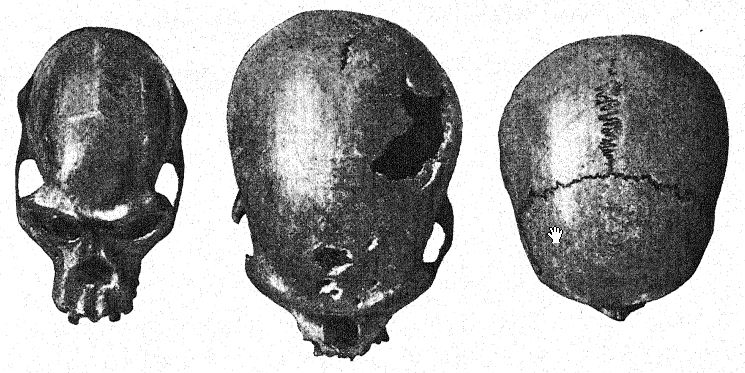
La première découverte d’un Néandertalien a été faite en 1845, huit ans avant que le type de la race néandertalienne ne soit mis au jour. Il s’agissait du crâne de Gibraltar26 trouvé par le lieutenant Flint, près de la carrière Forbes, sur la face nord du rocher de Gibraltar. Il s’agit d’un crâne bien conservé, avec seulement les os pariétaux manquants et la face et la base du crâne remarquablement complètes. En 1868, il a été offert par Busk au Musée du [ p. 216 ] Royal College of Surgeons, à Londres, où il se trouve aujourd’hui. Le lieu exact de la découverte ne peut plus être identifié avec certitude ; Français il a probablement été trouvé dans une grotte encore existante, et bien que son âge archéologique ne puisse être déterminé, comme ses caractéristiques anatoniques sont celles de la race néandertalienne, et comme tous les restes de cette race qui peuvent être datés avec certitude sont d’âge moustérien, il appartient probablement à la période moustérienne. Ces dernières années, sa grande importance dans l’histoire de l’humanité a été révélée par les études de Sollas, Keith et Schwalbe. Ainsi, il a été classé parmi les Néandertaliens et est considéré comme une forme particulièrement primitive, en raison de la taille extrêmement petite de son cerveau. Cette caractéristique et le léger développement des crêtes supraorbitaires, si caractéristiques des Néandertaliens, s’expliquent par la théorie selon laquelle le crâne appartenait à une femme.
Sera27 considère le crâne de Gibraltar comme le plus simiesque de tous les fossiles humains et pense qu’il ne devrait pas du tout être classé avec les Néandertaliens, mais devrait être considéré comme pré-Néandertalien ; ce point de vue est partagé par Keith. Boule, cependant, pense que ce crâne est du même âge géologique que ceux de Spy, La Chapelle, La Ferrassie et La Quina ; tout porte à croire,28 remarque-t-il, que le crâne de Gibraltar est un crâne féminin de type néandertalien. Il fait ailleurs référence aux crânes de Gibraltar, [ p. 217 ] de La Quina et de La Ferrassie II comme étant probablement ceux de femmes néandertaliennes.
Le crâne type de cette grande race humaine éteinte est celui de Néandertal – certainement le plus célèbre et le plus controversé de tous les vestiges anthropologiques – apprécié par Lyell et Huxley, mais ignoré par Darwin, et finalement établi par Schwalbe comme le chaînon manquant le plus important entre l’espèce humaine actuelle (Homo sapiens) et les singes anthropoïdes. En 185629, des ouvriers étaient occupés à dégager une petite grotte recouverte de terre glaise d’environ deux mètres de haut, la grotte dite de Feldhofner, dans le calcaire crétacé de la vallée connue sous le nom de Néandertal, sur le petit ruisseau Düssel coulant entre Elberfeld et Düsseldorf. Ils y découvrirent des ossements humains, probablement un squelette complet représentant une sépulture, qui, malheureusement, furent dispersés et broyés. Le docteur Fuhlrott a récupéré les parties restantes, dont la désormais célèbre calotte crânienne, les deux fémurs, l’os du bras droit, des portions de l’avant-bras, des os des deux côtés, la clavicule droite, ainsi que des fragments du bassin, de l’omoplate et des côtes. Tous les os étaient parfaitement conservés et se trouvent aujourd’hui au musée provincial de Bonn.

Français La découverte fit grand bruit, mais l’âge de ces fossiles resta d’abord douteux ; à environ 150 pas de la grotte, dans une petite caverne similaire, on trouva des os d’ours des cavernes et de rhinocéros. En 1858 parut le mémoire de Schaaffhausen30, dans lequel il donna la première description détaillée de ces restes comme appartenant à une race primitive originale différant en tout point de l’homme moderne, et il ne dérogea jamais de ce point de vue. [ p. 218 ] En 186331, Busk, Huxley et Lyell placèrent également ce squelette dans sa véritable position intermédiaire entre l’homme et les singes anthropoïdes. L’opinion déterminée de Virchow selon laquelle il ne s’agissait pas d’un type d’homme normal a exercé une telle influence que ce n’est qu’avec les travaux classiques de Schwalbe,32 entre 1899 et 1901, que ce squelette a acquis son importance primordiale pour toujours, et même cela après la découverte de deux autres races néandertaliennes.
Au début, on l’associa, à tort, à la prétendue race de Cannstatt, mais bien avant les travaux de Schwalbe, King33 la reconnut en 1864 comme une espèce distincte d’homme (Homo neanderthalensis), « l’homme de la vallée de Neander ». Peu après la découverte des méandres thaloïdes de Spy, en Belgique, Cope34 proposa en 1893 le même nom spécifique d’Homo neanderthalensis. En 1897, Wilser35 suggéra le nom d’Homo primigenius, largement adopté en Allemagne, tandis que parmi les auteurs français, la même espèce d’homme est parfois connue aujourd’hui sous le nom d’Homo mousieriensis. Cette variété de noms sert au moins à confirmer l’opinion unanime selon laquelle cet homme du Pléistocène moyen appartient à une espèce distincte.
Étant donné que cette race était très largement répandue, on peut parler de « Néandertaliens », tandis que les races ressemblant à l’espèce néandertalienne peuvent être qualifiées de « Néandertaloïdes ». La série complète de découvertes de membres de cette race est aujourd’hui très vaste.
En 1887, les géologues belges Fraipont et Lohesy36 découvrirent dans une grotte près de Spy, non loin de Dinant sur la Meuse, les restes de deux individus aujourd’hui dénommés Spy I et Spy II. Dans la même strate que les squelettes, sous une couche de calcaire tufacé, étaient enchâssés des outils en silex d’âge moustérien, ainsi que des restes de mammouth laineux, de rhinocéros laineux, d’ours des cavernes et de hyène des cavernes. Cette découverte est l’une des plus importantes de l’histoire de l’anthropologie, car elle a permis de dater avec certitude les hommes de Spy de la période industrielle moustérienne, et aussi parce que les auteurs ont immédiatement reconnu ces hommes comme appartenant à la race des [ p. 219 ] [ p. 220 ] Néandertalien et de Cannstatt, que l’on supposait alors identiques. C’est ici que furent révélées pour la première fois les proportions du crâne et du cerveau, les caractéristiques très primitives de la mâchoire inférieure et des dents, la petite taille et plusieurs caractéristiques simiesques des os des membres ; on y observa les crêtes supra-orbitaires proéminentes du type néandertalien, le front fuyant, le profil crânien inférieur à celui des races australiennes les plus basses existantes, le crâne étroit et dolichocéphale.
RÉPARTITION DES RESTES DES NÉANDERTALIENS (Comparer Fig. 104)
| I. Des temps inconnus du Paléolithique inférieur | |||
| 1848. | Gibraltar. | Carrière Forbes. | Crâne fragmentaire. |
| 1856. | Néandertal. | Düsseldorf, Allemagne. | Calotte crânienne et fragments de squelette. |
| 1859. | Arcy-sur-Cure. | Yonne, France. | Je baisse la mâchoire. |
| 1866. | La Naulette. | Belgique. | I mâchoire inférieure. |
| 1888. | Malarnaud. | Ariège, France. | I mâchoire inférieure. |
| ?Gourdan. | Hautes-Pyrénées . | I mâchoire inférieure. | |
| 1906. | Ochos. | Moravie. | Je mâchoire inférieure. |
| 2. Avec l’industrie du Moustérien supérieur | |||
| 1887. | Espion 1 , 11. | Près de Dinant, Belgique. | Deux crânes et squelettes. |
| 1907. | Petit-Puymoyen. | Char elite, France. | Fragments de mâchoires supérieure et inférieure. |
| 1909. | Pech de l’Azé. | Dordogne, France. | Crâne d’enfant. |
| 1910. | La Ferrassie II. | Dordogne, France. | Je suis squelette (femelle). |
| 1911. | La Cotte de St. Brelade. | Île de Jersey. | 13 dents humaines. |
| 1911. | La Quina II. | Charente, France. | Crâne et fragments de squelette. |
| 3. Avec l’industrie du Moustérien moyen | |||
| 1982. | Sipka. | Moravie. | Mâchoire d’un enfant. |
| 1908. | La Chapelle-aux-Saints. | Corrèze, France. | Crâne et squelette presque complets. |
| 1909. | La Ferrassie I. | Dordogne, France. | Portions d’un squelette. |
| 1910. | La Quina I. | Charente, France. | Os du pied. |
| 4. Avec l’industrie du Moustérien ancien | |||
| 1908. | Le Moustier. | Vallée de la Vézère, Dor dogne, France. | Squelette d’un jeune. |
| 1914. | Ehringsdorf. 37 | Près de Weimar. | Mâchoire inférieure. |
| 5. Avec industrie moustérienne ou acheuléenne | |||
| 1899. | Krapina. | Croatie, Autriche-Hongrie. | Portions de nombreux squelettes d’adultes et d’enfants. |
| 1892. | Taubach. | Près de Weimar. | Une dent de lait. |
On a constaté que les membres avaient conservé la disproportion anthropoïde entre le fémur et le tibia, et on a fait une découverte importante : cet homme de Néandertal, petit, massif, aux sourcils épais et au visage terne, était incapable de se tenir parfaitement droit, la structure de l’articulation du genou étant telle que les genoux étaient constamment légèrement fléchis. Autrement dit, l’homme de l’Espion n’avait pas encore pleinement acquis la position verticale des membres inférieurs.
On peut dire que cette découverte a établi les Néandertaliens dans tous leurs caractères comme une race inférieure très distincte, mais vingt-deux ans se sont écoulés avant que cela ne soit confirmé par la découverte d’un autre type encore plus ancien de Néandertaloïde à Krapina, dans le nord de la Croatie, en Autriche-Hongrie, comme décrit à la fin du chapitre II (p. 181 ci-dessus) ; type qui, avec ses variations locales [ p. 221 ] a été bientôt déterminé comme appartenant sans aucun doute au même groupe que l’homme de Néandertal et les hommes de Spy.
Bien des années auparavant, en 1866, l’anthropologue belge Dupont38 avait découvert les restes d’un autre membre de cette race dans une grotte au bord de la Lesse, près de La Naulette, non loin de Furfooz, dans le nord de la Belgique. Cette mâchoire, aujourd’hui connue sous le nom de mâchoire de La Naulette, est de type néandertalien. Elle était associée à des os de mammouth laineux, de rhinocéros, de renne et à quelques fragments d’autres ossements humains.
De nouveau, en 1882, Maska39 découvrit dans une grotte près de Sipka, en Moravie, au sud de Sternberg et à six milles à l’est de Neutitschein, des fragments de la mâchoire inférieure d’un enfant, extraordinairement forts, épais et larges, et montrant l’arrivée des dents permanentes. De la même région provient la mâchoire d’Ochos, en Moravie, découverte par Rzehak40 vers 1906. Seule la partie alvéolaire de la mâchoire fut retrouvée, mais elle servit à démontrer la très vaste répartition géographique de la race néandertalienne.
À cette époque, la Dordogne, connue depuis longtemps pour être un centre industriel moustérienne intensif, depuis la découverte du Moustier par Lartet en 1863, n’avait livré aucun squelette, ni aucune trace anatomique du type d’homme qui l’habitait à l’époque moustérienne. Mais à partir du printemps 1908, toute une série de découvertes de ce type se succédèrent, principalement des sépultures cérémonielles, à La Chapelle-aux-Saints, à la station type du Moustier même, à La Ferrassie, une autre station sur la basse Vézère, et à La Quina.
En octobre 1910, fut découvert le crâne connu sous le nom de La Ferrassie II, d’âge moustérien tardif ; il s’agit probablement de celui d’une femme, et les restes furent disposés dans ce qui était vraisemblablement une forme particulière d’enterrement cérémoniel, car les os, au lieu d’être disposés droits dans une certaine direction, étaient dans une position accroupie ou fléchie (voir Annexe, Note X).
Le squelette de Le Moustier a été découvert par Hauser dans la grotte inférieure du Moustier, dans la vallée de la Vézère, au printemps 1908, et soigneusement retiré avec l’aide du professeur Klaatsch.41 [ p. 222 ] Il appartenait à un jeune homme d’environ seize ans. L’élément le plus intéressant de la découverte résidait dans la manière dont le squelette était disposé.42 La tête reposait sur un certain nombre de fragments de silex soigneusement empilés – une sorte d’oreiller de pierre ; le mort gisait dans une posture endormie, la tête reposant sur l’avant-bras droit. Un coup de poing exceptionnellement beau se trouvait à proximité de la main, et de nombreux os carbonisés et fendus de bœuf sauvage (Bos primigenius) étaient placés autour, indiquant une offrande alimentaire. On pensait que les silex appartenaient à l’étage acheuléen, qui sous-tend la couche de véritable industrie moustérienne, connue depuis longtemps dans cette localité ; mais les archéologues français et Schmidt considèrent ces outils comme datant du Moustérien le plus ancien, époque à laquelle il est bien connu que le coup de poing acheuléen persistait. Malheureusement, le squelette n’était pas très bien conservé et, bien qu’Edaatsch ait eu tout à fait raison de le classer parmi les Néandertaliens, il ne doit pas être considéré comme une espèce distincte (Homo mousteriensis hauseri), mais plutôt comme un membre de la véritable race néandertalienne (Homo newnderthalensis). Il s’avère également être un individu plutôt trapu, robuste et de petite taille : les bras et les jambes sont relativement courts, en particulier l’avant-bras et le tibia.

[ p. 223 ]
Au même moment où le squelette du Moustier était exhumé, les abbés A. et J. Bouyssonie, ainsi que L. Bardon43, exploraient la culture moustérienne de la grotte près de La Chapelle-aux-Saints, à quelques kilomètres à l’est du Moustier, et découvrirent un squelette qui s’est révélé être de loin le plus beau de tous les fossiles néandertaliens, comprenant un crâne remarquablement bien conservé, la quasi-totalité de la colonne vertébrale, vingt côtes, des os du bras et de la majeure partie de la jambe, ainsi que plusieurs os des mains et des pieds. Il s’agissait également de la sépulture cérémonielle d’un individu âgé de cinquante à cinquante-cinq ans, disposée avec le plus grand soin d’est en ouest dans une petite dépression naturelle. On y a également trouvé des silex moustériens typiques, ainsi que plusieurs coquillages et des restes, principalement de rhinocéros laineux, de cheval, de renne et de bison. La découverte d’un crâne mature, avec les os du visage en place et dans un état de conservation relativement parfait, sans déformation de l’ensemble du crâne, a permis pour la première fois de déterminer définitivement non seulement tous les caractères et les proportions du squelette de l’homme de Néandertal, mais aussi la taille et les proportions réelles du cerveau. Ce superbe spécimen a été envoyé au Muséum de Paris, et les descriptions préliminaires de Boule44, puis sa monographie presque sans défaut45, ont suscité un intérêt mondial pour la race néandertalienne.

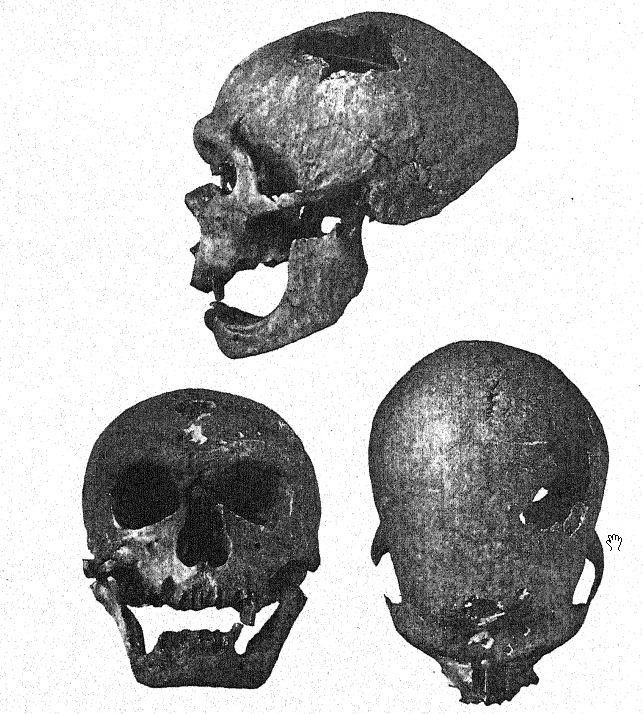
Un an plus tard, un troisième squelette néandertalien fut découvert par Peyrony dans la grotte de La Ferrassie, non loin du Bugue, en Dordogne. Les os étaient fortement brisés et les preuves d’une inhumation cérémonielle n’étaient pas parfaitement claires, mais d’un seul coup d’œil, le squelette était clairement reconnaissable grâce aux caractères du crâne, et notamment à ceux du front, comme appartenant à la race néandertalienne.
L’année suivante, en 1910, dans la grotte de La Quina, département de la Charente, au nord de la Vézère46, furent découverts les os du pied d’un homme ressemblant exactement au type de La Chapelle, et de nouveau en 1911, plusieurs parties du squelette d’un autre membre typique de l’espèce néandertalienne furent découvertes dans les premières couches moustériennes. Les os du crâne étaient quelque peu séparés au niveau des sutures. Il ne s’agissait certainement pas d’un cas d’enterrement cérémoniel. Comme le crâne de Gibraltar, il s’agirait probablement de celui d’une femme.

Français La découverte par Nicolle et Sineh47 de treize dents humaines dans une caverne moustérienne de la baie de Saint-Brélade, sur l’île de Jersey48, présente un intérêt géographique particulier, prouvant l’extension de la race néandertalienne aux îles Anglo-Normandes, alors que celles-ci faisaient probablement encore partie du continent. Ces dents étaient associées à des os de rhinocéros laineux, de renne et de deux variétés de cheval, ainsi qu’à des traces de foyers moustériens et d’outils en silex. Les caractéristiques distinctives des dents broyeuses néandertaliennes sont la taille robuste, l’implantation profonde et la forme élargie des racines, qui, avec la mâchoire lourde, témoignent de la dureté de la nourriture et de la force musculaire exercée lors de la mastication. [ p. 226 ] Les racines, au lieu de se rétrécir en pointe, comme chez l’homme moderne, forment une colonne large et robuste, soutenant la couronne, adaptée à un mouvement de balayage de la mâchoire. Cette particularité à elle seule exclurait les Néandertaliens de l’ascendance des races supérieures.
Ainsi, grâce à une longue série de découvertes, commençant en 1848 et se multipliant rapidement au cours des dernières années, nous avons trouvé les matériaux pour une connaissance complète de la structure du squelette des hommes, des femmes et des enfants de la race néandertalienne ; nous connaissons le développement relatif du cerveau ainsi que la stature des sexes ; nous avons déterminé que cette race, et elle seule, s’étendait sur toute l’Europe occidentale pendant l’Acheuléen tardif et toute la période du Mbustérien, et nous avons également appris qu’il s’agissait d’une race imprégnée de révérence pour les morts et donc probablement animée par la croyance en une forme d’existence future.
¶ Personnages de la race néandertalienne
Les crânes et squelettes49 de Néandertal, Spy, Krapina, Le Moustier, La Chapelle, La Ferrassie et Gibraltar présentent tant de traits distinctifs communs qu’il est incontestable qu’ils doivent être classés dans un groupe étroitement lié. Les traits distinctifs de ce groupe sont :
Premièrement, des caractéristiques que l’on retrouve également dans les différentes races humaines existantes, mais jamais chez les singes anthropoïdes, et donc humains ; deuxièmement, des caractéristiques qui n’ont jamais été trouvées combinées dans aucune race d’homme récent, le groupe représente donc une espèce distincte d’homme ; troisièmement, des caractéristiques en dehors des limites de variation des races humaines récentes, et intermédiaires entre elles et les limites de variation des singes anthropoïdes.
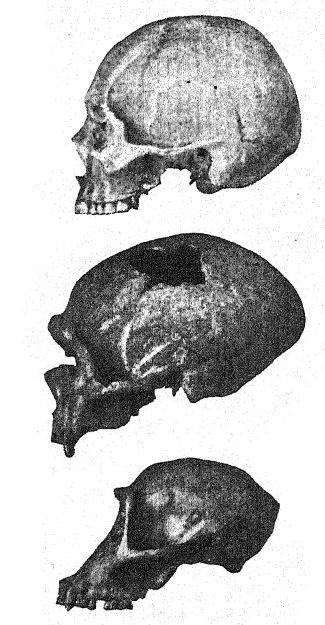
Avant d’examiner l’homme de Néandertal dans son ensemble, nous pouvons porter notre attention sur un certain nombre de ces caractéristiques particulières de la race. Tous les premiers observateurs ont été impressionnés par les sourcils épais et saillants et le front fuyant. Chez l’homme moderne, on observe souvent une proéminence marquée au-dessus des yeux, partant de la glabelle ou du point médian au-dessus du nez et s’étendant vers l’extérieur de chaque côté, mais généralement le tiers externe du bord de ces proéminences [ p. 227 ] se tourne vers le haut sous la ligne externe des sourcils. Chez les Néandertaliens, au contraire, ces proéminences sous les sourcils entourent tout le bord supérieur de l’orbite oculaire, s’étendant vers l’extérieur autour des bords externes du front, de sorte qu’on peut les appeler « tori supraorbitales » ; l’étendue de cette crête proéminente au-dessus et sur les côtés forme un véritable toit au-dessus des orbites, qui apparaissent comme deux cavernes latérales profondes. De telles proéminences latérales se produisent, bien que rarement, chez l’homme récent ; elles sont observées, par exemple, chez certains Australiens.
La vue de face du visage néandertalien, telle qu’on la voit sur le crâne féminin de Gibraltar, où les arcades sourcilières ne sont pas aussi proéminentes que sur les crânes masculins, n’en est pas moins remarquable par la grande hauteur du visage comparée à la planéité du front. En plaçant le crâne côte à côte avec celui de l’Australien, on constate immédiatement l’énorme différence de proportions du visage et du crâne chez ces deux types, bien que l’Australien représente l’une des races les plus basses d’Homo sapiens existantes ; on observe sur le crâne de Gibraltar l’espace très large entre les yeux et la très grande taille de l’ouverture nasale, qui indiquent un nez large et aplati ; il existe un espace tout aussi long entre le bas de l’ouverture nasale et la ligne d’insertion des incisives, indiquant une très longue lèvre supérieure.
[ p. 228 ]
La mâchoire est moins puissante que celle de l’homme de Heidelberg. Nous avons vu que la mâchoire de Heidelberg se distingue par sa force générale, sa lourdeur et son absence de menton, ou plutôt par un menton sans la moindre proéminence. À l’intérieur de cette plaque mentonnière très épaisse et arrondie, l’épine mentonnière caractéristique (spina mentalis) est absente ; à la place, une double rainure sert de point d’insertion aux muscles reliant le menton et la langue à l’os hyoïde. L’apophyse ascendante, point d’insertion des muscles de la mâchoire, est exceptionnellement large : 60 mm, contre environ 37 mm pour la mâchoire récente ; enfin, le condyle d’insertion au crâne est particulièrement large.
Comme la mâchoire de Heidelberg, celle des Néandertaliens se distingue par une grande épaisseur et une grande masse. En général, les contours sont similaires ; dans quelques cas, le processus mentonnier est suggéré par une légère proéminence, mais en général, le menton est fortement fuyant et il concorde avec celui de Heidelberg par l’absence de spina mentalis. Par ailleurs, il existe des différences marquées entre les mâchoires de Heidelberg et de Néandertal. La forme de cette dernière est maintenant connue grâce aux spécimens de Krapina, de Spy, de La Naulette, d’Ochos et de Sipka, ainsi qu’aux exemples parfaits de Le Moustier et de La Chapelle. Le spécimen de Sipka [ p. 229 ] prouve que même chez un enfant de dix ans, la mâchoire était remarquable par son épaisseur et sa force. Boule52 est entièrement d’accord avec Gorjanovic-Kramberger53 que le menton dans la mâchoire néandertalienne était seulement en cours de formation, et n’a atteint tout au long de la vie qu’une forme infantile, que les Néandertaliens peuvent cependant être classés comme Homines mentales, tandis que les Heidelbergiens, chez qui le menton est entièrement absent, peuvent être considérés comme Homines amentales.
Les proportions des dents des Néandertaliens sont tout aussi distinctives, notamment la taille des dents broyeuses et des dents coupantes. Comme dans la mâchoire de Heidelberg, elles forment une rangée serrée, d’où la canine ne fait pas saillie comme dans la dentition de Piltdown ; en fait, le contour de la mâchoire et les proportions des dents sont nettement humaines si on les compare à la mâchoire orangée de l’homme de Piltdown. La surface broyeuse des dents présente de nombreuses couches d’émail et les cuspides sont bien développées. Contrairement à celles de l’homme moderne, les incisives présentent des plis d’émail sur les faces internes ou linguales, une caractéristique rarement observée chez les dents coupantes modernes. Dans les dents de la mâchoire de Heidelberg, les cavités pulpaires sont exceptionnellement grandes, tandis que chez les dents de la race Krapina, on observe une caractéristique urnique : les molaires n’ont pas de racines normales, celles-ci ayant été plus ou moins résorbées, un phénomène très rare chez l’homme moderne. La dentition de La Chapelle est également distinctement humaine, mais extraordinairement massive, correspondant à la massivité générale du crâne et de l’appareil masticateur ; dans le détail, ce n’est pas celle des races civilisées, mais une forme exagérée du type appelé macrodonte.54 L’allongement de la couronne est également similaire à ce qu’on appelle l’hypsodonte.
Les dents broyeuses ne présentent pas toutes cette taille massive et cette forme colonnaire, car environ la moitié des dents de Krapina possèdent des racines distinctes et ressemblent davantage à des dents broyeuses modernes. Chez les Néandertaliens de Spy, les dents sont petites et les racines sont de taille moyenne55.
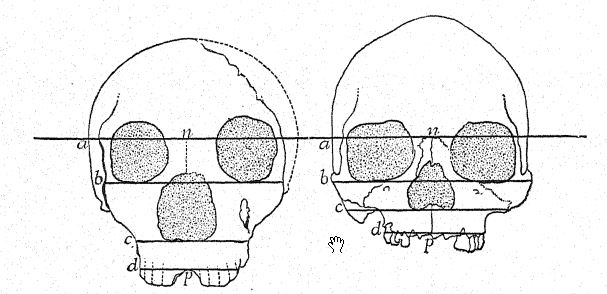
Cette étude du front et des arcades sourcilières, de la grande profondeur du visage et de la forme particulièrement haute et carrée des orbites oculaires nous prépare à une vue de profil du crâne de [ p. 230 ] La Chapelle en contraste avec celui du type européen le plus développé et le plus intellectuel, à savoir le profil du distingué paléontologue américain, feu le professeur Edward D. Cope, qui a légué son crâne et son squelette à des fins d’étude scientifique et de comparaison. Chez La Chapelle, on remarque immédiatement la platycéphalie, ou aplatissement de la calotte crânienne, le front fuyant, la proéminence marquée des arcades sourcilières, semblable à celle des singes anthropoïdes, l’allongement du visage comparé à l’aplatissement du crâne, la proéminence marquée ou prognathisme du visage dans son ensemble, la proéminence particulière des rangées de dents tranchantes comparées à la ligne verticale ou rentrante, et le recul de la rangée de dents dans le profil de Cope. Cette comparaison fait également ressortir le contraste frappant entre le menton proéminent de l’Homo sapiens et le menton profondément fuyant des Néandertaliens. Le contraste est à peine moins remarquable sur la vue supérieure du crâne, où le type néandertalien apparaît extrêmement dolichocéphale, l’arrière du crâne étant relativement large et l’avant se rétrécissant dans la région du cerveau antérieur jusqu’à s’élargir brusquement dans les processus supraorbitaires proéminents.
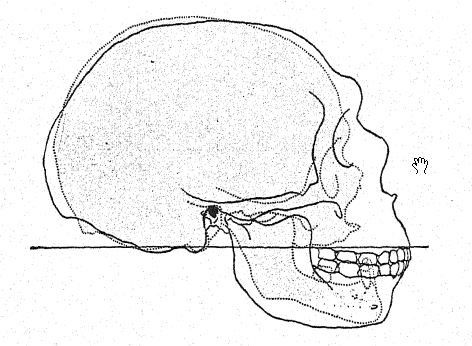
Français Comme le montre le diagramme de la page 8, Fig. i, la plus grande [ p. 231 ] longueur du crâne de Néandertal se trouve sur la ligne horizontale directement à travers la chambre cérébrale, connue sous le nom de ligne glabelle-inion, une ligne tirée d’une proéminence entre les arcades sourcilières jusqu’à un point à l’arrière du crâne connu sous le nom de protubérance occipitale externe, ou inion. C’est également la plus longue hne dans les crânes de Spy et de La Chapelle, ainsi que des singes anthropoïdes,56, mais dans le crâne d’Australie du Nord. Fig. 1, en raison de la plus grande expansion de la partie supérieure du cerveau, la plus grande longueur du crâne se situe à un point considérablement au-dessus de la ligne glabelle-inion. La section médiane du crâne du chimpanzé, de l’homme de Néandertal et de l’Australie du Nord illustre de manière frappante la généralisation faite par Schwalbe en 1901, selon laquelle le crâne de Néandertal est véritablement une forme intermédiaire entre celui des singes anthropoïdes et celui de l’Homo sapiens. Nous observons dans cette section éclairante la croissance du dôme du crâne, c’est-à-dire la grande cavité cérébrale située au-dessus de la ligne glabelle-inion g-i, en notant le contraste entre la longueur de la ligne verticale de la hauteur crânienne et l’espace situé sous la ligne glabelle-inion indiqué par les lettres. Cette ligne verticale très importante se termine en dessous, à l’ouverture où la moelle épinière pénètre dans la base du cerveau (voir Fig. 1).
[ p. 232 ]

Par de nombreux caractères, le crâne néandertalien se révèle plus proche de celui des singes anthropoïdes que de celui de l’Homo sapiens. Cette conclusion, à laquelle Schwalbe était parvenu en 1901,57, a été plus que confirmée par l’étude magistrale de Boule58 du crâne très complet de La Chapelle. Après son examen détaillé, il conclut : Quant à l’unité de la forme de la tête néandertalienne, ces traits ne sont pas particuliers au crâne de La Chapelle ; dans tous les cas, on les retrouve également dans les crânes de Néandertal, Gibraltar, Spy, Krapina, La Ferrassie, qui témoignent de l’homogénéité de ce type fossile humain appelé Néandertal. Ces traits montrent une affinité structurale entre les hommes fossiles de la période moustérienne et les singes anthropoïdes. Il convient de noter que nombre de ces traits peuvent également être trouvés dans des crânes humains récents de races inférieures, mais qu’ils sont très rares, très dispersés, très isolés, et ne se produisent que sous forme d’aberrations. C’est l’accumulation de tous ces traits dans chaque crâne d’une série entière qui constitue un assemblage entièrement nouveau et d’une grande importance. Dans le crâne, comme dans d’autres parties de l’anatomie des Néandertaliens, on ne doit pas s’attendre à trouver tous les caractères intermédiaires entre les anthropoïdes et l’homme moderne. Le long visage des Néandertaliens ressemble quelque peu à celui des Esquimaux et contraste avec le visage très court des Australiens et des Tasmaniens actuels. La dépression à la racine du nez, juste sous la glabelle, est très marquée chez tous les Néandertaliens ; l’arête nasale est moins importante que chez toutes les races récentes, à l’exception des mâles australiens, et pourtant le nez n’est pas aplati, mais plutôt arqué ou aquilin. Ce trait n’est pas caractéristique de tous les singes anthropoïdes et, à cet égard, les Néandertaliens, les Australiens et les Tasmaniens sont plus différents des singes anthropoïdes que certaines races blanches ; Français ainsi le nez néandertalien, loin de ressembler à celui des anthropoïdes, en diffère plus que celui de certains types humains récents.59 De nombreux anatomistes, à la suite de Huxley, ont décrit les crânes australiens et tasmaniens comme plus ou moins néandertaliens, et certains auteurs sont allés jusqu’à considérer ces races comme des Néandertaliens survivants. Il est vrai que certains crânes de ces races existantes sont extraordinairement [ p. 233 ] platycéphales et présentent un front fuyant, que d’autres présentent des crêtes supra-orbitaires presque aussi proéminentes que chez les Néandertaliens, que parfois la proéminence de l’inion occipital est très marquée, que certaines mâchoires présentent un menton très fuyant. Ainsi, l’une ou l’autre de ces caractéristiques néandertaliennes a été observée chez ces races inférieures existantes, mais toutes ces caractéristiques n’ont jamais été combinées dans une race comme des caractéristiques constantes, et invariablement associées, comme dans tous les crânes des Néandertaliens que nous connaissons.
En bref, le type de tête australien n’a rien de commun avec celui des Néandertaliens, si ce n’est quelques caractéristiques au niveau du front et du nez. Les traits distinctifs de la tête et du visage néandertaliens sont la platycéphalie, un front fuyant, un aplatissement de l’occiput (partie inférieure du crâne), la proéminence des crêtes supra-orbitaires, un menton fuyant ou absent, une projection de l’ensemble du visage due à la forme particulière de la mâchoire supérieure et la taille relativement petite des lobes trontaux du cerveau. En fait, conclut Boule : « Tous ces soi-disant « Néandertaliens » modernes ne sont rien d’autre que des variétés d’individus d’Homo sapiens, remarquables par l’exagération accidentelle de certains traits anatomiques qui sont normalement développés chez tous les spécimens d’Homo neanderthalensis. L’explication la plus simple de ces accidents est dans la plupart des cas l’atavisme ou la réversion. Nous ne pouvons pas affirmer qu’il n’y a jamais eu d’infusion de sang néandertalien dans les groupes appartenant à l’espèce Homo sapiens, mais ce qui semble tout à fait certain, c’est qu’une telle infusion ne peut avoir été qu’accidentelle, car il n’existe aucun type récent qui puisse être considéré même comme un descendant direct modifié des Néandertaliens. »
Cette opinion est confirmée par les recherches les plus récentes et les plus exhaustives de Berry et Robertson,60 qui concluent que ni les Australiens ni les Tasmaniens n’ont de lien direct avec l’Homo neanderthalensis ; les points superficiels de ressemblance crânienne s’expliquent uniquement par l’éloignement de l’ascendance. Les Australiens et les Tasmaniens ne descendent pas de la souche néandertalienne, mais d’une souche du Pliocène supérieur ou du Pléistocène inférieur, qui, d’après Sergi, peut être appelée Homo sapiens tasmanianus, dont l’aborigène tasmanien, aujourd’hui éteint, était la descendance presque inchangée. En ce qui concerne les caractères « inférieurs », comme le montre le diagramme (fig. 117), les crânes de Spy-Néandertal sont très proches de ceux des Tasmaniens et des Australiens, et le crâne de Gibraltar se situe à mi-chemin entre ce type et celui du Pithécanthrope en ce qui concerne douze caractères de comparaison différents.
Il est intéressant de noter[4] que les Tasmaniens se trouvaient à un stade d’industrie du silex très similaire à celui pratiqué par les Néandertaliens à l’époque moustérienne ; leurs silex étaient fabriqués à partir d’éclats produits artificiellement, y compris quelques exemples61 qui présentaient une netteté de taille des bords et une régularité de contour résultante, tandis que la plus grande partie était caractérisée par une taille maladroite et un contour irrégulier ; le statut inférieur des outils tasmaniens peut être décrit le plus correctement par le mot Pré-Aurignacien, c’est-à-dire du Moustérien ou d’un stade antérieur, mais en aucun cas « Éolithique ».
[ p. 235 ]
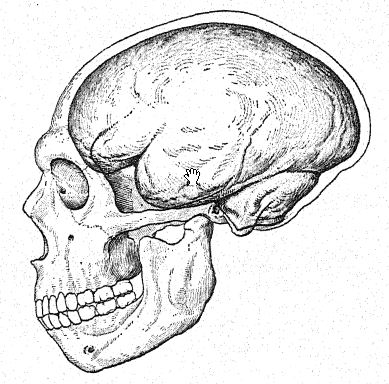
On savait que le cerveau de l’homme de Néandertal était de grande taille, même estimé à partir du type thal. Darwin fut contraint d’admettre que le célèbre crâne de Néandertal était bien développé et volumineux, et Broca proposa une explication ingénieuse du fait, autrement inexplicable, que la capacité moyenne du crâne de l’ancien habitant des cavernes est supérieure à celle de nombreux Français modernes. À savoir, la capacité moyenne du crâne des nations civilisées doit être abaissée par la préservation d’un nombre considérable d’individus, faibles de corps et d’esprit, qui auraient été rapidement éliminés à l’état sauvage, alors que chez les sauvages, la moyenne ne comprend que les individus les plus capables, capables de survivre dans des conditions de vie extrêmement difficiles. Les crânes de La Chapelle et de Spy ont permis de résoudre ce problème très intéressant, et les résultats confirment entièrement les estimations antérieures de Schaaffhausen et de Broca quant à l’importante capacité cubique du cerveau de Néandertal. Les estimations, par ordre décroissant, sont les suivantes :
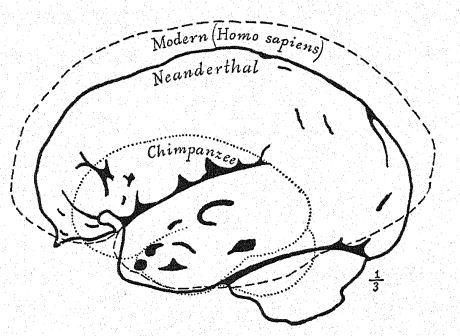
[ p. 236 ]
| Crâne de Spy II (Fraipont) | ? 1723 cm³ |
| “ La Chapelle (Boule, Verneau et Rivet) | 1626 “ |
| “ Espion I (Fraipont) | 1562 “ |
| “ Néandertal | 1408 “ |
| “ La Quina, femelle (approximation Boule) | 1367 “ |
| “ Gibraltar, femelle (estimation Boule) | 1296 “ |
Français La taille du cerveau chez les races actuelles d’Homo sapiens varie de 950 cm³ à 2020 cm³63 Ainsi, en ce qui concerne le volume de matière cérébrale, le cerveau de l’homme de Néandertal est certainement humain, mais sa forme lui manque des proportions caractéristiques de l’organisation supérieure du cerveau de l’homme moderne. Sous un autre aspect important, il est humain : la taille plus grande de l’hémisphère gauche, indiquant le développement de l’usage de la main droite. Dans sa forme générale, le cerveau ressemble davantage à celui des singes anthropoïdes par la taille relativement plus petite de la partie frontale, par la simplicité et la longueur des circonvolutions, et par la position et la direction des grandes fissures latérales connues sous le nom de « fissures de Sylvius » et de « Rolando ». Français D’après les études de Boule et Anthony64, le cerveau des Néandertaliens présente de nombreuses caractéristiques primitives. La partie antérieure du cerveau, appelée aire préfrontale, siège des facultés supérieures, n’est pas complètement développée mais présente une protubérance comme dans le cerveau des anthropoïdes. Le lobe frontal gauche en particulier, associé à la parole, est peu développé dans sa partie inférieure, ce qui laisse supposer un développement limité de la parole. La scissure latérale de Sylvius est relativement large et ouverte, ce qui, avec d’autres caractéristiques, suggère le cerveau de l’anthropoïde. Le cerveau du crâne de La Quina, que l’on pense être celui d’une femme, présente également de nombreuses caractéristiques primitives65. La capacité cubique absolue du cerveau est moins significative de l’intelligence que le développement relatif des parties du cerveau concernées par les processus mentaux supérieurs.
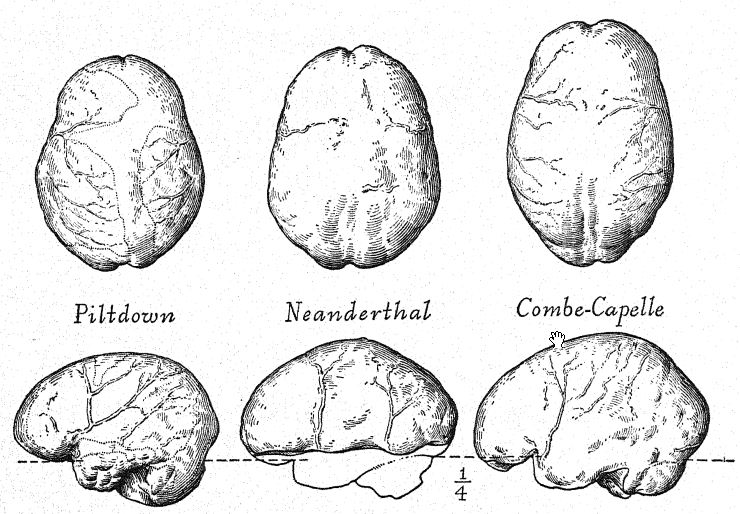
La stature des différents exemples de la race néandertalienne est estimée quelque peu différemment par Boule et par Manouvrier, et varie également selon le sexe :
| Néandertal (Boule) | 1,55 m. | 5 pi. | 1 po. |
| “ (Manouvrier) | 1,632 m. | 5 pi | 4 1/5 po. |
| La Chapelle (Boule) | 1,57 m. | 5 pieds | 1 4/5 po. |
| “ (Manouvrier) | 1,611 m. | 5 pi | 3 2/5 po. |
| Espion (Manouvrier) | 1,633 m. | 5 pi. | 4 3/10 po. |
| La Ferrassie I (Manouvrier) | 1,657 m. | 5 pieds | 5 1/5 po. |
| Moyenne des Néandertaliens supposés mâles | 1,633 m. | 5 pi. | 4 3/10 po. |
| La Ferrassie II (femelle) | 1,482 m. | 4 pieds | 10 3/10 po. |
Français La tête du Néandertalien est très grande par rapport à son corps court et trapu, qui dépasse rarement 1,65 m chez l’homme et 1,28 m chez la femme. Les proportions du corps et des membres des Néandertaliens jettent une lumière surprenante sur leur histoire ancestrale ainsi que sur leurs défauts en tant que race dépendante de la chasse. Proportionnellement à la longueur de la cuisse, la jambe est beaucoup plus courte que chez n’importe quelle race humaine actuelle. Le tibia ne représente que 76,6 % [ p. 238 ] de la longueur du fémur, alors que chez les races existantes ayant le tibia le plus court, comme les Esquimaux et la majorité des races jaunes, il n’est jamais inférieur à 80 % de la longueur du fémur. À cet égard, l’homme de Néandertal ne ressemble pas aux singes anthropoïdes, mais possède un tibia relativement plus court, car les gorilles ont un indice de 80,6 %, les chimpanzés de 82 %, les orangs-outans et les gibbons de plus de 83 % ; ainsi, tous les singes anthropoïdes et les races inférieures de l’homme ont une jambe relativement plus longue du genou vers le bas que la race de Néandertal.
La brièveté du tibia par rapport à la longueur du fémur est la preuve que les Néandertaliens étaient très maladroits et lents à marcher, car cette proportion est caractéristique de tous les animaux lents, alors qu’un tibia long et un fémur court indiquent qu’une race a naturellement le pied rapide.
De même, l’homme de Néandertal a un avant-bras très court, seulement 73,8 pour cent de la partie supérieure du bras ; il se rapproche des proportions observées chez les Esquimaux, les Lapons et les Bushmen.66 Ici encore, l’homme de Néandertal diffère des singes anthropoïdes, parmi lesquels l’avant-bras le plus court est celui du gorille, avec un rapport de 80 pour cent.
Français Il y a d’autres caractéristiques qui tendraient à montrer que les ancêtres des Néandertaliens étaient des habitants du sol plutôt que des habitants des arbres à une période géologique très reculée ; les bras sont beaucoup plus courts que les jambes, alors que chez les habitants des arbres [ p. 239 ] ils sont beaucoup plus longs. Ainsi, nous avons observé chez les singes anthropoïdes que le bras est très long par rapport à la jambe ; chez le chimpanzé, qui a relativement les bras les plus courts parmi les singes anthropoïdes, l’indice est de 104 pour cent, c’est-à-dire que les bras sont légèrement plus longs que les jambes. Au contraire, chez les Néandertaliens, la longueur des bras ne représente que 68 pour cent de la longueur des jambes ; ils sont donc très éloignés du type singe anthropoïde et se rapprochent le plus des types nègres australiens et africains.
Ainsi, pour résumer les proportions corporelles des Néandertaliens :
Bras court par rapport à la jambe, indice moyen 68 pour cent.
Avant-bras court par rapport au bras supérieur, indice moyen de 73,8 pour cent.
Tibia court par rapport au fémur, indice moyen de 76,6 pour cent.
Taille extrêmement courte par rapport à la taille de la tête.
La structure de l’épaule et du thorax est très intéressante. Toutes les côtes sont remarquablement robustes et volumineuses. Alors que chez les races actuelles elles présentent une section aplatie, chez les Néandertaliens, cette section est nettement triangulaire. Cela suggère un torse très musclé et robuste, en corrélation avec la tête gigantesque et les membres robustes. Les clavicules sont proportionnellement longues, présentant un rapport à l’humérus supérieur à 54 %, ce qui est bien supérieur à celui de la moyenne des races existantes ; cela indique une épaule très large. L’omoplate est également de type très différent de celle des races humaines supérieures, et même de celle des primates supérieurs ; elle est extrêmement courte et large.
Bien que, comme indiqué précédemment, le bras des Néandertaliens soit relativement court et donc non anthropoïde, il présente un mélange de caractères humains et simiens. Le bras supérieur, ou humérus, est véritablement de type humain, l’angle de torsion sur son axe étant de 148°, alors que chez les singes anthropoïdes, l’angle f de torsion ne dépasse jamais 141°. Parmi les os de l’avant-bras, le plus significatif est le radius, auquel est corrélé le mouvement de rotation de la main ; la structure de la tête du radius ressemble davantage à celle des singes anthropoïdes qu’à celle des espèces humaines actuelles. La structure de l’autre os de l’avant-bras, le cubitus, est également très primitive et présente certaines caractéristiques singes.
La structure de la main est un sujet du plus haut intérêt en lien avec la capacité des Néandertaliens à fabriquer des outils. La main est remarquablement grande et robuste, comparable en taille à celle des hommes de très grande taille des races actuelles. En ce qui concerne la force d’opposition du pouce contre les doigts par l’intermédiaire du muscle opposant, une caractéristique typiquement humaine, le stade de développement des Néandertaliens est nettement inférieur à celui des races actuelles, car l’articulation du métacarpien qui soutient le pouce est de forme particulière, convexe, et présente un véritable condyle convexe, alors que chez les races humaines actuelles, la surface articulaire de la partie supérieure de l’articulation du pouce est en forme de selle, c’est-à-dire concave de l’intérieur vers l’arrière et convexe de l’extérieur vers l’intérieur. Ainsi, les mouvements hautement perfectionnés du pouce chez l’Homo sapiens n’ont pas été atteints chez l’Homo neanderthalensis. Deux phalanges conservées dans le squelette de la Chapelle-aux-Saints montrent que les doigts étaient relativement courts et robustes.
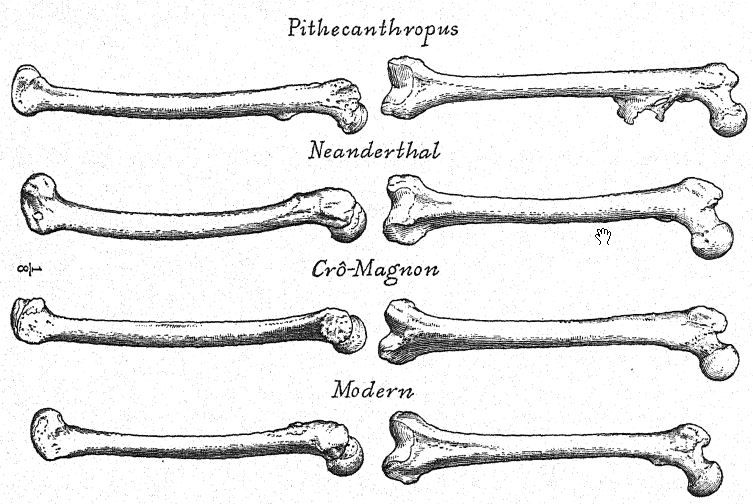
Dans la structure de la ceinture lombaire, notre homme fossile est tout à fait humain ; néanmoins, certains de ses caractères sont très primitifs et distinctifs.
De même, le fémur présente plusieurs caractères primitifs qui ne sont que rarement observés chez les races existantes, comme le troisième trochanter et la forte courbure générale vers l’avant.
La structure de l’articulation du genou par rapport au tibia est très particulière, car elle montre que le tibia était toujours rétroversé ou courbé vers l’arrière. Deux autres caractéristiques du tibia sont son extrême abréviation par rapport au fémur et l’absence d’aplatissement, ou platycnémisme. À la jonction du tibia et de la cheville (astragale), on observe deux facettes, telles qu’elles ne se conservent que chez les races d’hommes ayant conservé l’habitude de s’accroupir ou de replier leurs membres ; ces facettes ne se retrouvent pas chez les races ayant l’habitude de s’asseoir. Elles indiquent que la position de repos des Néandertaliens lors de leurs travaux industriels était accroupie, comme le montre notre reconstitution d’un Néandertalien au Moustier.
À ces membres puissants et singulièrement formés est associée une colonne vertébrale particulièrement courte et épaisse, dont chaque os est remarquable par son abréviation. Le cou, en particulier, est de construction totalement différente de celle des races humaines existantes. Il semblerait que la courbure concave du dos des Néandertaliens était dirigée directement vers le haut et se prolongeait dans la courbure concave du cou, comme chez les singes anthropoïdes, et en particulier chez le chimpanzé. Les vertèbres du cou, en particulier les cinquième, sixième et septième vertèbres, ainsi que la première dorsale, ressemblent beaucoup plus à celles du chimpanzé qu’à celles de l’Européen moderne ; les apophyses épineuses sont dirigées vers l’arrière plutôt que vers le bas. Ceci provoquait chez l’homme de Néandertal une courbure habituelle du cou et des épaules, l’empêchant de tenir la tête entièrement droite. Tandis que dans la colonne vertébrale des races existantes la position dressée est maintenue par quatre courbures gracieuses, deux vers l’avant et deux vers l’arrière, chez les Néandertaliens, comme chez les membres nouveau-nés des races supérieures, nous n’observons que trois courbures, deux concaves vers l’avant, à savoir la courbure du dos et du cou, que nous venons de décrire, et une courbure sacrée ou pelvienne ; il existe également une courbure lombaire convexe dans la partie inférieure de la colonne vertébrale des Néandertaliens, qui est cependant moins prononcée que chez les espèces humaines existantes.

En résumant les caractères de la colonne vertébrale chez les Néandertaliens, certains d’entre eux sont très primitifs, comme la structure des vertèbres du cou et le développement robuste des [ p. 243 ] apophyses épineuses, l’absence de courbure marquée dans la partie inférieure de la colonne vertébrale et la courbure très douce des os du sacrum.
Français L’aspect général de l’homme de Néandertal peut être caractérisé de la manière suivante : Une tête énorme placée sur un tronc court et épais, avec des membres très courts et trapus, et très robustes ; les épaules larges et voûtées, avec la tête et le cou habituellement penchés en avant dans la même courbure que le dos ; les bras relativement courts par rapport aux jambes ; la jambe inférieure, par rapport à la jambe supérieure, plus courte que chez n’importe laquelle des races humaines existantes ; le genou habituellement plié en avant sans la capacité de redresser l’articulation ou de se tenir complètement droit ; les mains extrêmement grandes et sans le jeu délicat [ p. 244 ] entre le pouce et les doigts caractéristique des races modernes ; le recours à une position accroupie pendant qu’il est occupé à la fabrication du silex et à d’autres industries. Ainsi, les attitudes ordinaires caractéristiques de Homo neanderthalensis seraient tout à fait différentes des nôtres et très disgracieuses. La tête lourde, le développement énorme du visage et la position en arrière du foramen magnum, par lequel la moelle épinière se connecte au cerveau, tendraient à projeter la partie supérieure du corps en avant, et cette tendance, avec la moindre courbure du cou, les épaules lourdes et la forme aplatie de la tête, donneraient à cette partie du corps un aspect plus ou moins anthropoïde.

¶ Répartition géographique des stations moustériennes
La race néandertalienne de l’époque moustérienne a établi des stations dans toute l’Europe occidentale, dont plus de cinquante ont déjà été découvertes, contre cinquante-sept stations acheuléennes ou plus connues. À certains endroits, les anciens camps ouverts des silexiers acheuléens étaient encore fréquentés, comme le long de la Tamise, de la Somme et de la Marne. Ainsi, Abbeville, Saint-Acheul, Montières et Chelles, dans le nord de la France, témoignent d’une succession d’industries moustériennes succédant à l’Acheuléen, au Chelléen et, à Saint-Acheul, même au Pré-Chelléen. Il se pourrait bien qu’il s’agisse de stations estivales, fréquentées aux saisons favorables en raison de leur abondante réserve de silex. À environ 200 kilomètres à l’est de Saint-Acheul, en Belgique, sur un petit affluent de la Meuse, se trouve la grotte de Spy, qui, avec des outils moustériens, a révélé deux squelettes fossiles humains de la race néandertalienne.
Français Dans le sud du Devonshire se trouve la célèbre caverne de Kent’s Hole, près de Torquay, découverte dès 1825 par MacEnery et décrite en 1840 par Godwin-Austen.68 Il est intéressant de noter que des dents de tigre à dents de sabre (Machcorodus latidens) ont été trouvées dans cette caverne, ce qui conduit Boyd Dawkins à penser que cet animal a survécu jusqu’à la fin des temps géologiques : il sera rappelé comme un contemporain des premiers ouvriers du silex chelléens d’Abbeville. [ p. 245 ] La vie animale de Kent’s Hole, telle que décrite à l’origine par Godwin-Austen, comprenait des restes « d’éléphant, de rhinocéros, de bœuf, de cerf, de cheval, d’ours, de hyène et d’un félin de grande taille » — une faune dont on sait maintenant qu’elle appartient à la période de la quatrième glaciation.[5]
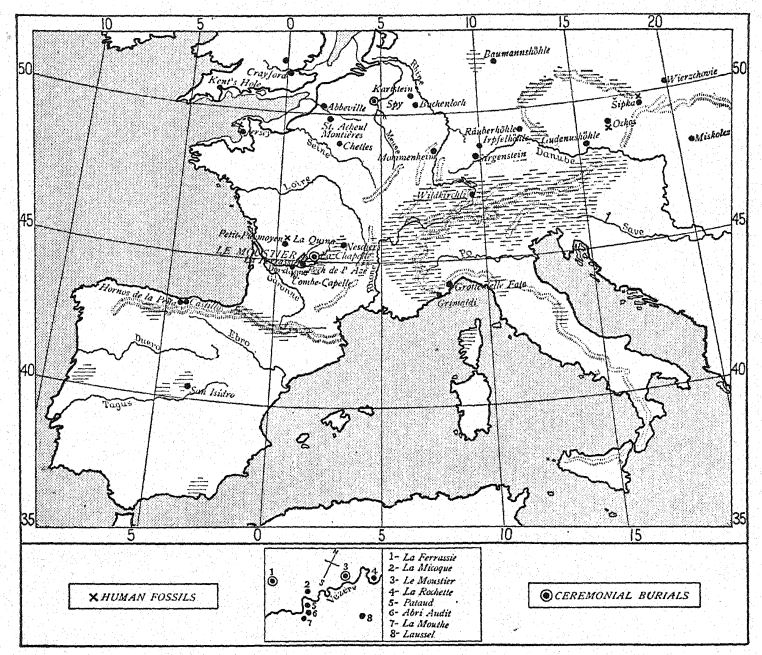
Au sud se trouvent trois stations, dont celle de La Cotte de St. Brelade, sur l’actuelle île de Jersey, alors partie continentale, qui a livré des éclats moustériens et treize dents humaines de type néandertalien.
Plus au sud encore, en Dordogne, se trouve la station type décrite précédemment, celle du Moustier, centre d’un groupe de huit sites regroupés le long des rives nord et sud de la Vézère, devenus célèbres pour les connaissances qu’ils fournissent sur les étapes successives du développement des outils moustériens, à commencer par la station de culture primitive de La Micoque, puis par La Ferrassie, Le Moustier, La Rochette, Pataud, La Mouthe, Laussel et enfin l’Abri Audit, qui marque la fin du développement de l’industrie moustérienne et, de l’avis de nombreux archéologues, de sa transition vers l’Aurignacien. Plusieurs de ces sites ont fait l’objet de découvertes importantes, tant de fossiles humains que de transitions notables dans le progrès des inventions. Autour de ce groupe vézérien se trouvent les stations de Petit-Puymoyèn, de La Quina, où ont été découverts des outils de la dernière étape de l’industrie moustérienne ainsi qu’un fossile humain de type néandertalien, et de La Chapelle-aux-Saints, qui a livré le seul squelette complet d’homme de Néandertal découvert à ce jour.

En Espagne se trouve la station de San Isidro, près des sources du Tage, et les grottes magnifiquement situées de Castillo et Hornos de la Peña, sur les pentes nord des monts Cantabriques.
[ p. 247 ]
Français Contrairement aux très nombreux sites acheuléens d’Italie, il est surprenant de constater que seulement deux grottes moustériennes ont été découvertes jusqu’à présent dans cette région : les Grottes delle Fate dans les montagnes de Ligurie, et le très important groupe d’avant-toits sur la Riviera, près de Menton, connu sous le nom de Grottes de Grimaldi, près du bord de mer et à l’endroit même où les Alpes italiennes aboutissent à la mer. En traversant vers le nord, nous remarquons la superbe grotte suisse de Wildkirchli, sur les sources du Rhin, à 1 500 mètres d’altitude.

Français Dans toute l’Allemagne, il n’existe qu’environ sept stations d’âge moustérien incontesté. Six d’entre elles sont des grottes, et la septième, Mommenheim, est un redépôt fluvial de loess le long d’un petit ruisseau, où un seul outil a été retrouvé.69 Il est intéressant d’observer qu’en Allemagne, ces sites moustériens occupent le grand coin de territoire entre les champs de glace scandinaves au nord et les Alpes au sud, et que Wildkirchli se trouvait en fait dans la zone de glaciation ; tandis que les grottes de Rauberhöhle et de Sipka n’étaient pas loin des glaciers qui recouvraient les Carpates, et Baumannshöhle n’était pas si éloignée du grand champ de glace scandinave. Dans la région des sources du Rhin et du Danube, l’industrie du [ p. 248 ] La race néandertalienne n’a jusqu’à présent été retrouvée qu’aux stations d’Irpfelhöhle, de Rauberhöhle et de Sirgenstein. Cette dernière caverne est particulièrement importante car elle comprend toute l’histoire paléolithique de cette région, présentant une série de couches culturelles successives depuis l’époque moustérienne jusqu’à l’arrivée de la race néolithique. Plus à l’est se trouvent les Gudenushöhle, près de Krems, en Basse-Autriche, et Ochos et Sipka, en Moravie, tandis qu’au-delà de la frontière russe se trouvent Wierschovie et Miskolcz. Bien au nord-ouest de Wildkirchli se trouvent les stations de Mommenheim et de Kartstein, et au nord celle de Baumannshöhle.
¶ Le savoir-faire des Néandertaliens
La vie communautaire dense du Moustérien a peut-être favorisé une évolution sociale, le développement de l’imagination et des traditions tribales, ainsi que les débuts de croyances religieuses et de cérémonies dont on trouve des traces apparentes parmi les races très différentes du Paléolithique supérieur. Cette vie n’est cependant pas marquée par le progrès industriel ou l’invention.
Les stades successifs de l’industrie moustérienne n’ont pas encore été aussi clairement définis que ceux de l’Acheuléen (Schmidt70). Dans les stations et cavernes moustériennes ouvertes de Belgique et d’Angleterre, Schmidt a observé les stades du Moustérien ancien, moyen et récent. Breuil et Obermaier considèrent La Micoque comme appartenant à la fin de l’Acheuléen, mais comme marquant la transition vers le Moustérien. Breuil considère l’industrie de la station de Combe-Capelle comme représentant la plus ancienne véritable culture moustérienne. Les recherches menées jusqu’à présent semblent justifier les subdivisions suivantes de la culture moustérienne dans le sud-ouest de la France :
6. Culture d’Abri Audit, marquant la transition de l’industrie du Moustérien supérieur à l’Aurignacien inférieur,
5. Industrie du Moustérien tardif. Outils de type La Quina avec grattoirs et enclumes en os.
4. Industrie du Moustérien moyen, avec une prédominance de belles et grandes pointes moustériennes soigneusement « retouchées » sur le bord et parfois sur une face, une « retouche » approchant parfois la technique supérieure du Solutréen.
[ p. 249 ]
3. Industrie primitive du Moustérien ancien, avec un inventaire limité d’outils.
2. Étape de Combe-Capelle, avec coups de poing en forme de cœur et « pointes » moustériennes typiques. (Arrivée des rennes.)
1. Culture de La Micoque, transition de l’Acheuléen au Moustérien. (Pas de rennes.)
L’industrie du silex, bien que très différente dans son apparence extérieure, est reconnaissable comme une évolution directe de l’Acheuléen, avec la suppression ou le déclin de certains outils et l’amélioration d’autres. Elle est le produit du même type d’esprit travaillant avec les mêmes matériaux, mais dans des conditions climatiques différentes et avec de nouvelles exigences, notamment en matière de vêtements de protection contre les intempéries. On ne peut s’empêcher de penser que l’abandon de la vie libre et ouverte des époques chelléenne et acheuléenne primitive, ainsi que la concentration des Néandertaliens sous les abris et dans les grottes, ont eu un effet raréfiant tant sur le physique que sur l’industrie elle-même. Les outils moustériens, comparés à ceux de l’Acheuléen, donnent l’impression d’être l’œuvre d’une race moins musclée et moins vigoureuse.
Outre les nombreuses et belles transitions observées71 entre les industries acheuléennes et moustériennes à Saint-Acheul, les découvertes de Laussel, sur la Vézère, près des Eyzies, apportent des preuves solides en faveur d’un lien étroit entre ces cultures. Là, large et profonde devant cet abri de Laussel, se trouve la couche moustérienne, et juste en dessous se trouve une véritable couche acheuléenne près des eaux de la vallée de la Beune. Cela prouve qu’à l’époque acheuléenne, cette vallée était aussi approfondie qu’aujourd’hui, et une union étroite de l’Acheuléen et du Moustérien est à nouveau évidente. Dans la vallée de la Somme, près de Saint-Acheul, Commont a également observé des preuves d’un lien étroit similaire entre ces cultures. Avec de tels témoignages dans le nord et le sud de la France, la race néandertalienne, connue vers la fin de l’Acheuléen et couvrant notamment toute la période moustérienne, se rapproche beaucoup plus de nous. Si nous attribuons l’industrie moustérienne à la dernière période glaciaire, nous lui donnons une durée d’environ 30 000 ans, et c’est à peu près la durée que des anatomistes réfléchis ont déjà attribuée à l’homme de Néandertal.
¶ Outils moustériens spéciaux
Deux instruments sont particulièrement représentatifs de l’industrie moustérienne, du début à la fin : la pointe et le racloir. Le premier, pointu et en forme de lance, mesure de 2,5 à 10 cm de long ; le second est un large grattoir, de 2,5 à 5 cm de large ; tous deux ont la particularité d’être constitués d’un grand éclat de silex, détaché d’un bulbe ou d’un nodule plus grand, et d’être retouchés d’un seul côté, laissant de l’autre côté la surface conchoïdale lisse de l’éclat. Cette pointe et ce grattoir sont très caractéristiques non seulement des premiers stades, mais de l’industrie moustérienne tout au long de son évolution, y compris les types de La Quina récents, et leur mode de fabrication est manifestement une modification de l’usage fait suite à la découverte des éclats de Levallois à l’Acheuléen récent.
0 5 Ip


Un point du plus grand intérêt pour le développement industriel de l’Europe occidentale à cette époque est la découverte de l’utilisation de l’éclat, que ce soit dans les lames de Levallois ou lors du déclin moustérien du coup de poing. Les éclats retouchés, de formes diverses, étaient plus faciles à fabriquer et à réparer, et servaient tout aussi bien au dépouillement et au dépeçage du gibier, qui étaient auparavant assurés par l’ancien coup de poing73.
Français En conséquence, le coup de poing, façonné à partir du noyau du nodule, commence à jouer un rôle très secondaire et n’apparaît que rarement dans les niveaux moustériens. Même à Saint-Acheul, au cœur même de son ancien règne, on commence à trouver des formes décadentes et une exécution médiocre, ce qui rend difficile de reconnaître qu’il s’agit des successeurs des coups de poing acheuléens finement retouchés. Alors que les coups de poing de la station type du Moustier continuent de conserver les anciens motifs acheuléens — l’ovale, le cœur, le pointu — ils sont tous de plus petite taille et assez grossièrement retouchés. Ainsi, après des milliers d’années de [ p. 252 ] développement et d’utilisation, le coup de poing tombe dans une période de dégénérescence et de désuétude finale. L’histoire de cet instrument, que nous avons retracée depuis ses prototypes pré-chelléens, présente une analogie des plus intéressantes avec le cours de l’évolution observé chez tant de formes animales et végétales. Il traverse de nombreux stades d’amélioration et atteint un sommet de perfection et d’adaptation ; il entre alors en compétition avec une autre forme évoluant sur un plan fondamentalement différent et supérieur, et disparaît dans la lutte pour l’existence face à la plus grande utilité du type qui le remplace.
¶ Étapes successives de l’industrie moustérienne
La succession des étapes industrielles est particulièrement bien illustrée le long de la Vézère. La plus ancienne industrie moustérienne est celle de Combe-Capelle, avec ses coups de poing en forme de cœur, grossièrement façonnés, mais dépourvus de toute surface préparée pour la prise en main.
Dans la vallée de la Somme, Commont74 a observé les trois étapes suivantes dans le progrès de l’industrie moustérienne :
3. Une culture du Moustérien récent, située sur les couches supérieures, près du sommet du même dépôt de gravier, présente des caractéristiques techniques entièrement nouvelles. L’ancienne culture du coup-de-poing n’est plus valorisée, et tous les outils découverts ici sont des éclats travaillés sur une seule face et présentant une retouche d’une finesse extraordinaire.
2. Un horizon moustérien moyen qui se situe dans les couches inférieures d’un dépôt de gravier, appartenant au loess le plus récent, et qui ne contient qu’un petit coup de poing,
1. Un Moustérien ancien, avec de nombreux coups de poing lancéolés, se trouve à la base du « nouveau lœss », ce qui montre que la tradition du coup de poing perdure et que le type de coup de poing est toujours préservé. À ceux-ci sont associés les nouveaux types d’outils, et notamment les « pointes à main », si typiques de l’industrie moustérienne.
Les niveaux les plus récents (2, 3) contiennent des éclats plus longs, qui présentent déjà une tendance vers les lames, ou « lames », du Paléolithique supérieur.
Dans les abris et cavernes de Dordogne, on observe la même séquence industrielle, bien que la succession chronologique des strates ne soit pas toujours clairement définie. À la grotte de Combe-Capelle, les coups de poing en forme de cœur conservent avec la plus grande force les anciennes traditions, mais même ici, ils sont surpassés en nombre par les « pointes » moustériennes bien façonnées, taillées d’un seul côté.
Le développement ultérieur de l’industrie moustérienne peut être observé dans la station type du Moustier, où les niveaux inférieurs présentent un Moustérien primitif composé principalement d’éclats très fins et irrégulièrement façonnés, façonnés en petits grattoirs, pointes triangulaires, perçoirs et disques. La couche sus-jacente comprend des pointes moustériennes très soigneusement travaillées, fréquemment retouchées d’un côté sur toute la surface ; c’est ici que la technique moustérienne atteint son apogée, si bien que Schmidt la qualifie de « Moustérien supérieur ». Au-dessus de cette couche se trouve un niveau de formes typiques du Moustérien tardif, très différentes des petits éclats primitifs du niveau inférieur et ressemblant aux formes caractéristiques de La Quina, le type dominant étant le racloir finement façonné de La Quina. Les quelques petits coups de poing observés dans ce niveau du Moustier constituent la seule distinction entre l’industrie de cette région et celle de La Quina, où aucun coup de poing n’a été trouvé. Au Moustier se trouvent également les enclumes en os typiques, initialement identifiées à La Quina.
L’industrie moustérienne des Néandertaliens se consacra donc principalement au développement d’outils de plus petite taille, la plupart retouchés d’un seul côté, avec une amélioration constante de la technique. Cependant, les principaux types d’outils moustériens demeurent les mêmes qu’à l’Acheuléen, comme le montre le tableau ci-joint.
L’instrument connu sous le nom de pointe, ou « pointe à main », est une forme moustérienne principale et très caractéristique, perfectionnée depuis l’étape acheuléenne. De forme pointue et taillée sur un seul côté, elle perdure jusqu’à la fin du Moustérien, et se retrouve encore dans les niveaux moustériens de Spy, en Belgique.
La pointe double, forme à deux pointes en forme de lance, atteint parfois presque la forme allongée de la pointe solutréenne [ p. 254 ] de laurier, mais jamais sa finesse, sa symétrie et la perfection de sa technique.
Il existe cinq ou six variétés bien définies de racloirs, soigneusement façonnés à partir d’éclats. La forme principale est en croissant, avec un bord incurvé vers l’extérieur. D’autres formes ressemblent à des scies, avec des bords droits ou tranchants en couteau. Une autre forme, aux bords incurvés très nets et symétriques, présente des bords finement retouchés, comme pour lisser des tiges en os ou en bois. Le perceur est également façonné à partir d’un éclat allongé, parfois terminé par une pointe très fine à l’une de ses extrémités. Il est à noter que le grattoir, ou outil à raboter, si bien développé dans les industries du Paléolithique supérieur, n’apparaît que sporadiquement au Moustérien. Par exemple, à La Quina, aux derniers stades de l’industrie moustérienne, sur 220 instruments recueillis au hasard, on comptait 166 grattoirs de six formes différentes, 45 « pointes à main » de cinq formes différentes et 5 pointes doubles, contre 5 grattoirs ou rabots. On trouve très peu de formes en forme de couteau. Il semble que le racloir et le perjoir aient été les principaux instruments employés dans la préparation des peaux pour l’habillement.
Au début du Moustérien, le coup de poing était peut-être encore utilisé par les Néandertaliens à la chasse, et la fine « pointe » en forme de lance et la plus rare « double pointe » ont peut-être été développées en réponse aux besoins des chasseurs, qui s’aventuraient désormais à la chasse au bison, à l’urus, au cheval sauvage et au renne.
Industriel.
- Coup de poing (décadent), pierre à main,
- ovoïde.
- en forme de cœur.
- pointu.
- Hachette, hachoir.
- Grattoir, outil de rabotage.
- Pergoir, foreuse, tarière.
- Couteau, couteau.
- Racloir, grattoir,
- tranchant comme un couteau,
- bord incurvé vers l’extérieur,
- à bords sciés,
- à double tranchant,
- en forme de bec,
- à multiples facettes.
- Pointe, ‘pointe à la main’.
- Percuteur ? marteau-pierre ?
Guerre et poursuite.
- Pointe, ‘pointe à la main’.
- Pointe double, fer de lance ?
- Coup de poing, main-pierre.
- Pierre de jet.
- Couteau, couteau.
[ p. 255 ]
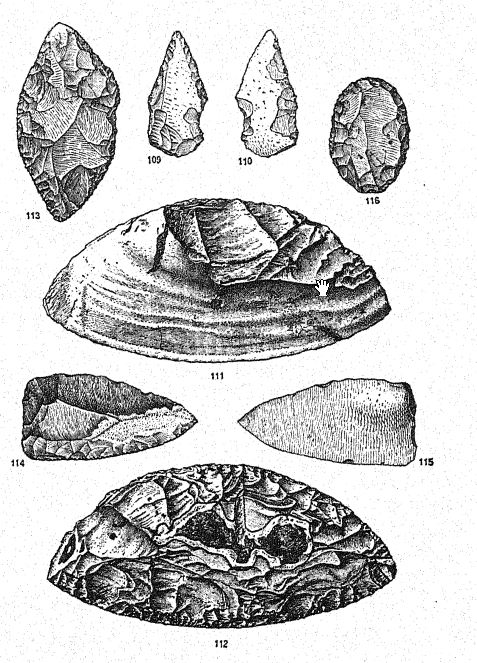
Les caractéristiques les plus frappantes de tous les instruments ayant pu servir à la chasse sont : premièrement, l’absence de toute preuve formelle de leur fixation à un manche ou à un manche ; deuxièmement, l’absence de pointe barbelée ou à tête. L’usage de la barbelure, comme nous le verrons, semble être une découverte relativement récente des cultures tardives du Paléolithique supérieur.
La transition du Moustérien à l’Aurignacien apparaît dans l’Abri Audit, également situé dans la vallée de la Vfée. On y trouve encore des coups de poing de forme irrégulière, successeurs décadents des types en forme de cœur de l’industrie moustérienne la plus ancienne ; il s’agit presque de la dernière phase du déclin de l’ancienne fabrication de coups de poing. Alors que le coup de poing en forme de lance de l’Acheuléen récent n’apparaît jamais dans aucune véritable industrie moustérienne, le type plus court et plus en forme de cœur de Combe-Capelle traverse tout le Moustérien et, après de nouvelles étapes de dégénérescence, passe dans la culture de l’Abri Audit et persiste même jusqu’à l’Aurignacien ancien. À cette dernière station, les « pointes » moustériennes typiques font presque défaut.
[ p. 256 ]
Le Moustérien, observe Schmidt,75 qui préserve les traditions de la culture du coup-de-poing du Paléolithique inférieur, est l’une des phases les plus intéressantes du développement de l’industrie paléolithique, en ce que ses étapes successives présentent les toutes dernières phases de la grande industrie du coup-de-poing, dont seuls les types d’amande et de grattoir ovale apparaissent, et cela très rarement, au début de l’Aurignacien. D’autre part, au Moustérien récent, nous observons une tendance vers l’industrie des lames du Paléolithique supérieur. L’étude et l’observation attentives des subdivisions de la culture moustérienne ont jusqu’à présent été limitées au centre et au sud de la France, et elles n’ont pas encore été retracées en Espagne ; mais dans les grottes de Belgique et d’Angleterre, les types du Moustérien ancien, moyen et récent sont connus.
Des enclumes en os, façonnées à partir des surfaces dures des os des pattes avant et des pieds du bison et du cheval, ont été découvertes à La Quina en 1906. Elles présentent une surface aplatie avec des incisions croisées trop régulières pour être accidentelles et trop éloignées de l’articulation pour être le résultat d’une tentative maladroite de sectionner l’articulation.76 Ce n’était cependant pas la seule utilisation de l’os à l’époque moustérienne, car on trouve parfois en Dordogne des outils pointus primitifs en os, mélangés à des silex moustériens. Une variété d’outils en os grossièrement façonnés est également présente à Wildkirchli, en Suisse.
¶ Disparition de la race néandertalienne
Nous avons vu que les Néandertaliens ont vécu en Europe pendant une très longue période, plusieurs milliers d’années, période durant laquelle ils ont sans doute connu une évolution considérable, passant de types inférieurs à des types supérieurs, puis à des variétés, sous l’influence du climat, de l’alimentation et des habitudes raciales. Par conséquent, les restes connus de Néandertaliens présentent une variation marquée de la forme de la tête, ainsi que de la dentition : certains sont plus primitifs et simiesques ; d’autres, comme Spy II, ressemblent davantage aux races modernes. La variété de Krapina a une tête plus large que la variété néandertalienne typique. La variété de Gibraltar est à bien des égards de type inférieur. L’individu connu sous le nom de Spy II est de type supérieur aux autres Néandertaliens. Les variations de stature connues sont minimes.
Pour ces raisons et d’autres, Hrdlicka, qui a récemment effectué une vaste étude comparative des principaux vestiges néandertaliens d’Europe, est d’avis que les Néandertaliens ont en partie évolué vers les races inférieures d’Homo sapiens ; étant non seulement dans une certaine mesure ancestraux de formes très primitives comme la race Brünn ou Predmost du Paléolithique supérieur, mais contribuant même à la race supérieure des Crô-Magnons. Il soutient également que les traces de sang et de physionomie néandertaliennes ne manquent pas, même chez les Européens modernes.
Un point de vue contraire est exposé dans le présent volume : les Néandertaliens représentent une branche secondaire de l’espèce humaine, totalement éteinte en Europe occidentale. L’auteur partage ce point de vue avec Boule et Schwalbe. Certes, les preuves apportées par les sites funéraires connus du Paléolithique supérieur ne corroborent pas la théorie de la persistance des Néandertaliens. Il est toutefois possible que les squelettes du Paléolithique supérieur découverts à Predmost, et actuellement en attente de description par Maska, puissent modifier cette conclusion et confirmer la théorie de Hrdlicka selon laquelle les Néandertaliens ont survécu et laissé des descendants ou des hommes d’origine mixte, Néandertaliens et Homo sapiens, le long de la vallée du Danube.
Quel qu’ait pu être leur sort dans d’autres régions, le changement racial le plus soudain que nous connaissions dans toute la préhistoire de l’Europe occidentale est certainement la disparition de la race néandertalienne à la fin de la période culturelle moustérienne, qui correspond à la dernière période industrielle du Paléolithique inférieur, et son remplacement par la race Crô-Magnon. D’après les données géologiques, la date de ce remplacement se situerait entre 20 000 et 25 000 ans avant notre ère. Pour autant que nous le sachions actuellement, les Néandertaliens ont été entièrement éliminés ; aucune trace de la survie du type néandertalien pur n’a été trouvée dans aucun des sites funéraires du Paléolithique supérieur ; et les cas présumés de survie de la souche néandertalienne [ p. 258 ] ou de personnes portant les caractères crâniens néandertaliens n’ont pas non plus été confirmés. Nous sommes enclins à être d’accord avec Boule et Schwalbe sur le fait que les cas supposés parmi les races modernes de transmission de caractères néandertaliens sont simplement des types inférieurs ou réversibles qui, après une analyse approfondie, ne présentent jamais la combinaison hautement distinctive et particulière des caractéristiques néandertaliennes.
Il y a des raisons de croire que les Néandertaliens ont connu une dégénérescence physique et industrielle durant les conditions de vie très difficiles de la quatrième glaciation. Cependant, l’infériorité et la diminution numérique qui en ont résulté n’expliquent pas leur extinction totale. Nous sommes enclins à l’attribuer à l’arrivée, vers la fin du Paléolithique inférieur, d’une nouvelle race hautement supérieure dans tout le pays néandertalien d’Europe occidentale. Les archéologues ont trouvé des traces d’une nouvelle culture et d’une nouvelle industrie dans certaines stations moustériennes, antérieures à la disparition de l’industrie moustérienne typique. Un tel mélange est observé dans la vallée de la Somme, dans le nord de la France.
De ces rares preuves, nous pouvons déduire que la nouvelle race a rivalisé un temps avec les Néandertaliens avant de les déposséder de leurs principales stations et de les chasser du pays ou de les tuer au combat. Les Néandertaliens, sans aucun doute, combattaient avec des armes de bois et avec la fléchette et la lance à pointe de pierre, mais rien ne prouve qu’ils possédaient l’arc et les flèches. Il existe au contraire une certaine possibilité que la race Crô-Magnon nouvellement arrivée ait pu être familière avec l’arc et les flèches, car une flèche barbelée ou une pointe de lance apparaît sur des dessins d’une période ultérieure de l’histoire de Crô-Magnon, dite Magdalénienne. Il est donc possible, bien que très loin d’être démontré, que lorsque les Crô-Magnons sont entrés en Europe occidentale, à l’aube du Paléolithique supérieur, ils étaient équipés d’armes qui, avec leur intelligence et leur physique supérieures, leur auraient donné un très grand avantage dans les luttes contre les Néandertaliens.
[ p. 259 ]
¶ Bibliographie
(1) Commont, 1912.1, p. 294.
(2) Smith, W., 1894.1, chap. XV. ,
(3) Dietrich, 1910.1, pp. 329, 330.
(4) Penck, 1909.1.
(5) Leverett, 1910.1, pp. 306-314.
(6) Geikie, 1914.1.
(7) Op. cit., p. 2 j 2,
(8) Op. cit., pp. 265-266.
(9) Keith, 1911.1, p. 23, Fig. 5.
(10) Munro, 19 12.1, pp. 46, 47.
(11) Lartet, 1861.1; 1875.1,
(12) De Vibraye, 1864.1.
(13) Massenat, 1868.1.
(14) Smith, W., 1894.1, chap. XIV. (is) Geikie, 1914.1, p. 119,
(16) Smith, Wy op. cit. pp. 196, 197.
(17) Op. cit., p. 224.
(18) Geikie, 1914. i, p. 118.
(19) Bachler, 19 1 2.1.
(20) Schmidt, 1912. 1, pp. 18-32, 165 171.
(21) Op. cit., Tableau ci-contre p. 270.
(22) Osborn, 1910.1, pp. 419, 420.
(23) Niezabitowski, 1911.1.
(24) Harlé, 1908.1, p. 302.
(25) Obermaier, 1912. i, p. 135.
(26) Keith, 1911.2.
(27) Boule, 1913. 1, pp. 220, 221.
(28) Op. cit., p. 64.
(29) Fischer, 1913.1, pp. 33^, 337
(30) Schaaffhausen, 1875.1 ; 1858.1.
(31) Lyell, 1863,1, pp. 80-92.
(32) Schwalbe, 1897.1; 190i.1; 190i.2; 1904.1.
(33) Roi, 1864.1.
(34) Cope, 1893.1.
(35) Wilser, 1898.1.
(36) Fraipont, 1887.1.
(37) Schwalbe, 1 914. 2.
(38) Dupont, 1866.1.
(39) Masque, 1886.1.
(40) Rzehak, 1906.1.
(41) Fischer, 1913.1.
(42) Kiaatsch, 1909.1.
(43) Bouyssonie, 1.909. X.
(44) Boule, 1908.1 ; 1908.2 ; 1909.1 ; 1911. 1 ; 1912.1. :
(45) Boule, 1913.1.
(46) Martin, H., 1911.1.
(47) Nicolle, 1910.1.
(48) Keith, 19 II. I.
(49) Fischer, 1913. i, P- 352.
(50) Schwalbe, 1914. i, p. 544, fig. 4 et 5.
(51) Fischer, op. cU.
(52) Boule, 1913. 1, p. 85.
(53) Gorjanovic-Kramberger, 1909.1.
(54) Boule, 1913. 1, p. 104.
(55) Tomes, 1914.1, pp. 588-598.
(56) Schwalbe, 1901.2; 1914.x, pp. 534, 535.
(57) Schwalbe, 1901.1.
(58) Boule, 1913. 1.
(59) Op. cit., p. 66, 67, 72, 75*
(60) Berry, 1914.1.
(61) Johnson, 1913.1.
(62) Quatrefages, 1884.1, p. 394.
(63) Martin, R., 1914. i, p. 645.
(64) Boule, 1910.1; 1911.1.
(65) Antoine, 1912.1.
(66) Boule, 1913. 1, p. 119.
(67) Op. cit., p. 120.
(68) Geikie, 1 914. 1, p. 130; Godwin Austen, 1840, 1. .
(69) Schmidt, 1912.1, pp. 23, 32, 66,
75, 76, loi, 169.
(fo) Op. cit., p. 12S.
(71) Schuchhardt, 19x3.1, p. 144.
(72) Dechelette, 1908.1, vol. I, pp. 98-101.
(73) Obermaier, 19 12.1, p. 130.
(74) Commune, 1909.1.
(75) Schmidt, 1912. 1, pp. 126-128.
(76) Dechelette, 1908.1, vol. I, pp. 104, 105.
(77) Hrdlicka, 1914.1
¶ Notes de bas de page
La quatrième glaciation a été entièrement qualifiée de Mecklembourgeoise par Geikie ;6 la récession pourrait correspondre à sa quatrième étape interglaciaire, le Forestien inférieur. Il s’agit du Wiinn de Penck dans la région alpine, avec un premier et un deuxième maximum séparés par la récession connue sous le nom de Laufenschwankung. En Amérique, c’est le Wisconsin précoce avec l’intervalle de récession péorien, suivi du Wisconsin tardif, qui constitue la dernière grande glaciation d’Amérique. ↩︎
Obermaier, Breuil et Schmidt attribuent La Micoque à la transition entre l’Acheuléen tardif et le Moustérien précoce. ↩︎
Le climat des toundras est extrême, la température hivernale tombant en moyenne à -27° F. en dessous de zéro, tandis qu’en été la température est d’environ 50° F. Dans les steppes subarctiques, la température moyenne de janvier dépasse à peine 30° F., tandis que celle de juillet est de 70° F. ↩︎
Le dernier représentant de cette race très primitive de la grande île de Tasmanie s’est éteint en 1877.62 ↩︎
Cette caverne, comme beaucoup de celles découvertes aux premiers temps de la recherche anthropologique, n’a pas été soigneusement explorée en référence à la stratification horizontale très importante des couches d’éclats de silex et de restes d’animaux. ↩︎